Черный дрозд Turdus merula L. у северо-восточной границы ареала: характеристика постювенальной линьки

Автор: Хохлова Татьяна Юрьевна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 8 (113), 2010 года.
Бесплатный доступ
Черный дрозд, постювенальная линька, северо-восточная граница ареала, адаптации
Короткий адрес: https://sciup.org/14749809
IDR: 14749809
Текст статьи Черный дрозд Turdus merula L. у северо-восточной границы ареала: характеристика постювенальной линьки
Черный дрозд номинативного подвида ( T. merula merula L. ) – представитель фауны европейских широколиственных лесов, более 150 лет неуклонно расширяющий границы гнездовой области в северо-восточном направлении [36], [35], [9], [15], [17], [18], [21] и др . Движение на север сопровождается значительными сдвигами в годовом цикле птиц, в том числе включением в него регулярных сезонных миграций. Это резко сокращает возможности осуществления в гнездовой области смены оперения – обязательной фазы годового цикла, которая в центральных популяциях вида заканчивается только к концу октября [28].
Вместе с тем линька – наиболее гибкий процесс из всех явлений жизненного цикла птиц: за счет его модификаций происходит «подгонка» годовых циклов птиц к условиям регионов с разными климатическими и фотопериодическими параметрами [3], [4], [36], [31]. Регуляция линьки осуществляется совместным действием двух механизмов фотопериодического контроля – эндогенного и экзогенного. Степень влияния каждого из них варьирует как в разных популяциях одного вида, так и у разных видов, каждый из которых сформировался в процессе приспособления к жизни в определенном широтном диапазоне [5], [31], [13]. Межвидовые различия в линьке проявляются в ее сроках, объеме, последовательности, длительности и особенностях, зависящих от местоположения птиц в пределах ареала [28], [27], [29], [31].
В предлагаемой статье представлена подробная характеристика постювенальной линьки черного дрозда в условиях Карелии. Аналогичное описание смены оперения белобровика – вида, высоко адаптированного к жизни в северотаежной зоне, опубликовано ранее [24]. Сравнение линьки этих близкородственных видов, принадлежащих разным фаунистическим комплексам, позволило выявить ряд различий и особенностей, которые в условиях данного региона обеспечивают преимущество аборигенному виду и препятствуют продвижению на север выходца из южных широт.
МАТЕРИАЛ И МЕТОДЫ
В работе проанализированы данные исследований и отловов птиц, проводившихся в разных точках Южной Карелии в 1972–2008 годах. Основная часть материалов собрана в 1979–2007 годах в Восточном Приладожье на Ладожском стационаре Института биологии КарНЦ РАН «Маячино», где велись многолетние экологопопуляционные исследования с применением массовых отловов и индивидуального мечения птиц [8], [6], [7]. Дополнительно использованы данные отловов птиц в заповеднике «Кивач» (Центральная Карелия, 1981–2008), на Педа-сельгском стационаре КарНЦ РАН (Прионежье, 1974–1977) и Ладожском стационаре ЛГУ (Гум-барицы, 1970–1981).
Всего для характеристики постювенальной линьки использовано 970 описаний состояния оперения при первых и повторных отловах 607 птиц неизвестного происхождения и 64 описания 44 местных сеголетков с точно известным возрастом, а также данные наблюдений за ходом линьки оперения 7 птенцов, взятых в вольере. При оценке процесса формирования ювенального наряда использованы данные ежедневных наблюдений за ростом и развитием птенцов из 5 гнезд и 1–2-кратные описания гнездового наряда птенцов с известной датой вылупления из 89 гнезд. Состояние оперения пойманных дроздов описывали по методике А. Р. Гагинской и Т. А. Рымкевич [2]. Наименования отдельных участков оперения приводятся в тексте с использованием принятых сокращений [11].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Сроки и ход постювенальной линьки птиц во многом зависят от сроков репродукции популя- ции. На территории Карелии черные дрозды регулярно встречаются в течение 8–9 месяцев – с марта по ноябрь. Откладка яиц в годы с ранней весной начинается в последней декаде апреля (22.04.2007), с поздней – только в середине мая (17.05.1978). Последние кладки появляются в конце июня – начале июля (14.07.2003). Таким образом, в зависимости от характера погоды продолжительность периода откладки яиц может варьировать от 6 до 9 недель, но даже в годы с поздним началом гнездования птицы имеют возможность осуществить 2 цикла размножения. И хотя не все пары выкармливают по два выводка, благодаря большому количеству повторных кладок, возобновленных после гибели части ранних гнезд, доля поздних выводков, вылетающих в июле – августе, превышает 35 % (рис. 1).
