Сравнительный анализ информативности эритроцитарных антигенов и ДНК-микросателлитов как генетических маркеров в селекционно-племенной работе со свиньями канадской селекции

Автор: Проскурина Н.В., Тихомирова Т.Н., Гладырь Е.А., Ларионова П.В., Зиновьева Н.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: ДНК-технологии в генетике и селекции
Статья в выпуске: 6 т.42, 2007 года.
Бесплатный доступ
Сравнивали информативность двух систем анализа генетического полиморфизма - эритроцитарных антигенов (ЭА) и ДНК-микросателлитов (ДНК-МС) в характеристике генеалогической структуры стад свиней пород йоркшир, ландрас и дюрок канадской селекции. Показаны различия по степени гетерозиготности исследуемых групп в зависимости от используемой системы анализа. Выявлены достоверные корреляционные взаимосвязи между степенью гетерогенности родителей, рассчитанной с использованием ЭА и ДНК-МС, и признаками продуктивности, а также зависимость характера корреляций от породной принадлежности животных.
Генетические маркеры, днк-диагностика, гетерогенность подбора
Короткий адрес: https://sciup.org/142133228
IDR: 142133228 | УДК: 636.4:636.082.2:577.2
Comparative analysis of informativeness of erythrocytes antigenes and DNA-microsatellites as genetic markers in breeding-pedigree work with pigs of Canadian selection
The authors have compared the informativeness of two systems of analysis of genetic polymorphism - erythrocytes antigens (EA) and DNA-microsatellites (DNA-MS) for characteristic of genealogical structure pig's herds of the Yorkshire, Landrace and Durock of Canadian selection. The differences in degree of heterozygosis in studied groups in connection with used analysis system were shown. The reliable correlations were revealed between degree of heterozygosis of parents calculated with use of EA and DNA-MS and productivity determinants, the character of correlations depends on breed of animals.
Текст научной статьи Сравнительный анализ информативности эритроцитарных антигенов и ДНК-микросателлитов как генетических маркеров в селекционно-племенной работе со свиньями канадской селекции
Грамотно сформированная линейная структура стада является одним из ключевых элементов эффективного ведения селекционноплеменной работы на свиноводческих предприятиях — племзаводах, племрепродукторах, в генофондных хозяйствах, селекционногибридных центрах. Практикуемая достаточно часто система ротации по поколениям при получении хряков и свиноматок приводит к тому, что генетические различия между линиями стираются. Поэтому эффект от межлинейных ротационных кроссов в поколениях снижается, что вызывает необходимость завоза генетически разнородного племенного материала из других предприятий России или из-за рубежа с целью «освежения крови».
Не отрицая периодическую необходимость подобных биотехнических мероприятий, следует отметить ряд преимуществ замкнутого разведения: возможность прогнозирования племенной ценности потомства, управление степенью инбридинга и консолидацией линий, снижение риска привнесения инфекционных заболеваний. Однако очевидно, что разведение животных в замкнутых популяциях предъявляет повышенные требования к системе селекционно-племенной работы.
В качестве инструмента контроля и управления структурой стада рассматривается использование генетических маркеров — эритроцитарных антигенов и ДНК-микросателлитов. Генетические маркеры позволяют судить о степени гетерозиготности животных, консолидации наследственных качеств пород, типов, линий, а также генетических расстояниях между ними (1, 2). Имеются многочисленные данные о связи полиморфных вариантов локусов групп крови (B, G, E, F, K, L) свиней с признаками продуктивности (3-5) . В ряде работ показано, что эти корреляции довольно сложные, а в большинстве случаев неоднозначные (6-10). Подавляющее большинство исследований посвящено выявлению связи отдельных аллелей ЭА с признаками продуктивности.
Целью настоящей работы было определение информативности двух систем анализа генетического полиморфизма — эритроцитарных антигенов (ЭА) и ДНК-микросателлитов (ДНК-МС) для характеристики структуры стад и оценки гетерогенности подбора родительских пар по признакам продуктивности у свиней трех пород канадской селекции.
Методика . Исследования проводили на трех группах свиней пород канадской селекции: ландрас ( n = 38), дюрок ( n = 55), йоркшир ( n = 73) (ООО «Троснянский бекон», Орловская обл.). При отборе проб крови использовали гепарин в качестве антикоагулянта, кожи (в случае длительной транспортировки) — 96 % этиловый спирт в качестве консерванта. Группы крови определяли по 20 ЭА 10 систем групп крови согласно инструкции (11), применяли иммуноспецифические сыворотки производства Всероссийского НИИ племенного дела.
