Модели оценки комбинационной способности привоев и подвоев для прогноза урожайности у привитых деревьев яблони

Автор: Дьяков А.Б., Драгавцева И.А., Ефимова И.Л., Доможирова В.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Цитогенетические и математические методы в селекции растений
Статья в выпуске: 5 т.49, 2014 года.
Бесплатный доступ
Вегетативное размножение лучших сортов плодовых культур прививкой на клоновые подвои обеспечивает однородность деревьев, раннее наступление плодоношения, позволяет создавать уплотненные насаждения. С целью выявления подвоя, обеспечивающего максимальную урожайность привитого на нем сорта, проводятся дорогостоящие, длящиеся десятки лет испытания большого числа сортоподвойных комбинаций. В многочисленных публикациях сообщались в основном результаты такого эмпирического подбора. Поэтому до сих пор нет ни теории, ни методов прогноза урожайности для привитых деревьев по показателям привоев и подвоев. Повысить эффективность таких поисков и снизить затраты средств и времени на их проведение можно лишь на основе теоретических разработок принципов прогнозирования признаков у привитых деревьев по характеристикам привоев и подвоев. Исследования авторов статьи направлены на выяснение особенностей использования принципов и математических моделей биометрической генетики для познания связей в системе прививочные компоненты-привитое растение. Реализация этой программы была начата с изучения возможностей применения формул для вычисления комбинационной способности привоев и подвоев как меры их влияния на количественные признаки получаемых комбинаций. Экспериментальным материалом служили результаты испытания 28 комбинаций (4 сорта яблони - Джонатан, Голден Делишес, Айдаред и Корей на 7 подвоях - 1-48-1, 1-47-55, 1-48-46, М2, М3, М4 и М7) по урожайности, усредненные за 21 год плодоношения. Установлено, что генотипы привоев различаются как по урожайности, усредненной по всем подвоям, так и по степени зависимости этого признака от конкретных подвоев, а подвои неодинаково дифференцируют привои по фенотипическому проявлению урожайности. Широко используемые метод и математические модели оценки комбинационной способности у родительских линий F 1 гибридов можно с успехом применять для определения участия общей и специфической комбинационной способности генотипов привоев и подвоев во влиянии на урожайность привитых деревьев. Основанием служит тот факт, что в структуре вариации комбинационной способности прививочных компонентов преобладает доля общей комбинационной способности, которая (по аналогии с родительскими формами гибридов) представляет собой функцию аддитивных генов. Вычисления на основе формул биометрической генетики показали, что общая комбинационная способность прививочных компонентов в 6,4 раза больше их специфической комбинационной способности. Это обусловило высокую эффективность прогнозов по урожайности привитых деревьев на основании общей комбинационной способности привоев и подвоев (отмечен высокий коэффициент корреляции между фактическими и прогнозируемыми оценками урожайности - r = 0,930). Тесная корреляция между прогнозируемыми по общей комбинационной способности и фактическими оценками урожайности привойно-подвойных комбинаций свидетельствует о высокой степени интегрированности свойств прививочных компонентов в определении количественных признаков привитого дерева. Предварительный анализ выявил возможность еще большего повышения точности таких прогнозов за счет вычленения из дисперсий оценок специфической комбинационной способности линейного компонента.