В ранних гнездах черного дрозда птенцы появляются в первой декаде мая при увеличивающемся световом дне, в последних – только в конце июля в период его быстрого сокращения. Разрыв в возрасте птенцов с предельными сроками вылупления составляет более 2,5 месяца . Это сказывается на всех параметрах постюве-нальной линьки, и прежде всего на возрасте, в котором начинается выпадение ювенального пера. У птенцов большинства воробьиных птиц он значительно меньше при поздних сроках рождения, чем при ранних, за счет чего происходит частичная синхронизация годовых циклов птиц одной популяции [5], [14], [12], [31].
Ювенальный наряд черных дроздов, как и других воробьиных, начинает формироваться на первой неделе жизни. Еще в гнезде оформляются большинство птерилий, перо на которых начинает расти в возрасте от 3–4 (брюшная, спинная) до 6–7 дней (средние нижние кроющие крыла). Черные дрозды обычно начинают насиживание задолго до завершения кладок, из-за чего вылупление птенцов иногда растягивается до 3–3,5 суток. Различия в развитии птенцов сохраняются до самого вылета, нередко увеличиваясь при неблагоприятных условиях из-за отставания в росте наиболее слабых.
К моменту вылета птенцов из гнезд (13–16-й день) их оперение уже достаточно развито для того, чтобы они могли взлетать с небольшим набором высоты. Однако окончательное формирование ювенального наряда заканчивается значительно позже.
Полное отрастание части контурного пера, заложенного в гнездовой период, завершается к 15–20 дням. Рулевые и маховые заканчивают рост и освобождаются от остатков роговых чехликов к 38–43 дням. Только после вылета из гнезда начинает оперяться боковой отдел брюшной птерилии, появляются большие нижние кроющие первостепенных и второстепенных маховых (БНКПМ, БНКВМ), верхний ряд нижних маргинальных кроющих (НКмарг), нижние кроющие плеча (НКпл); зарастают еще неопе-ренные участки в верхней проксимальной части пропотагиальной складки и верхней наружной части голени; отрастают дополнительные ряды перьев на брюшной, спинной, плечевой, головной (ушной отдел) и анальной птерилиях.
Из перечисленных категорий контурных перьев только БНКПМ и БНКВМ закладываются в определенный период. Их рост начинается в 17–27 дней и заканчивается в 32–43 дня одновременно или чуть позднее, чем у маховых и рулевых перьев. Но у некоторых особей отдельные перья нижних кроющих могут появляться позднее и завершают рост только к середине линьки. Сроки и последовательность зарастания остальных участков сильно варьируют, но в целом прослеживается тенденция к сокращению периода формирования их первой генерации к концу сезона. Так, у молодых птиц, появившихся в ранние или нормальные сроки, разрыв между началом роста БНКПМ, БНКВМ и других дополнительных участков оперения больше: в возрасте 40–50 дней после полного отрастания нижних кроющих крыла у них наступает пауза, когда не заметно каких-либо признаков отрастания пера других категорий. У птенцов из поздних выводков такой перерыв отсутствует.
Лишь у незначительной части птиц формирование ювенального оперения заканчивается до линьки, у многих происходит его совмещение с начальными этапами смены пера (I–II стадии), у единиц оно захватывает также III–IV стадии (рис. 2). При этом четкой связи между степенью перекрывания этих процессов и сроками линьки не выявлено.
Процесс зарастания аптерий очень индивидуален. Обычно он начинается незадолго до линьки или вскоре после ее начала. В отдельных случаях спинная и боковая аптерии могут оставаться неопушенными до середины II стадии, а брюшная – до III–IV стадии. В наиболее типичном варианте сначала появляется пух на спинном и боковом участках, позднее – на брюшном. Зарастание боковой аптерии может завершиться полностью уже к середине I стадии линьки, спинной – в начале III, брюшной – только на V. Рост пуха на всех трех аптериях заканчивается лишь к концу линьки.