ДНК выделяли в соответствии с методическими рекомендациями (12). Для анализа ДНК-МС профиля использовали методику, включающую проведение двух мультиплексных полимеразных цепных реакций (ПЦР), которая позволяет амплифицировать фрагменты 12 аутосомальных ДНК-МС маркеров. Набор ДНК-МС был выбран в соответствии с рекомендациями Международного общества генетиков (ISAG). Первичную обработку данных капиллярного электрофореза выполняли с помощью программного обеспечения MegaBACE. Данные об аллелях каждого животного суммировали в электронной таблице Microsoft Excel. Полученная таким образом матрица генотипов служила основой для статистической обработки результатов, которую осуществляли по Б. Вейру (13) с использованием программ MSA_WIN v 2.65, Phylip, TreeView. Расчет генетических расстояний и построение дендрограмм проводили по M. Nei (14).
Результаты. Число аллелей в локусах ЭА у исследуемых групп животных варьировало от 1 до 4 и в среднем составляло 2,50±0,22 при наибольшем — у свиней породы ландрас (2,85±0,24) и наименьшем — у свиней породы йоркшир (2,50±0,16). Число аллелей ДНК-МС в зависимости от локуса варьировало от 3 до 11 и в среднем составляло 5,91±0,42, что свидетельствует о более высоком уровне полиморфизма ДНК-МС. Как и по группам крови, по ДНК-МС наибольшим числом аллелей в локусе характеризовались свиньи породы ландрас (6,58±0,62), наименьшим — животные породы йоркшир (5,33±0,37). Таким образом, использование в качестве маркеров ЭА и ДНК-МС давало сходную картину относительных различий по среднему числу аллелей между исследуемыми группами свиней. При этом в абсолютном выражении среднее число аллелей на микросателлитный локус было в 2,4 раза выше, чем на локус систем групп крови, что свидетельствует о большей информативности ДНК-МС.
1. Фактическая (Het ф акт . ) и ожидаемая
(Hetmeop) степень гетерозиготности в группах свиней разных пород, рассчитанная на основании генетического полиморфизма эритроцитарных антигенов (ЭА) и ДНК-микросателитов (ДНК-МС)
|
Порода ( n ) |
Показатель |
ЭА |
ДНК-МС |
|
Дюрок |
Het факт. |
0,411±0,132 |
0,486±0,194 |
|
( n = 55) |
Het теор. |
0,326±0,078 |
0,623±0,133 |
*
Работа выполнена при финансовой поддержке РФФИ, грант № 06-04-08196.
Het факт. -Het теор.
Ландрас Het факт.
( n = 38) Het теор.
Het факт. -Het теор.
Йоркшир Het факт.
( n = 73) Hetтеор.
Het факт. -Het теор.
+8,48 0,413±0,132 0,315±0,091
+9,84 0,569±0,119 0,420±0,059 +14,91
–13,66 0,552±0,201 0,721±0,071
–16,93 0,622±0,093 0,735±0,075
–11,30
Из сравнения фактической и ожидаемой степени гетерозиготности в группах (табл. 1) следует, что показатели, рассчитанные с использованием двух систем анализа, одинаково отражали характер распределения исследуемых групп по неоднородности: наиболее гетерогенными оказались свиньи породы йоркшир, наименее — породы дюрок. Однако абсолютная величина показателей при использовании двух аналитических систем существенно различалась: при оценке по ЭА фактический уровень гетерозиготности по группам варьировал от 41,1 (порода дюрок) до 56,9 % (порода йоркшир), по ДНК-МС — соответственно от 48,6 до 62,2 %.
Сравнение фактической и теоретически ожидаемой степени гетерозиготности выявило противоположные тенденции. Так, во всех исследованных группах по ЭМ-маркерам наблюдалось превышение уровня гетерозиготности над теоретически ожидаемым (от 8,5 до 14,9 % соответственно в группах свиней пород дюрок и йоркшир).
При использовании в качестве критерия оценки ДНК-МС у всех животных отмечался дефицит гетерозигот: от 11,3 (порода йоркшир) до 16,9 % (порода ландрас). Такие различия объясняются неодинаковой полиморфностью используемых аналитических систем. Учитывая, что в исследованиях использовались линейные свиньи, полученные с применением умеренного инбридинга, выявленный дефицит гетерозигот более объективен с точки зрения селекции.