Модели, привой, подвой, комбинационная способность, прогноз урожайности
Короткий адрес: https://sciup.org/142134835
IDR: 142134835 | УДК: 634:631.541.1:51-7
The models for estimation of a combining ability of varieties and rootstocks to forecast yielding in apple trees
Vegetative reproduction of the best varieties of fruit crops by grafting on clonal rootstocks provides uniform trees, early onset of fruiting, it allows to create dense plantings. In order to identify rootstock, which gives maximum crop yield of the grafted varieties, a large number of expensive tests of cultivar-rootstock combinations, lasting dozens of years, are carried out, and the results of such empirical selection were reported in numerous publications. Therefore, so far there is no theory or methods of forecasting yields for trees grafted on indicators of variety and rootstocks. To increase efficiency of such searches and to lower expenses and time for their carrying out, there is the only possible way based on theoretical researches targeted to development of the principles of prediction of signs of grafted trees according to characteristics of varieties and rootstocks. Our researches summarized herein, are curried out to clarify the features of use of the principles and mathematical models of biometric genetics for knowledge of communications in a system «grafted components-grafted plant». The implementation of this program started with studying possible applications of the formulas for calculation of the combinational ability of varieties and rootstocks as measures of their influence on the quantitative signs of the resulting combinations. The data on yielding in 28 such combinations (4 grades of apple, namely Jonathan, Golden Delicious, Idared, and Korah, and 7 rootstock, 1-48-1, 1-47-55, 1-48-46, M2, M3, M4 and M7), averaged up for 21 years fruiting, were used as the experimental material. It is established that the genotypes of varieties differ on both the productivity averaged for all rootstocks and the degree of dependence of this sign from the rootstocks. Besides, the rootstocks unequally differentiate varieties according to phenotypic manifestation of productivity. The widely used method and mathematical models for an assessment of combining ability of the parental lines of F 1 hybrids can be successfully applied to estimate the influence of common and specific combining ability of the varieties and rootstocks on crop yield in grafted trees. It is based on the fact that in variation of the total combining ability of grafting components the general combining ability (GCA) as a function of additive genes, similar to that observed in the parental forms of hybrids, is relatively prevalent. When formulas of biometric genetics are used, the calculations show that the general combining ability (GCA) of grafting components is 6.4 times more than their specific combining ability (SCA). It caused high efficiency of the forecasts on productivity of grafted trees based on the GCA of varieties and rootstocks. The high coefficient of correlation between the actual and predicted estimates of productivity has been noted ( r = 0,930). Close correlation between the predicted on the GCA and actual estimates of productivity of the variety-rootstock combinations shows the high degree of integration of grafting components in the manifestation of quantitative sings of a grafted tree. The preliminary analysis revealed the possibility for further improving accuracy of such forecasts due to separating the linear component from the dispersions of the specific combinational ability (SCA) estimates.
Текст научной статьи Модели оценки комбинационной способности привоев и подвоев для прогноза урожайности у привитых деревьев яблони
Современное промышленное садоводство основано на вегетативном размножении лучших сортов плодовых культур прививкой на клоновые подвои (1, 2). Это обеспечивает однородность деревьев, раннее наступление у них плодоношения, позволяет создавать уплотненные насаждения. Очень важен также учет влияния подвоев на урожайность сортов. В случаях удачного выбора подвоев для конкретных сортов яблонь и груш их урожайность удается повысить за счет комбинационного эффекта в 1,5-2,0 раза (3-6).
Работа поддержана грантом РФФИ № 13-01-96-519.
С целью выявления подвоя, обеспечивающего максимальные оценки урожайности привитого на нем сорта, проводятся дорогостоящие, длящиеся десятки лет эмпирические испытания большого числа сортоподвойных комбинаций. Повысить эффективность таких поисков и снизить затраты средств и времени на их проведение можно лишь на основе теоретических исследований, направленных на разработку принципов и математических моделей прогнозирования признаков привитых деревьев по характеристикам привоев и подвоев (7-9).
Попытки решения этой задачи предпринимались давно. Еще Ф. Кобель (10) писал, что причины неодинакового поведения разных подвоев трудно выяснить и вопрос этот до сих пор остается неизученным, а крупнейший специалист по трансплантации растений Н.П. Кренке (11) констатировал невозможность подбирать подвой на основе чисто теоретических рассуждений. Позже Й. Шмадлак (1), описывая сложности взаимовлияния прививочных компонентов, заключил, что знания о таких взаимосвязях еще очень незначительны. Не была решена эта задача и в последующие годы. В многочисленных публикациях сообщались в основном результаты эмпирического подбора привойно-подвойных комбинаций, поэтому до сих пор нет ни теории, ни методов прогноза урожайности привитых деревьев по показателям привоев и подвоев.
Результаты научных исследований и многовековая практика выращивания привитых деревьев свидетельствуют о том, что урожайность привойно-подвойных комбинаций определяется генотипами как привоев, так и подвоев. Поэтому изучение характера взаимодействия прививочных компонентов в формировании свойств привитого дерева должно быть основано на принципах генетики. Особенности взаимовлияния компонентов прививок изучались в течение ряда десятилетий (12-14). Однако эти исследования проводились только для выяснения возможности наследования изменений по признакам одного компонента прививки под влиянием другого. Генетики даже не ставили задачу прогнозирования таких изменений у привитых растений, так как было доказано, что они не наследуются, представляют собой «временные модификации» (13) и, следовательно, обозначенная проблема представлялась для генетики неактуальной. Однако подобные модификации временны лишь в том смысле, что они сохраняются до тех пор, пока один прививочный компонент воздействует на другой, но не передаются семенному потомству. При этом С.Я. Краевой (14) на основании результатов своих опытов сделал вывод об адекватности таких модификационных изменений привоев генотипическим особенностям их подвоев. Для практики садоводства явление специфичности влияния конкретного генотипа подвоя на признаки привитого на него сорта плодовой культуры имеет чрезвычайно большое значение, так как служит генетической предпосылкой для прогнозов наиболее эффективных привойно-подвойных комбинаций. Высокая урожайность у лучших из таких комбинаций представляет собой временную модификацию с точки зрения генетики, но она постоянна для садовода, поскольку он выращивает только привитые деревья и не размножает сорта семенами.