Май Июнь Июль
Рис. 1. Сроки начала откладки яиц (1) и вылета выводков (2) в гнездах, находившихся под наблюдением в Приладожье в 1978–2007 годах (n = 201)
|
Част оперения |
Стадия линьки и ее продолжительность (сутки) |
||||||
|
Птерилия |
Отделы птсрклий |
I |
П |
III |
IV |
V |
|
|
10-12 |
9-18 |
12-20 |
9-17 |
11-14 |
|||
|
1 оловная Брюшная Спинная: Плечевая Бедренная Голенная .Анальная Хвостовая Крыловая |
лобно-зат ыпоч кый глазной ушной челюстной межчелюстной шейный вентральный пост вентральный пекторальный боковой межлопаточный крестцовый НКХ ВКХ СВКПМ БВКВМ СВКВМ МВКВМ кк ВК проп. М ал. Кр ал. ВКК ПКТМ СНКПМ СНКВМ НКК НК проп. НК март. НК плеча |
— |
|||||
|
— |
|||||||
|
—------ |
О— О— О—О—0—0 |
— —--- |
|||||
|
-0—0—»-0—0 |
|||||||
|
о+о+о+о+о-8-о |
|||||||
|
- |
4 - |
||||||
|
-гФ-гОт-о^-о-гО—о |
— |
— |
|||||
|
О—0—0—О— 0—0 0—о—о—о—о—о |
— |
—————— |
|||||
|
О— 0—0— Ф— о— о |
|||||||
|
- |
” - |
+ + + •*• + + + |
Z Z 1Z Z Z ■" |
<Н-о-|-о4^Н-о^-о |
+ + + + + "F |
||
|
Аптерии |
Брюшная Боковая Спинная |
4 4 * 4 . < , ♦ = < . |
......4.4 |
С 4 » » 4 4 4 . 4 |
» 4 • 4..... |
- 4 - 4 4 + - 1- ► |
|
|
2 t 4 ‘ ♦ |
|||||||
|
:::":::;:: |
......... |
......... |
|||||
|
* * * * ’ » ’ » ♦ |
|||||||
|
----" |
. »-»-« 2. ------3. ::: 4. |
||||||
Рис. 2. Схема постювенальной линьки черного дрозда:
1 - перо заменяется у отдельных особей; 2 - то же у большинства; 3 - то же у всех; 4 - отрастание ювенальных перьев
Последовательность смены пера у молодых черных дроздов в основных чертах укладывается в схему, обычную для дроздовых [14], но имеет свои особенности. Как правило, линька начинается с брюшной и спинной птери-лий, на периферии которых у многих птиц в этот период еще продолжают отрастать перья первой генерации (рис. 2). Процесс вовлечения в нее всей массы пера можно разделить на 3 стадии, опираясь на несколько основных птерилий, которые вступают в нее строго последовательно, в отличие от остальных птерилий, порядок замены пера которых характеризуется большой индивидуальной изменчивостью. Период отрастания пера разделен еще на 2 стадии. Кроме того, в случае необходимости по состоянию оперения на ключевых птерилиях для каждой стадии можно выделить начальную и конечную фазы.
-
1 -я стадия : от начала выпадения пера на спинной птерилии и в вентральном отделе брюшной птерилии до начала смены нижних и верхних кроющих хвоста (НКХ, ВКХ). На этой стадии в линьку вступают бедренная и голенная птерилии, у многих птиц – шейный и поствентральный отделы брюшной птерилии (шея, ПВБ), верхние кроющие пропотагиальной складки (ВКпроп) и нижние кроющие плеча (НКпл), у единиц – пекторальный отдел брюшной птерилии, верхние и нижние кроющие крыла (ВКК, НКК), нижние кроющие пропотаги-альной складки (НКпроп), верхний ряд нижних маргинальных кроющих (НКмарг).
Продолжительность стадии – 10–12 дней. Показателем перехода к ее завершающему этапу может служить появление на брюшной и спинной птерилиях перьев, опахала которых достигли половины длины пера (6–7-й день).
-
2-я стадия: от выпадения ювенальных НКХ и ВКХ до начала смены перьев на межчелюстном и челюстном участках головной птерилии. К концу этой стадии у всех птиц начинается смена перьев шейного отдела, ПВБ, ВКпроп, НКпл, НКК. У многих особей включаются в линьку пек-торальные, большие верхние кроющие первостепенных (БВКПМ) и второстепенных маховых (БВКВМ), кроющие крылышка (Кал), ВКК, средние нижние кроющие первостепенных и второстепенных маховых (СНКПМ, СНКВМ), НКпроп, НКмарг. У некоторых птиц процесс распространяется также на лобно-затылочный и ушной отделы головной птерилии, средние верхние кроющие первостепенных и второстепенных маховых (СВКПМ, СВКВМ), малые верхние кроющие второстепенных маховых (МВКВМ), карпальное перо (КК), нижние кроющие третьестепенных маховых (НКТМ). У некоторых птиц заканчивается отрастание НКК. У разных особей эта стадия продолжается от 9 до 18 дней. Ее «продвинутость» легко оценить по состоянию НКХ и ВКХ, опахала которых раскрываются на 4–5-й день отрастания и достигают половины длины пера после 7-го дня.
-
3-я стадия: от начала выпадения перьев межчелюстного и челюстного отделов до полного отрастания БВКВМ. На этом этапе в линьку включаются все оставшиеся птерилии. Лишь у некоторых особей смена пера на анальной, го-ленной птерилиях и в глазном отделе головной птерилии сдвигается на начало следующей стадии. У всех птиц в этот период завершается отрастание НКК, у большинства оканчивается также рост СВКПМ и СВКВМ, КК, Кал, ВКК, СНКПМ, СНКВМ, НКмарг, а у некоторых, кроме того, пекторальных, МВКВМ, ВКпроп, НКпроп, НКпл. Продолжительность стадии – 12–20 дней. Ее завершающий этап выделяется по состоянию межчелюстного и челюстного отделов, а также по превышению длины опахал всех НКХ и ВКХ половины длины пера.