При оценке генетических различий между генеалогическими группами внутри пород на основании генетических расстояний для группы животных породы йоркшир следует отметить некоторые различия в формирующейся кластерной структуре генеалогического дерева, построенного с использованием двух систем анализа: по ЭА наибольшим генетическим родством характеризовались генеалогические группы Спартак и Фауна, наименьшим — Сателлит и Спикер, Спикер и Флора; по ДНК-МС более тесные родственные связи отмечались между семействами Флора и Фауна и линиями Сателлит и Спартак (рис., А).

---------Спикер
Сателлит
Спартак
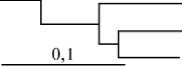
— Флора Фауна
б
Фасоль
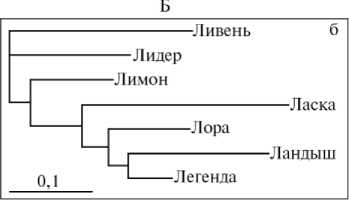
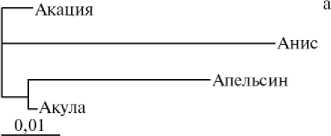
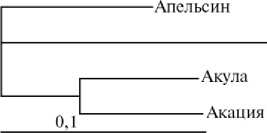
Дендрограмма генеалогических групп свиней породы йоркшир (А) , ландрас (Б) и дюрок (В) при оценке генетических расстояний на основании полиморфизма по локусам эритроцитарных антигенов (а) и ДНК-микросателитам (б) .
Проявилась также консолидированность семейств свиноматок и линий хряков. Кластерная структура генеалогического дерева свиней породы йоркшир, построенного на основании анализа ДНК-МС, полностью согласуется с данными о происхождении животных в соответствии со сведениями по племенному учету, а также с используемой системой подбора на племпредприятии. Кроме того, диапазон числовых значений генетических различий, полученный на основе анализа ДНК-МС, более информативен.
Расчет генетических дистанций на основании результатов анализа ЭА у свиней породы ландрас (см. рис., Б) выявил минимальные различия, которые не позволили сделать вывод о характере генеалогических связей между линиями и семействами. Анализ, проведенный на основе данных ДНК-МС, показал, что наибольшей удаленностью характеризовались животные линии Ливня и семейства Ласки, более близким генеалогическим родством — генеалогические группы Лора—Легенда, Ландыш—Легенда. Кластерная структура генеалогического дерева в группе свиней породы ландрас, построенного на основании результатов анализа ДНК-МС, полностью соответствует данным племенного учета на предприятии.
У животных породы дюрок (см. рис., В) обнаружены некоторые различия в характере связей между линиями и семействами в зависимости от используемой системы анализа. Кластерная структура дендрограммы, построенной на основании анализа ДНК-МС (семейства Акулы и Акации, с одной стороны, и линии Аниса и Апельсина, — с другой, попарно формируют два кластера), полностью согласуется с данными о происхождении животных.
Таким образом, информативность ДНК-МС для характеристики генеалогической структуры групп свиней пород йоркшир, ландрас и дюрок канадской селекции выше. Меньшая маркерная информативность ЭА и, как следствие, более низкая чувствительность к небольшим различиям между группами в некоторой степени искажают генеалогические связи между внутрипородными структурами (линиями и семействами) исследуемых пород.
На следующем этапе исследований при оценке влияния степени гетерогенности при подборе пар, рассчитанной по результатам анализа полиморфизма ЭА и ДНК-МС, на признаки продуктивности свиней выявили ряд достоверных корреляций (табл. 2).
Следует отметить, что корреляции в группах свиней материнских пород (йоркшир и ландрас) носили одинаковый и отличный от выявленных у свиней отцовской породы дюрок характер. Так, у свиней материнских пород высокие достоверные положительные корреляции (P > 0,9999) наблюдались между гетерогенностью подбора и средней массой поросенка в 21-суточном возрасте и при отъеме, причем при использовании в качестве критерия оценки гетерогенности ДНК-МС сила корреляций была выше, ЭА — ниже. Отрицательные корреляции отмечены по количественным показателям плодовитости — общему числу поросят при рождении и числу живых поросят при рождении.
У свиней породы дюрок выявлено значительно больше достоверных корреляционных зависимостей, однако в основном они были противоположными таковым у животных материнских пород. Сильные положительные корреляции наблюдались между степенью гетерогенности, рассчитанной по ЭА и ДНК-МС, и числом поросят при рождении (в том числе живых), сильные отрицательные — по средней массе поросенка в возрасте 21 сут. По показателям массы гнезда при рождении и числа голов через 21 сут корреляции, рассчитанные с использованием ЭА и ДНК-МС, носили противоположный характер.