При изучении изменений, обусловленных взаимовлиянием привоев и подвоев, генетикам требовались объекты, различающиеся по качественным признакам. Например, С.Я. Краевой (14) показал, что при прививках на табак или дурман в побегах томата вместо алкалоида соланина синтезировались соответственно никотин или атропин, а культурный сорт картофеля в качестве привоя индуцировал клубнеобразование у подвоя дикого вида Demissum. Однако для практики садоводства важны количествен- ные изменения урожайности, адаптивности и других полигенных признаков у привитых деревьев (15). Такую изменчивость должна изучать биометрическая генетика, но даже в обстоятельной основополагающей монографии К. Мазера и Дж. Джинкса (16) проблемы прививок совсем не рассматриваются. По мнению авторов монографии, обычно коррелятивные реакции обусловлены не плейотропией, а сцеплением генов. Хотя подобные реакции привоя на воздействие подвоев нельзя объяснить сцеплением генов, при изучении взаимовлияния прививочных компонентов необходимо использовать принципы и математические модели биометрической генетики, в первую очередь формулы оценки комбинационной способности привоев и подвоев как меры их влияния на количественные признаки привойно-подвойных комбинаций.
Длительность периодов от всходов до первого цветения и плодоношения плодовых культур ограничивает проведение рекогносцировочных скрещиваний для генетического изучения исходных форм и выбора среди них перспективных доноров до выполнения селекционных программ (17). Несмотря на это, такие эксперименты проводятся как с целью анализа структуры генотипических дисперсий (18-20), так и для оценок общей и специфической комбинационной способности компонентов скрещиваний (21-23). Однако исследователи, изучавшие наследственную изменчивость полигенных признаков плодовых культур, не ставили задачу использования принципов и моделей биометрической генетики для изучения влияния привоев и подвоев на урожайность и другие признаки привитых деревьев. Не было даже попыток использовать формулы вычисления комбинационной способности компонентов скрещиваний для оценок комбинационной способности прививочных компонентов, хотя привитое дерево представляет собой целостный организм (1, 2, 11, 12), в какой-то мере подобно тому, как гибридное растение F 1 является целостным организмом, несущим наследственные задатки гамет двух родителей.
Цель представляемой работы заключалась в изучении эффективности применения математических моделей биометрической генетики для оценок общей и специфической комбинационной способности привоев и подвоев при прогнозировании урожайности их комбинаций, а также для получения сведений о степени интегрированности свойств прививочных компонентов в признаках привитых деревьев.
Методика . Испытания проводили на четырех сортах яблони (Джонатан, Голден Делишес, Айдаред и Корей — районированные сорта зимнего срока созревания, имеют сходный тип дерева — ветвистое с раскидистой формой кроны), каждый из которых был привит на семи подвоях (1-48-1, 1-47-55, 1-48-46, М2, М3, М4 и М7) (ОПХ «Центральное» Северо-Кавказского НИИ садоводства и виноградарства — СКЗНИ-ИСиВ, г. Краснодар; сад посадки 1977 года). Используемые в опыте подвои представляли группу среднерослых (за исключением полукарликового подвоя М7; подвои М2, М3, М4 и М7 — английской селекции, подвои 148-1, 1-47-55, 1-48-46 — селекции СКЗНИИСиВ).
Анализируемые привойно-подвойные комбинации яблони были высажены по одной схеме посадки (7x5 м). За годы исследований в насаждении применялись однотипные технологии ухода за почвой, кроной и защиты деревьев от болезней и вредителей. Погодные условия за период исследований были в основном типичными для зоны возделывания. В отдельные годы имели место различные погодные стрессоры.
Урожайность (xik) каждого i-го сорта на k-м подвое усреднили за 21 год плодоношения для сведения к минимуму случайных ошибок при выполнении оценки.