-
4-я стадия укладывается в промежуток между окончанием отрастания БВКВМ и всех
нижних кроющих крыла и длится от 9 до 17 дней. К концу стадии у всех птиц завершается обновление ВКХ, НКХ и кроющих крыла, за исключением отдельных случаев, когда еще продолжается рост ряда ВКпроп, поздно вступивших в линьку. У многих особей заканчивается также смена контурного пера на отдельных участках туловища и головы, и только в вентральном отделе, межлопаточном отделе и на головной птерилии оперение у всех птиц продолжает расти до конца следующей стадии.
-
5-я стадия определяется по отсутствию признаков линьки нижних кроющих крыла, а также НКХ и ВКХ. Наряд на этой стадии уже почти полностью сформирован, лишь небольшая часть пера находится на последних стадиях отрастания. Продолжается 11–14 дней. Завершающая фаза наступает после того, как опахала наиболее поздно сменяющихся перьев голенной и головной птери-лий достигнут половины длины пера.
В случаях, когда определение «продвинуто-сти» линьки по всем ключевым птерилиям затруднено (при плохом освещении, дожде и т. п.), для ориентировочной оценки могут быть использованы только НКХ и ВКХ, которые начинают сменяться на 2-й стадии и полностью отрастают к началу 5-й стадии линьки. При обычной растянутости периода выпадения перьев (до 1 недели) рост нового пера на этих птерилиях продолжается 33–38 дней .
Шелушение кожных покровов можно наблюдать у отдельных особей еще до смены оперения. Но настоящее отслоение отмерших слоев у большинства птиц начинается лишь на 3–4-й стадиях линьки и заканчивается уже после ее завершения. Наличие признаков обновления кожных покровов у перелинявших особей свидетельствует о недавнем завершении смены пера и может условно считаться 6-й стадией постю-венальной линьки вида.
Полнота постювенальной линьки. Юношеское оперение черных дроздов, особенно самцов, хорошо контрастирует по цвету с пером следующей генерации. Это позволяет с большой точностью оценивать полноту его замены.
Постювенальная линька у данного вида, как и у большинства воробьиных птиц в Карелии, частичная. Не обновляются рулевые, первостепенные и второстепенные маховые перья. У подавляющего большинства сохраняются также старые БВКПМ, БНКПМ, перья бокового отдела брюшной птерилии, нижний ряд Нмарг, часть БВКВМ. У многих не сменяются Ккарп и Мал, у отдельных особей – СВКВМ, верхний ряд Нмарг, перья в дистальной части голени и отдельные контурные, преимущественно на участках, закладывающихся в послегнездовой период. При наибольшей полноте линьки сменяется все мелкое оперение, за исключением части БНКПМ и БНКВМ. Известен даже случай замены одного третьестепенного махового пера. При этом четко выявляется тенденция к сокращению полноты линьки к концу сезона.
Таблица 2
В озраст молодых дроздов, окольцованных в гнездах, при повторных отловах во время постювенальной линьки в Карелии
|
Cтадия линьки |
n |
Возраст (сутки) |
||
|
min |
max |
M ± m |
||
|
0 |
48 |
14 |
57 |
34,60 ± 1,44 |
|
1 |
4 |
46 |
59 |
53,50 ± 3,21 |
|
2 |
2 |
64 |
68 |
66,00 ± 2,83 |
|
3 |
5 |
76 |
91 |
84,20 ± 2,95 |
|
4 |
3 |
85 |
110 |
98,33 ± 8,90 |
|
5 |
2 |
104 |
114 |
109,00 ± 7,07 |
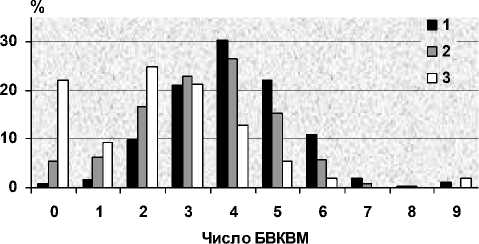
Рис. 3. Число несменяющихся ювенальных БВКВМ у молодых черных дроздов на северо-востоке (Карелия) и юго-западе ареала (Великобритания, по [25] ) (1 – Карелия, 61–62° с. ш., 2 – Великобритания, 57° с. ш., 3 – Великобритания, 51° с. ш.)