Выявление влияния гетерогенности группового подбора на продуктивность имеет важное практическое значение. Вместе с тем более четкие выводы о роли генетических различий родителей в проявлении признаков продуктивности могут быть сделаны при проведении индивидуальных подборов пар. При проведении таких подборов отмечали больше корреляционных зависимостей между степенью гетерогенности и признаками продуктивности (см. табл. 2).
2. Корреляции между генетическими дистанциями и признаками продуктивности у свиней пород йоркшир, ландрас и дюрок при разных способах подбора по гетерогенности
|
Признак |
Йоркшир |
Ландрас |
Дюрок |
|
ЭА ДНК-МС |
ДНК-МС |
ЭА ДНК-МС |
|
П о д б о р л и н и я |
х с е м е й с т в о |
||||
|
Родилось поросят: |
|||||
|
всего |
–0,25±0,099* |
–0,37±0,091 |
–0,36±0,167*** |
0,71±0,079 |
0,93±0,021 |
|
живых |
–0,23±0,100** |
–0,36±0,167* |
0,66±0,090 |
0,98±0,005 |
|
|
Число поросят: |
|||||
|
в 21-суточном |
|||||
|
возрасте |
0,39±0,135* |
–0,28±0,147*** |
|||
|
при отъеме |
0,35±0,135* |
||||
|
Крупноплодность |
0,37±0,138* |
0,41±0,133* |
|||
|
Живая масса поросенка: |
|||||
|
в 21-суточном воз- |
|||||
|
расте |
0,60±0,067 |
0,96±0,007 |
0,61±0,121 |
–0,91±0,027 |
–0,61±0,101 |
|
при отъеме |
–0,32±0,095 |
0,72±0,051 |
0,63±0,114 |
–0,46±0,126 |
|
|
Масса гнезда: |
|||||
|
при рождении по- |
|||||
|
росят |
–0,21±0,101*** |
0,95±0,013 |
–0,46±0,125 |
||
|
в 21-суточном воз- |
|||||
|
расте |
0,31±0,096* |
–0,31±0,173**** |
–0,60±0,102 |
||
|
при отъеме |
–0,17±0,102**** |
–0,37±0,165*** |
0,25±0,150**** |
||
|
П о д б о р х р я к х |
с в и н о м а т к а |
||||
|
Родилось поросят: |
|||||
|
всего |
–0,27±0,098* |
–0,30±0,108* |
0,98±0,005 |
–0,51±0,097 |
0,94±0,015 |
|
живых |
–0,99±0,000 |
–0,99±0,001 |
0,36±0,184**** |
–0,37±0,139* |
|
|
Число поросят: |
|||||
|
в 21-суточном воз- |
|||||
|
расте |
–0,83±0,031 |
–0,85±0,032 |
–0,90±0,030 |
||
|
при отъеме |
–0,83±0,031 |
–0,85±0,032 |
–0,51±0,097 |
–0,96±0,011 |
|
|
Крупноплодность |
0,83±0,031 |
0,85±0,032 |
0,86±0,053 |
–0,98±0,004 |
–0,96±0,011 |
|
Живая масса поросенка: |
|||||
|
в 21-суточном воз- |
|||||
|
расте |
0,68±0,056 |
0,85±0,033 |
–0,59±0,137 |
–0,43±0,131* |
|
|
при отъеме |
0,90±0,018 |
0,63±0,071 |
–0,48±0,100 |
–0,99±0,001 |
|
|
Масса гнезда: |
|||||
|
при рождении по- |
|||||
|
росят |
0,78±0,040 |
0,77±0,048 |
0,42±0,174*** |
–0,94±0,014 |
–0,79±0,060 |
|
в 21-суточном воз- |
|||||
|
расте |
0,99±0,002 |
0,98±0,003 |
0,36±0,184**** |
–0,73±0,075 |
|
|
при отъеме |
–0,20±0,102*** |
–0,98±0,003 |
–0,99±0,001 |
–0,87±0,030 |
–0,99±0,001 |
П р и м е ч а н и е. Для расчета генетических расстояний использовали маркерные системы эритроцитарных антигенов
(ЭА) и ДНК-микросателлитов (ДНК-МС), Р > 0,999.
* P > 0,99; ** P > 0,98; *** P > 0,95; **** P > 0,90.
Так, у свиней породы йоркшир наблюдали достоверные отрицательные связи с количественными показателями плодовитости: числом поросят при рождении (в том числе живых), числом поросят через 21 сут и при отъеме. Характер зависимостей, выявленных с использованием в качестве критериев оценки гетерогенности ЭА и ДНК-МС, был одинаковым.