Показатели комбинационной способности прививочных компонентов и прогнозируемые оценки урожайности привитых деревьев вычисляли с помощью формул биометрической генетики (24-26). Результаты подвергали корреляционному и регрессионному анализам по всей совокупности данных и по стратифицированным расслоенным выборкам. Для определения достоверности выявленных зависимостей между прогнозируемыми и фактическими оценками урожайности сравнивали вычисленные коэффициенты корреляции с их критическими величинами при разных уровнях существенности с учетом числа степеней свободы и тесноты зависимостей.
Результаты . Влияние проявившихся за период наблюдений погодных стрессоров нашло отражение в урожайности деревьев как интегрального показателя адаптивности и продуктивности многолетних плодовых растений.
Проведенные испытания выявили большие различия по урожайности у привойно-подвойных комбинаций — от x ik = 122,7 ц/га у сорта Джонатан на подвое 1-47-55 до x ik = 241,6 ц/га у сорта Корей на подвое М4 (табл. 1). Общая изменчивость оказалась в большей степени обусловленной генотипическими различиями привоев, чем подвоев. Была обнаружена также специфичность влияния отдельных подвоев и привоев: наибольшую способность дифференцировать сорта отмечали у подвоя 1-47-55, наименьшую — у М4 (разница между максимальной и минимальной урожайностью сортов соответственно 80,2 и 46,4 ц/га), а степень реакции сортов на генотипические особенности подвоев была наибольшей у сорта Джонатан, наименьшей — у Годден Делишес (подобная разница составила соответственно 72,5 и 26,5 ц/га). Неодинаковая способность подвоев дифференцировать сорта по урожайности и разная реакция сортов на особенности подвоев дают основание считать, что испытанные прививочные компоненты различаются по комбинационной способности.
1. Средняя урожайность (x ik , ц/га) у разных генотипов привойно-подвойных комбинаций яблони (ОПХ «Центральное» СКЗНИИСиВ, г. Краснодар, 1982-2002 годы)
|
Привой, сорт |
Подвой, форма |
Среднее, x i . |
||||
|
1-48-1 | |
1-47-55 | 1-48-46 | М2 | |
М3 1 |
М4 |
| М7 |
||
|
Джонатан |
129,3 |
122,7 173,1 169,2 |
158,8 |
195,2 |
146,7 |
156,4 |
|
Голден Делишес |
187,4 |
202,9 204,4 213,9 |
202,0 |
211,3 |
187,4 |
201,3 |
|
Айдаред |
196,4 |
186,7 232,7 203,6 |
187,5 |
217,4 |
185,3 |
201,4 |
|
Корей |
189,9 |
166,5 235,0 232,5 |
234,5 |
241,6 |
212,2 |
216,0 |
|
Среднее, x. к |
175,8 |
169,7 211,3 204,8 |
195,7 |
216,4 |
182,9 |
x.. = 193,8 |
|
Примечание. x , . — среднее конкретного сорта на каждом конкретном подвое, x.. — среднее по всем комбинациям. |
подвое, |
x. к — |
среднее каждого сорта на |
|||
Для оценки генетических свойств родительских форм по результатам испытаний их гибридов в биометрической генетике предлагаются два параметра — общая и специфическая комбинационная способность (24). О смысле этих показателей можно судить по их описанию В.К. Савченко (25), согласно которому под общей комбинационной способностью (ОКС) понимается средняя ценность родительской линии в гибридных комбинациях, а под специфической комбинационной способностью (СКС) — случаи, когда конкретные комбинации оказываются лучше или хуже, чем можно было ожидать на основе среднего качества линий, участвующих в скрещиваниях. Основой анализа комбинационной способности служит математическая модель, вариант которой для полной схемы диаллельных скрещиваний Л.В. Хотылева и Л.А. Тарутина (26) представили, как 58
x ik = X.. + g i + g k + S ik + r ik + e ik , [1] где x ik — оценка каждого гибрида от скрещивания i -го и k -го родителей; х.. — средняя величина признака у гибридов всех комбинаций скрещиваний; g i — эффект ОКС i -го родителя; g k — эффект ОКС k -го родителя; Sik — эффект СКС i -го и k -го родителей; r ik — реципрокный эффект; e ik — случайное отклонение оценок для каждого гибрида.
Результаты диаллельных скрещиваний позволяют проводить детальный генетико-статистический анализ структуры наследственной изменчивости по количественным признакам растений, если выполняется ряд требований к подбору экспериментального материала. При оценке комбинационной способности генотипов подобные требования к подбору изучаемых объектов не предъявляются, что делает возможным более широкое применение упомянутого метода (26). Это обстоятельство также позволяет допустить, что указанный метод и соответствующие математические модели могут применяться для вычислений при оценке комбинационной способности генотипов у привоев и подвоев по результатам испытаний урожайности привойно-подвойных комбинаций плодовых деревьев.