Таблица 1
Число несменяющихся ювенальных больших верхних кроющих второстепенных маховых перьев (БВКВМ) при разных сроках постювенальной линьки черных дроздов в Карелии
Стадия линьки Число несменившихся БВКВМ у молодых особей, отловленных в данный период, M + m / n (lim)
|
01–15.08 |
16–31.08 |
01–15.09 |
16–30.09 |
01.10–15.11 |
|
|
3 |
3,42 ± 0,67 |
3,85 ± 0,27 |
4,28 ± 0,21 |
5,12 ± 0,32 |
5,00 ± 0,00 |
|
6 (1–5,5) |
23 (2–6) |
38 (2–7) |
21 (2,5–9) |
1 (5) |
|
|
4 |
3,50 ± 0,43 |
3,83 ± 0,19 |
4,52 ± 0,19 |
4,63 ± 0,20 |
|
|
– |
5 (2–4) |
43 (1–6) |
45 (2–7) |
8 (4–5) |
|
|
5 |
3,26 ± 0,30 |
3,58 ± 0,18 |
4,57 ± 0,37 |
||
|
17 (0–5) |
70 (0–9) |
14 (1–7) |
|||
|
6 |
3,28 ± 0,27 |
3,44 ± 0,48 |
|||
|
9 (2–4) |
9 (1–5) |
Наиболее широко используемым показателем полноты линьки является число несменив-шихся ювенальных БВКВМ и БВКТМ. В Карелии у молодых черных дроздов (N = 258) может оставаться от 0 до 9 перьев, при этом у большинства (75 %) – 3–5 перьев, в среднем 3,9 пера. Их разное число на левом и правом крыльях отмечено у 7,4 % птиц, хотя, по-видимому, во многих случаях это вызвано внешними причинами (травмами). При поздних сроках линьки ее объем сокращается и, соответственно, число неме-няющихся БВКВМ увеличивается (табл. 1). Однако связь полноты замены БВКВМ и остального оперения у черного дрозда лишь статистическая из-за большого разнообразия индивидуаль- ных вариаций. Так, например, у птицы, сменившей не только все БВКВМ и КК, но и 19-е маховое перо, все же сохранились 2-е и 3-е Мал. Вместе с тем у нескольких особей с 5 и 6 старыми БВКВМ полностью заменялось оперение крылышка.
Полнота линьки у дроздов в Карелии достоверно меньше, чем в юго-западных частях ареала вида, причем не только в сравнении с их местными популяциями, но и с зимующим населением этих регионов [34], [33], [37], [25], [19]. Его уменьшение по направлению к северо-востоку вполне закономерно (рис. 3) и соответствует возрастанию доли птиц с поздними сроками линьки в северных популяциях. Вместе с тем сравнение этих же популяций по другим показателям, характеризующим линьку, выявило различия, которые, вероятно, обусловлены генетически. Так, у дроздов, обитающих в Англии у 57° с. ш. [25] и отличающихся значительно большим объемом линьки БВКВМ, неожиданно низкой оказалась доля особей, сменивших КК (38 % против 40 % в Карелии) и особенно Мал (1 и 12 %).
Возраст, в котором у черных дроздов карельской популяции начинается постювенальная линька, колеблется от 45 до 70 дней. Лишь в одном случае она отмечена у 42-дневного птенца из очень позднего выводка, содержавшегося в вольере. У этой птицы начало линьки совпало с завершением отрастания ювенальных первостепенных маховых перьев, у основания которых в тот момент еще сохранились остатки роговых чехликов. Как и у других птиц, смена оперения у молодых черных дроздов из поздних выводков протекает в более раннем возрасте. Однако эта известная закономерность выражена у данного вида слабее, чем у северных видов дроздов, и выявляется только при сопоставлении крайних вариантов.
В отличие от белобровиков, меняющих оперение в ранние сроки, большинство молодых черных дроздов вынуждены покидать регион до полного отрастания нового оперения: на его завершающем этапе (5-я стадия) поймано всего 2 птенца из первых выводков, вылупившихся 02 и 10.06 (табл. 2).
дата отлова
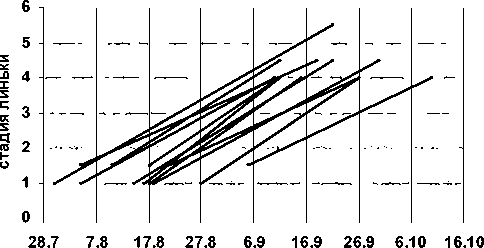
Рис. 4. Примеры хода постювенальной линьки черных дроздов при разных сроках ее начала (по данным повторных отловов маркированных особей)
Индивидуальная продолжительность линьки, по данным повторных отловов меченых особей (рис. 4) и расчетным данным, варьирует в Карелии в пределах 2 недели – от 56 до 70 дней, составляя в среднем 65 дней. Все оперение вовлекается в линьку за 35–45 дней, формирование перьев новой генерации продолжается еще 20–25 дней.