У свиней породы ландрас имела место сильная достоверная положительная корреляция с общим числом поросят при рождении; корреляция с числом живых поросят при рождении также была положительной, но менее выраженной. У животных породы дюрок количественные показатели плодовитости (число живых поросят при рождении, в возрасте 21 сут и при отъеме) отрицательно коррелировали с гетерогенностью подбора.
Итак, ЭА и ДНК-МС могут служить инструментами для характеристики структуры стад свиней трех изученных пород канадской селекции, однако при небольших генетических различиях между внутрипородными генеалогическими структурами ДНК-МС как маркеры более информативны. Выявлен ряд достоверных корреляционных зависимостей между степенью гетерогенности подбора родительских пар
(групповых — линия х семейство и индивидуальных — хряк % свиноматка), оцененной по ЭА и ДНК-МС, и признаками продуктивности свиней. В большинстве случаев корреляции носят одинаковый характер независимо от используемой системы анализа. Показано влияние породной принадлежности животных на силу и направленность корреляций. Наиболее четкие зависимости установлены в группе свиней породы йоркшир. Увеличение степени гетерогенности подбора приводит к достоверному снижению количественных показателей продуктивности и повышению показателей массы. В группе свиней породы ландрас выявленные корреляции зачастую носили противоположный характер. В группе свиней породы дюрок прослеживается зависимость снижения показателей массы с увеличением степени гетерогенности при подборе пар. На основании выявленных нами закономерностей могут разрабатываться системы генетического контроля структуры стад и управления процессом селекционно-племенной работы в свиноводстве.
Л И Т Е Р А Т У Р А
-
1. Н о в и к о в А.А., Р о м а н е н к о Н.И. Создание новой линии свиней с использованием генетических маркеров. В сб.: Селекция, кормление, содержание сельскохозяйственных животных и технология производства продуктов животноводства. Лесные Поляны, 1999, 7: 56-60.
-
2. З и н о в ь е в а Н.А., Э р н с т Л.К. Проблемы биотехнологии и селекции сельскохозяйственных животных. Изд. 2-е, доп. Дубровицы, 2006.
-
3. Т и х о н о в В.Н., Н и к и т и н С.В. Связь системы B групп крови с многоплодием и жизнеспособностью свиней. В сб. науч. тр. Новосибирского СХИ. Новоси
бирск, 1984: 110.
-
4. Н о в и к о в А.А., К о з и н И.Е., Р о м а н е н к о Н.И. Дифференциация крупной белой породы свиней по генетической структуре групп крови в связи с хозяйственно полезными признаками. В сб. науч. тр. ВНИИплем. Лесные Поляны, 2000, вып. 10: 87-91.
-
5. С е р д ю к Г.Н. Иммуногенетика свиней: теория и практика. СПб, 2002.
-
6. Б е з е н к о С.П. О направлениях иммуногенетики в селекции свиней. Свиноводство, 1982, 5: 25-26.
-
7. Б е з е н к о С.П., М и л ь ч е в с к а я Р.И., А л ь к е й с и Т.В. Методическое пособие по определению генетических и иммунологических характеристик свиней для
использования в селекции. Дубровицы, 2006: 36-40.
-
8. Т о л п е к о Г.А. Формирование иммуногенетической структуры популяции свиней в связи с методами разведения и отбором по продуктивности. Автореф. докт. дис. Новосибирск, 1985.
-
9. С о л д а т е н к о в Н.К. Использование иммуногенетических тестов для повышения эффективности селекционно-племенной работы в промышленных комплексах. Автореф. канд. дис. СПб, 1992.
-
10. Г а р а й В.В., Е п и ш и н В.А., Н и к о л а е в а И.В. Количественная оценка показателей продуктивности племенных стад. В сб.: Современные проблемы развития свиноводства. Жодино, 2000: 40-42.
-
11. Сыворотки иммуноспецифические для определения групп крови свиней. Технические условия, наставление и инструкция по изготовлению, контролю и применению. Дубровицы, 2001.
-
12. З и н о в ь е в а Н.А., П о п о в А.Н., Э р н с т Л.К. и др. Методические рекомендации по использованию метода полимеразной цепной реакции в животноводстве. Дубровицы, 1998.
-
13. В е й р Б. Анализ генетических данных. М., 1995.
-
14. N e i M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, 1978, 89: 583-590.
ООО «Агрофест-Орел»,