При использовании с этой целью уравнения [1] следует учесть, что реципрокные прививки не имеют смысла, поэтому r ik надо исключить из слагаемых формулы. Представленные в таблице оценки урожайности каждой привойно-подвойной комбинации получены при усреднении результатов учетов за 21 год плодоношения привитых деревьев, поэтому величины случайных отклонений оценок e ik минимальны, что позволяет условно пренебречь ими для упрощения вычислений и проводить анализ комбинационной способности привоев и подвоев на основе формулы:
x ik = х.. + g i + g k + S ik , [2] причем мы принимаем g i как эффект ОКС i привоя, а g k — как эффект ОКС k подвоя. Для вычислений величин g i и g k используются данные таблицы 1 и формулы:
g i = x i . - x.., [3]
g k = x. k - x.., [4] то есть оценка эффекта ОКС i i -го привоя равна его средней урожайности на всех подвоях за вычетом значения средней урожайности всех изученных в опыте привойно-подвойных комбинаций. Аналогично вычисляется и ОКС k каждого k -го подвоя.
2. Эффекты общей комбинационной способности привоев (g i , ц/га) и подвоев (g k , ц/га) и прогнозируемые по их значениям оценки урожайности (x ik , ц/га) привойно-подвойных комбинаций яблони (ОПХ «Центральное» СКЗНИИСиВ, г. Краснодар, 1982-2002 годы)
|
Привой, сорт |
Подвой, форма |
g i |
||||||
|
1-48-1 |
1-47-55 |
1-48-46 | |
М2 |
М3 |
М4 |
М7 |
||
|
Джонатан |
138,4 |
132,3 |
173,9 |
167,4 |
158,3 |
179,0 |
145,5 |
- 37,4 |
|
Голден Делишес |
183,3 |
177,2 |
218,8 |
212,3 |
203,2 |
223,9 |
190,4 |
7,5 |
|
Айдаред |
183,4 |
177,3 |
218,9 |
212,4 |
203,3 |
224,0 |
190,5 |
7,6 |
|
Корей |
198,0 |
191,9 |
233,5 |
227,0 |
217,9 |
238,6 |
205,1 |
22,5 |
|
g k |
- 18,0 |
- 24,1 |
17,5 |
11,0 |
1,9 |
22,6 |
- 10,9 |
|
Чтобы выяснить, в какой мере методика и математические модели определения комбинационной способности родительских форм гибридов F1 применимы для оценок комбинационной способности привоев и подвоев, а также установить возможность прогноза урожайности привитых деревьев по величинам ОКС и СКС прививочных компонентов, необходимо вычислить эти величины, рассчитать по этим показателям прогнозируемую урожайность привойно-подвойных комбинаций и определить корреляцию между фактическими и прогнозируемыми оценками. По экспе- риментально полученным данным (см. табл. 1) мы вычислили значения gi и gk для каждого привоя и подвоя (табл. 2).
Для прогнозов оценок урожайности привитых деревьев по ОКС прививочных компонентов и обусловленных СКС отклонений от таких оценок используются разные формулы. Вклады эффектов g i и g k в прогнозные по ОКС оценки урожайности х ik вычисляются с помощью уравнения:
x ik = х.. + g i + g k , [5] которое можно представить как
X k = х i . + g k , [6] так как из уравнения [3] следует, что х i . = g i + х...
Формула [6] свидетельствует, что обусловленную общей комбинационной способностью i -го привоя и к -го подвоя урожайность х ik привойно-подвойной комбинации (без учета эффектов СКС) можно представить как среднюю на разных подвоях урожайность х i . i -го сорта с поправкой g k на эффект ОКС k к -го подвоя. Очевидно, что вычисления прогнозируемых оценок урожайности х ik (см. табл. 2) с использованием формул [5] и [6] дают одинаковые результаты.