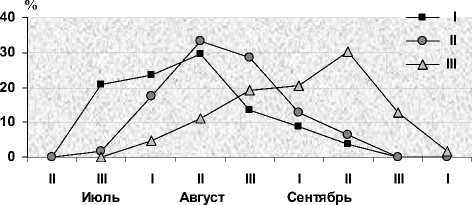
Рис. 5. Динамика отловов молодых черных дроздов с оперением на I–III стадиях его замены в Карелии
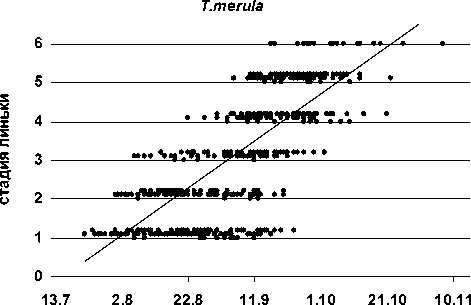
Рис. 6. Распределение в отловах молодых черных дроздов (n = 598) с оперением на разных стадиях постювенальной линьки
Разные особи проходят отдельные этапы смены ювенального наряда с разной скоростью. При этом существует тенденция к более дружному включению в нее всех участков оперения при поздних сроках линьки, хотя и среди птиц с ранней сменой оперения встречаются особи с сокращенным периодом выпадения ювенального пера. Маркерами, позволяющими оценить это явление, могут служить нижние и верхние кроющие хвоста (НКХ и ВКХ), которые начинают меняться позднее других категорий перьев. У 90 % птиц, линяющих в августе, опахала достигают половины длины пера только после полного отрастания БВКВМ (4-я стадия), у 2/3 особей с более поздними сроками линьки – задолго до его завершения (3-я стадия). Однако ускоренное выпадение перьев на мелких птерилиях мало влияет на общую продолжительность линьки, поскольку она зависит, прежде всего, от растянутости замены оперения наиболее крупных птерилий – спинной и брюшной. В связи с этим при поздних сроках линьки, как правило, сокращаются 2–3-я и удлиняются 4–5-я стадии, что сказывается на конфигурации кривых распределения в отловах птиц с оперением на разных стадиях замены, включая начальные (рис. 5).
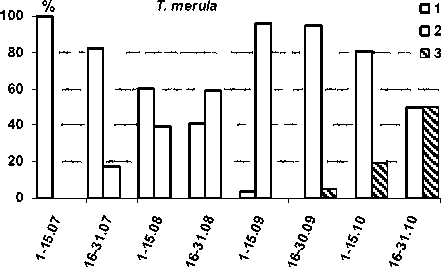
Рис.7. Состояние оперения молодых черных дроздов (n = 969) при разных сроках отловов (1 – до линьки, 2 – линяющие, 3 – перелинявшие)
Сроки линьки популяции. Молодые черные дрозды меняют оперение в очень поздние сроки (рис. 6, 7). Наиболее ранние встречи сеголетков с первыми признаками смены ювенального наряда – 22.07 (Маячино, 1989 и 1993), поздняя – 16.09 (Кивач, 1985; 23.09 эта птица все еще находилась на 1-й стадии линьки). К моменту, когда у птенцов из поздних гнезд только начинает выпадать перо, некоторые слетки уже успевают завершить линьку, однако даже они часто заканчивают ее в дискомфортных условиях из-за резкого ухудшения погоды и сужения кормовой базы к концу сентября. Поскольку подавляющее большинство черных дроздов включаются в линьку только к середине сентября (рис. 7), значительная часть особей завершает ее уже за пределами региона. Однако постепенно сроки первой регистрации птиц, завершивших смену наряда, сдвигались на более ранний период.
В первые годы полностью перелинявшие молодые птицы появлялись в отловах лишь в самом конце сентября – начале октября. Наиболее ранние даты их отловов крупногабаритными ловушками в Гумбарицах – 26.09.1973 и 28.09.1980. В Маячино, в биотопах, где молодые черные дрозды оседают на линьку, птицы, завершившие смену пера, попадались с 04.10 в 1981 и 1983 годах, а затем все раньше – 29.09.1988, 17.09.1990,
16.09.1992. Соответственно, росла доля особей, заканчивающих (5-я стадия) или закончивших линьку (imm) в более ранние сроки [30].
Видовые особенности постюве-нальной линьки черного дрозда, обусловленные его «южным» происхождением, ярко проявляются при сравнении с белобровиком, высоко адаптированным к жизни в северных широтах. Сравнение показало, что в условиях Карелии смена оперения этих двух видов проходит по сходной схеме и одинаково подчиняется известным закономерностям, обусловленным наличием фотопериодической регуляции: возраст, в котором птенцы начинают смену пера, и ее объем к концу сезона уменьшаются, а скорость несколько увеличивается. Вместе с тем сами показатели, по-видимому, закрепленные генетически, различны. Вариабельность включения в линьку разных пте-рилий и скорость смены пера у северного вида выше, а возраст, в котором сеголетки меняют оперение, и полнота его замены меньше [24]. Это позволяет северному виду обновлять наряд на месяц раньше, избегая массового совмещения линьки с миграцией [16], [20], [23].