Сопоставление значений прогнозируемой урожайности х ik (см. табл. 2) с полученными экспериментально оценками x ik (см. табл. 1) позволяет установить, с какой степенью надежности удается прогнозировать урожайность привойно-подвойных комбинаций яблони на основании учета эффектов общей комбинационной способности привоев и подвоев. Вычисленный по данным этих таблиц коэффициент корреляции между значениями x ik и х ik оказался высоким: r = 0,930 (переходит 0,1 % уровень существенности), чему соответствовал и коэффициент детерминации для сопряженной изменчивости ( Cd = 86,5 %). Это свидетельствует о достаточно высокой эффективности таких прогнозов, что позволяет включать в опыты по испытанию привитых деревьев только наиболее перспективные сочетания привоев и подвоев, не анализируя комбинации с достаточно надежно низкой прогнозной оценкой урожайности. Характеризуя значимость методов оценки комбинационной способности генотипов для селекции, П.Ф. Рокицкий (24) писал, что они были разработаны вначале применительно к кукурузе, но вполне приложимы и к другим объектам, поскольку дают значительно большую информацию о результатах скрещиваний, чем чисто эмпирический анализ скрещиваний различных рас, линий или пород. Для прогнозной оценки урожайности привойно-подвойных комбинаций плодовых культур использование аналогичного метода важнее настолько, насколько испытание привитых деревьев требует больше времени и средств, чем изучение гибридов F 1 однолетних культур.
Сравнение данных, представленных в таблицах 1 и 2, свидетельствует о том, что ни одна из вычисленных величин х ik точно не совпала с полученными при испытаниях значениями x ik , хотя 7 из 28 прогнозных оценок отклонились от фактических менее чем на 2 ц/га (сорт Джонатан на подвоях 1-48-46, М2, М3 и М7, сорт Голден Делишес — на М2 и М3, а также сорт Корей — на 1-48-46). Причина несовпадений х к и x ik становится понятной при сравнении формул [2] и [5]: при прогнозах только по общей комбинационной способности привоев и подвоев (см. табл. 2) не учитываются эффекты взаимодействия между прививочными компонентами, то есть специфическая комбинационная способность Sik , эффекты которой вычисляются по формуле (22):
S ik x ik + x.. — x i . — x. k ; [7] данные для этих вычислений взяты из таблицы 1, а результаты представлены в таблице 3.
3. Эффекты специфической комбинационной способности ( S ik , ц/га) для сочетаний привоев и подвоев яблони (ОПХ «Центральное» СКЗНИИСиВ, г. Краснодар, 1982-2002 годы)
|
Привой, сорт |
Подвой, форма |
||||||
|
1-48-1 |
1-47-55 |
1-48-46 | |
М2 | |
М3 |
| М4 |
| М7 |
|
|
Джонатан |
- 9,1 |
- 9,6 |
- 0,8 |
1,8 |
0,5 |
16,2 |
1,2 |
|
Голден Делишес |
4,1 |
25,7 |
- 14,4 |
1,6 |
- 1,2 |
- 12,6 |
- 3,0 |
|
Айдаред |
13,0 |
9,4 |
13,8 |
- 8,8 |
- 15,8 |
- 6,6 |
- 5,2 |
|
Корей |
- 8,1 |
- 25,4 |
1,5 |
5,5 |
16,6 |
3,0 |
7,1 |
Как и при оценках проявлений СКС в парах родителей гибридов F 1 , особенность эффектов специфической комбинационной способности отдельных сочетаний привоев и подвоев заключается в том, что суммы этих эффектов равны нулю как для каждого привоя, так и по столбцам таблицы (отклонения за счет округлений не превышают 0±1) (см. табл. 3):
( Е sik = о).
k
При сопоставлении величин СКС (см. табл. 3) было обнаружено, что в среднем эффекты взаимодействия сорта Джонатан с изученными подвоями минимальны, у подвоя 1-47-55 взаимодействие с сортами наиболее значительное, у М2 и М7 — минимальное. Выявить более четкие закономерности варьирования для величин Sik по всей совокупности данных, представленных в таблице 3, не удалось. Наблюдалась только тенденция к увеличению значений Sik с ростом x ik у сортов Джонатан и Корей.
В целом для всей совокупности данных (см. табл. 1 и 3) корреляция между величинами x ik и Sik оказалась хотя и существенной на 5 % уровне значимости, но низкой — r = 0,368 при Cd = 13,5 %. Следовательно, прогнозы урожайности привитых деревьев на основании оценки СКС прививочных компонентов, то есть по эффектам взаимодействия «привой—подвой», ненадежны. В связи с этим следует отметить, что в биометрической генетике под термином «взаимодействие» понимается отклонение от аддитивности компонентов вариации. В обсуждаемом случае представленные в таблице 2 прогнозные величины х k урожайности комбинаций яблони вычислялись по аддитивным вкладам эффектов привоев и подвоев, а их СКС можно оценивать не только по формуле [7], но и по следующей формуле:
S ik = x ik - х ik , [8] поскольку взаимодействие здесь понимается как проявление неаддитивности. Выше было показано, что предсказуемая по эффектам ОКС привоев и подвоев вариация урожайности у привитых деревьев характеризовалась Cd = 86,5 %, а непредсказуемые вклады СКС — Cd = 13,5 %, то есть для изученной совокупности прививочных компонентов предсказуемая часть вариации признака у привойно-подвойных комбинаций оказалась в 6,4 раза больше непредсказуемой.