Миграция – следующая фаза годового цикла птиц, которая требует больших энергетических затрат и сопровождается перестройкой всего обмена веществ [5]. Поскольку в условиях Севера в более выгодном положении оказываются особи, рано заканчивающие смену наряда, можно было ожидать, что процесс закрепления черных дроздов в северных широтах будет сопровождаться сдвигом линьки на более ранний и благоприятный период. И такая тенденция была обнаружена при анализе распределения в отловах птиц с разным состоянием оперения в первые годы формирования популяции и в последующий период [30]. Так, если во второй декаде сентября в 1979–1990 годах перелинявшие особи составляли 33 % от числа пойманных птиц, то в 1991–2005 годах уже 42,1 % (n = 45 и 46), а в третьей декаде месяца – соответственно 50 и 66,7 % (n = 34 и 42). Применение критерия Фишера для оценки этих различий показало их высокую достоверность (p < 0,01). При этом анализ весенних температур и хода размножения птиц в Маячино [10], [1], [26] показал, что этот сдвиг нельзя связать с влиянием погодных факторов на сроки размножения. Первый период (1979–1990) в целом характеризовался более ранними, теплыми и ровными веснами, а также немного более ранними сроками гнездования птиц. Ранние даты начала кладок черных дроздов, зарегистрированные до 1990 года (n = 102), – 26.04.1990 и 27.04.1989. После 1990 года они сдвинулись на 29–30.04.2001 (n = 84); в более поздние календарные сроки (в среднем на 1 неделю) отмечено и массовое появление выводков.
Таким образом, анализ постювенальной линьки черных дроздов периферийной северовосточной популяции выявил ряд особенностей, связанных с их «южным» происхождением, которые могут тормозить экспансию этого вида на север. Вместе с тем прослеживается тенденция к постепенному смещению линьки на более ранние сроки, что можно расценивать как показатель постепенной адаптации черного дрозда к обитанию в условиях сокращенного благоприятного периода.
БЛАГОДАРНОСТИ
Выражаю искреннюю признательность В. Б. Зимину, Н. В. Лапшину, А. В. Артемьеву, М. В. Яковлевой, Л. С. Захаровой и сотрудникам Ладожского орнитологического стационара ЛГУ, которые на протяжении многих лет принимали участие в сборе материала, а также А. В. Коро-сову за помощь в статистической обработке материала.
Список литературы Черный дрозд Turdus merula L. у северо-восточной границы ареала: характеристика постювенальной линьки
- Артемьев А. В. Влияние погоды на биологию гнездования мухоловки-пеструшки, Fice-dula hypoleuca (Passeriformes, Muscicapidae) в Карелии//Зоологический журнал. 2002. Т. 81. № 7. С. 841-849.
- Гагинская А. Р., Рымкевич Т. А. Методические указания к прижизненной обработке птиц//Исследование продуктивности вида в пределах ареала. Вильнюс: Мокслас, 1973. С. 155-166.
- Дольник В. Р. Биоэнергетика линьки воробьиных птиц как адаптация к миграции//Новости орнитологии. Алма-Ата: Наука, 1965. С. 124-126.
- Дольник В. Р. Годовые циклы биоэнергетических приспособлений к условиям существования у 16 видов Passeriformes//Миграции птиц Прибалтики. Л.: Наука, 1967. С. 115-163.
- Дольник В. Р. Миграционное состояние птиц. М.: Наука, 1975. 398 с.
- Зимин В. Б., Лапшин Н. В., Артемьев А. В., Хохлова Т. Ю. Результаты кольцевания птиц в Карелии//Кольцевание и мечение птиц в России и сопредельных государствах в 1988-1999 гг. М.: Центр кольцевания птиц, 2002. С. 73-116.
- Зимин В. Б., Лапшин Н. В., Артемьев А. В., Хохлова Т. Ю. Обзор орнитологических исследований на территории Карелии//Наземные и водные экосистемы Cеверной Европы: управление и охрана: Материалы междунар. конф., посвящ. 50-летию КарНЦ РАН. Петрозаводск, 2003. С. 55-60.
- Зимин В. Б., Сазонов С. В., Лапшин Н. В., Хохлова Т. Ю., Артемьев А. В., Анненков В. Г., Яковлева М. В. Орнитофауна Карелии. Петрозаводск: Карельский научный центр РАН, 1993. С. 1-220.
- Мальчевский А. С., Пукинский Ю. Б. Птицы Ленинградской области и сопредельных территорий. Т. 2. Л.: Изд-во ЛГУ, 1983. 504 с.
- Назарова Л. Е., Филатов Н. Н. Изменчивость климата по данным метеорологических наблюдений//Климат Карелии: изменчивость и влияние на водные объекты и водосборы. Петрозаводск: Карельский научный центр РАН, 2004. С. 12-34.
- Носков Г. А., Рымкевич Т. А. Методика изучения внутривидовой изменчивости у птиц//Методика исследования продуктивности и структуры видов птиц в пределах их ареалов. Вильнюс: Мокслас, 1977. С. 37-48.
- Рыжановский В. Н. Экология послегнездового периода жизни воробьиных птиц Субарктики. Екатеринбург: Изд-во Урал. ун-та, 1997. 288 с.
- Рыжановский В. Н. Роль фотопериодических условий в жизни воробьиных птиц высоких широт//Зоологический журнал. 2008. Т. 87. № 6. С. 732-747.