Полученные результаты свидетельствуют о том, что дальнейшее совершенствование прогноза урожайности у привойно-подвойных комбинаций требует углубленного анализа той части вариации этого признака у привитых деревьев, которая обусловлена непредсказуемыми эффектами СКС их прививочных компонентов. При таком анализе следует использовать как биологический, так и статистический подходы. Биологический подход должен быть основан на изучении морфофизиологических особенностей привоев, подвоев и привитых деревьев, характеризующихся разной степенью проявления СКС. При статистическом подходе следует разделять дисперсию взаимодействия привоев и подвоев на два компонента — прогнозируемый линейный и непредсказуемый нелинейный.
Некоторые причины завышенных оценок S ik для отдельных сочетаний i -х привоев с к -ми подвоями выявлялись уже при анализе данных, представленных в таблицах 1 и 2. Вследствие того, что при вычислениях X ik используется одна и та же усредненная величина g i для i -го сорта, различия по значениям х ik Между парами сортов были одинаковыми на всех подвоях, то есть не учитывались особенности реакции i -го привоя на конкретный подвой. Например, по средней урожайности x i . и по g i сорта Голден Делишес и Айдаред различались на 0,1 ц/га, поэтому и все значения их х ik различались на 0,1 ц/га. Одинаковыми и равными 59,6 ц/га оказались различия как средних величин x i ., так и прогнозных оценок х ik у сортов Корей и Джонатан. По той же причине (из-за описанного способа вычислений) к -й подвой вносил равные вклады g k в прогнозируемые по ОКС величины х ik , поЭтому, например, значения х ik для Любого из четырех сортов на подвоях 1-48-1 и 1-47-55 различались на одинаковую величину — 6,1 ц/га. Вследствие такого прогнозирования х ik по ОКС прививочных компонентов у низкоурожайных привойно-подвойных комбинаций оценки оказываются завышенными, у высокоурожайных — заниженными, что приводит к завышению оценок специфической комбинационной способности привоев и подвоев и снижению эффективности прогнозов урожайности у привитых деревьев. Насколько велики такие искажения, можно судить по параметрам уравнений регрессии для графиков зависимости х ik от x ik , построенных по данным таблиц 1 и 2 (рис.).
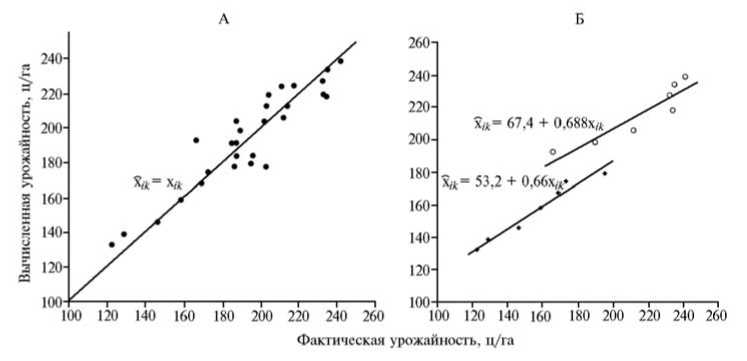
Соотношение вычисленной по общей комбинационной способности привоев и подвоев (х к , ц/га) и фактической (x ik , ц/га) урожайности для привитых деревьев у сортов яблони Джонатан, Голден Делишес, Айдаред и Корей ( • ) (А) , а также у сортов яблони Джонатан ( ♦ ) и Корей ( о ) (Б) (ОПХ «Центральное» СКЗНИИСиВ, г. Краснодар, 1982-2002 годы).
В соответствии с формулами [3], [4] и [5], по которым вычислялась прогнозируемая по ОКС привоев и подвоев урожайность х ik , коэффициент регрессии (b) величин 'kk на величины x ik должен быть равен 1, а линия — проходить через начало координат. Это означает, что если фактическая урожайность привойно-подвойной комбинации выше средней x.. на 1 ц/га, то прогнозная оценка ее урожайности также в среднем возрастает на 1 ц/га. Поэтому линия регрессии на таком графике должна описываться формулой х к = x ik . Линия на приведенном графике (см. рис., А) вполне соответствует распределению точек на нем. При этом сдвиги точек вдоль линии обусловлены вычисленными оценками ОКС прививочных компонентов, а степень удаленности точек служит мерой как проявления СКС, так и несовпадения прогнозных и фактических оценок.