- Рымкевич Т. А., Савинич И. Б., Носков Г. А. и др. Линька воробьиных птиц Северо-Запада СССР. Л.: Изд-во ЛГУ, 1990. С. 1-304.
- Хохлова Т. Ю. Эколого-фаунистическая характеристика орнитофауны Заонежья//Вестник ЛГУ. Сер. биологии. 1977. № 15. С. 22-30.
- Хохлова Т. Ю. Соотношение сроков вылупления и постювенальной линьки у трех видов дроздов в Южной Карелии//Тез. докл. ХI Прибалт. орнитол. конф. Таллинн, 1983. С. 213-215.
- Хохлова Т. Ю. О формировании периферийной популяции черного дрозда в Южной Карелии//Изучение птиц СССР, их охрана и рациональное использование: Тез. докл. IХ Всесоюзн. орнитол. конф. Л., 1986. С. 309-310.
- Хохлова Т. Ю. Особенности биологии черного дрозда Turdus merula L. у северо-восточной границы ареала//Фауна и экология наземных позвоночных. Петрозаводск: КНЦ АН СССР, 1988. С. 51-71.
- Хохлова Т. Ю. Опыт оценки масштаба иммиграции в периферийных популяциях птиц с использованием явления географической изменчивости полноты их линьки//Территориальное поведение птиц. Петрозаводск: КНЦ РАН, 1994. С. 134-151.
- Хохлова Т. Ю. Адаптивные особенности репродукции и линьки периферийной популяции черного дрозда T. merula L.//Экосистемы Севера: структура, адаптации, устойчивость: Материалы общеросс. совещания (Петрозаводск, 1993). М., 1995. С. 204-211.
- Хохлова Т. Ю. Особенности динамики численности черного дрозда (T. merula L.) в период экспансии и закрепления вида на территории Карелии//Динамика численности птиц в наземных ландшафтах: Материалы Всеросс. совещания. М., 2007. С. 101-110.
- Хохлова Т. Ю. Черный дрозд (Turdus merula) у северо-восточной границы ареала: особенности территориальных связей и миграций в период формирования периферийной популяции в Южной Карелии (обзор)//Зоологический журнал. 2010. Т. 89. № 2. С. 212-221.
- Хохлова Т. Ю., Захарова Л. С., Зимин В. Б. Сроки и динамика сезонных явлений годового цикла у дрозда-белобровика в Карелии//Фауна и экология птиц и млекопитающих северо-запада СССР. Петрозаводск: КНЦ АН СССР, 1983. С. 11-29.
- Хохлова Т. Ю., Яковлева М. В. Постювенальная линька дрозда-белобровика в Карелии//Экология наземных позвоночных. Петрозаводск, 1991. С. 40-57.
- Both C., Artemyev A. V., Blaauw B., Cowie R. J. et al. Large-scale geographical variation confirms that climate change causes birds to lay earlier//Proc. Royal Society of London. 2004. Ser. B. Vol. 271. P. 1657-1662.
- Busse P. Key to sexing and ageing of European Passerines//Beitr. Natur. Niedersachsens. 1984. Vol. 37 (Suppl.). P. 1-224.
- Ginn H. B., Melville D. S. Moult in birds. British Trust of Ornitology Guide. 19. UK: Tring, 1983. 342 p.
- Jenni L., Winkler R. Moult and ageing of European passerines. London: Academic Press, 1994. 225 p.
- Khokhlova T. Y. Juvenile moult and spatial behaviour of first-year Blackbirds Turdus merula on the northeast edge of the range//Avian Ecol. Behav. 2009. Vol. 15. P. 1-22.
- Noskov G. A., Rymkevich T. A., Iovchenko N. P. Intraspecific variation of moult: adaptive significance and ways of realization//Proc. 22 Int. Ornithol. Congr. (August 1998, Durban, South Africa). Durban: University of Natal., 1999. P. 544-563.
- Parslou J. L. Changes in status among breeding birds in Britain and Ireland. P. 5//Brit. Birds. 1967. Vol. 60. P. 390-404.
- Richter A. Zum Umfang der Jugendmauser am Flugel der Amsel Turdus merula//Ornit. Beo-bach. 1972. Vol. 69. P. 1-16.
- Snow D. W. A Study of Blackbirds. Second ed. London: Brit. Museum (Nat. History), 1988. P. 1-196.
- Spencer R. Changes in the Distribution of Recoveries of Ringed Blackbirds//Bird Study. 1975. Vol. 22. P. 177-190.
- Stresemann E., Stresemann V. Inheritance of the adaptation in moult//Proc. 14th Int. Ornithol. Congr. Oxford, 1967. P. 75-80.
- Swensson L. Identification Guide to Europaean Passerines. Second edition. Stockholm, 1975. 184 p.
- Baillie S. R., Swann R. L. The Extent of Post-Juvenile Moult in the Blackbird//Ringing & Migration. 1980. Vol. 3. P. 21-26.