На графике слева (см. рис., А) точки для всех 28 комбинаций четырех сортов на семи подвоях были представлены общей совокупностью. Иная закономерность выявлялась, если в той же системе координат рассматривали характер распределения тех же точек, но отдельно для каждого сорта на разных подвоях. На таком графике (см. рис., Б) на примере сортов Джонатан и Корей обнаружилось, что в уравнениях регрессий х к на x ik появлялись свободные члены, а коэффициенты регрессий снижались настолько, что с увеличением фактической урожайности на 1 ц/га ее прогнозируемая по ОКС величина возрастала приблизительно на 2/3 ц/га, поскольку в среднем b = 0,674. Это обусловлено уменьшением различий между прогнозными оценками урожайности при вычислениях по формулам [3], [4] и [5]. Подобные систематические ошибки вычислений служат одной из причин отклонений точек от линии х ik = x ik (см. рис. А), завышения показателя СКС и соответствующего снижения эффективности прогнозов для сортоподвойных комбинаций по ОКС привоев и подвоев. Из этого следует, что разработка математической модели, свободной от таких ошибок, позволит еще больше повысить точность прогнозов урожайности привитых деревьев. На основании дополнительного повышения точности подобных прогнозов может быть создан метод и предложены формулы для разделения эффектов СКС на два компонента — прогнозируемый линейный и непрогнозируемый нелинейный.
В существующих руководствах по оценке ОКС и СКС у родителей гибридов F 1 (22-24) не учитываются систематические ошибки вычислений СКС и прогнозов урожайности гибридов по ОКС родительских форм, выявленные нами, и даже не ставится задача расчленения эффектов СКС на линейный и нелинейный компоненты. Сама оценка комбинационной способности родителей по учетам урожайности гибридов эмпирична, основана на многочисленных скрещиваниях и испытаниях (25-27). При этом Л.В. Хоты-лева и Л.А Тарутина (26) констатируют, что все эти сложности порождаются недостаточной изученностью генетики комбинационной способности. По той же причине трудно судить, насколько целостный организм привитого дерева функционально аналогичен гибриду F 1 от скрещивания двух родителей. Однако довольно высокая точность прогнозов урожайности комбинаций по ОКС привоев и подвоев свидетельствует о значительной степени интегрированности свойств прививочных компонентов в признаках привитого дерева.
Таким образом, генотипы привоев различаются как по средней на разных подвоях урожайности, так и по степени зависимости этого признака от особенностей конкретных подвоев, а подвои в неодинаковой мере дифференцируют привои по фенотипическому проявлению урожайности. Эта изменчивость комбинационной способности у прививочных компонентов обусловила вариацию продуктивности привойно-подвойных комбинаций яблони в проведенном 21-летнем испытании в диапазоне от 122,7 до 241,6 ц/га. Установлено, что методы и математические модели, широко используемые для оценки комбинационной способности родительских линий у гибридов F1, можно с успехом применять для определения общей и специфической комбинационной способности (ОКС и СКС) у привоев и подвоев и влияния их генотипов на урожайность привитых деревьев с целью прогнозирования лучших привойно-подвойных комбинаций. Высокая эффективность прогнозов урожайности таких комбинаций у яблони обусловлена тем, что в структуре вариации комбинационной способности прививочных компонентов преобладает доля ОКС, которая (по аналогии с родительскими формами гибридов) представляет собой функцию аддитивных генов. Тесная корреляция между прогнозируемой по ОКС и фактиче- ской урожайностью привойно-подвойных комбинаций свидетельствует о высокой интегрированности свойств прививочных компонентов в определение количественных признаков у привитого дерева. СКС прививочных компонентов, зависящая от неаддитивных эффектов их генотипов, не может использоваться для прогнозов урожайности привитых деревьев. Доля вклада этой части вариации комбинационной способности в изменчивость фактической урожайности в парах «привой—подвой» оказалась в 6,4 раза меньше таковой для предсказуемой аддитивной части. На основании предварительного анализа доли вариаций, обусловленной эффектами СКС прививочных компонентов, выявлена возможность повышения надежности прогнозов урожайности привойно-подвойных комбинаций посредством вычленения из этих эффектов линейного компонента дисперсии.
