Содержание гликоалкалоидов и состав белков в листьях картофеля при повреждении колорадским жуком

Автор: Цветкова М.А., Цветков И.Л., Коничев А.С.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Иммунитет и защита растений
Статья в выпуске: 1 т.39, 2004 года.
Бесплатный доступ
Определяли содержание гликоалкалоидов и качественный состав водорастворимых белков в листьях различных видов, подвидов и сортов картофеля (род Solanum) при повреждении надземной части растений колорадским жуком. Рассматривается возможность корреляционной связи между накоплением гликоалкалоидов в поврежденных листьях картофеля и функцией новообразованного белка с молекулярной массой 93 кД. Предлагается использовать в качестве критерия оценки устойчивости растений картофеля к листогрызущим насекомым содержание гликоалкалоидов в листьях и адреналина у питавшихся ими особей колорадского жука.
Короткий адрес: https://sciup.org/142132956
IDR: 142132956 | УДК: 635.21:631.524.86:632.7
Glycoalkaloids content and protein composition in potato leaves after its damage by potato beetle
The authors determined the glycoalkaloids content and qualitative composition of water soluble protein in leaves of different species, subspecies and varieties of potato (Solanum genus) after damage of overground part of plants by potato beetles. It was shown, that glycoalkaloids content in potato leaves is associated with plant's protectability from leaves gnawing insects and after damages by potato beetles the glycoalkaloids content increases in a majority of investigated plants. The correlation between glycoalkaloids accumulation in damaged potato leaves and a function of neogenic protein with mass of 93 kD was considered. The authors propose to use as criterion for estimation of tolerance of potato plants to leaves gnawing insects the glycoalkaloids content in leaves and adrenalin content in feeding by them potato beetles.
Текст научной статьи Содержание гликоалкалоидов и состав белков в листьях картофеля при повреждении колорадским жуком
Основным защитным механизмом при формировании устойчивости растений картофеля к поражению клубней моллюсками, личинками насекомых и паразитическими червями является образование эндогенных аллелохимических соединений — вторичных продуктов метаболизма (терпеноиды, флавоноиды, фенолы, гликоалкалоиды, небелковые аминокислоты, хиноны, гликозиды, гликозинолаты и др.), оказывающих токсическое действие и тормозящих рост и развитие вредителей (1, 2).
Наиболее ощутимый вред посадкам картофеля наносят насекомые-листоеды и прежде всего колорадский жук (что особенно актуально для Европы и России). Факторы, обусловливающие устойчивость растений к листогрызущим насекомым, многообразны: механическая защита листовой пластинки (опушенность и др.), им-мунохимические реакции, накопление в тканях листа разнообразных защитных белков (дефензины, ингибиторы протеиназ, растительные токсины) и фитотоксинов небелковой природы (аллелохимические вещества), существенно снижающих питательную ценность зеленой массы для насекомых-листоедов (3).
Токсические свойства, которыми обладают гликоалкалоиды в отношении насекомых, признаются сейчас очень многими авторами. В специальных исследованиях установлено, что гликоалкалоиды картофеля способны ингибировать ацетилхолинэ-стеразу, что существенно затрудняет проведение нервного импульса и блокирует координирующую деятельность центральной нервной системы, в том числе и у насекомых (4). На суспензионных культурах эритроцитов кролика, клеток корня свеклы и протопластов Penicillium notatum Westling показано, что гликоалкалоиды существенно изменяют проницаемость клеточных мембран, а в определенной концентрации вызывают лизис клеток (5). Следует отметить, что для колорадского жука и насекомых вообще поступление с пищей некоторых чужеродных стероидов просто необходимо для создания собственных клеточных элементов из-за неспособности синтезировать стериды и, в частности, холестерин самостоятельно (6). Однако включение в этот процесс стероидных гликоалкалоидов может иметь нежелательные и даже фатальные последствия для метаболизма насекомых.
В задачу нашей работы входило определение содержания и качественного состава защитных белков и небелковых соединений, в частности гликоалкалоидов, в листьях растений разных видов, сортов картофеля и дикорастущих сородичей при повреждении колорадским жуком.
Методика . Объектом исследования служили следующие виды картофеля: Solanum commersonii Dun., S. demissum Lindl., S. spegazzini Bitt., S. polyadenium Greenm., S. andigenum Juz. et Buk. (ssp. argentinicum и ssp. colombianum ), S. goniocalyx
Juz. et Buk. и S. tuberosum L. (сорта Вятка, Елизавета, Жуковский, Зарево, Луговской, Невский, Петербургский, Темп).* В качестве грунта использовали верхний слой тщательно просеянной почвы хвойного леса, которую помещали в деревянные ящики; плотность посадки составляла одно растение на 1-2 дм2. Образцы картофеля культивировали при ежедневном искусственном поливе.
Имаго колорадского жука первого «летнего» поколения, проходящего нажиров-ку, собирали в полевых условиях на посадках картофеля (Смоленская обл., Гагаринский р-н). Для того чтобы избежать нежелательного проявления полиморфизма природных популяций колорадского жука и соответствующих им особенностей пищевой специализации, мы старались отбирать жуков, полностью идентичных по рисунку переднеспинки (7).
Особей колорадского жука с одинаковым «фенообликом» подсаживали по 10 шт. на листья четырех-пяти верхних ярусов отдельных растений картофеля. Во избежание свободного перемещения жуков их помещали в садки из тонкой капроновой сетки, укрепленные у основания листьев. Удаляли жуков с растения после того, как они уничтожали приблизительно 1/3 часть всей площади листовой пластинки (обычно через 16-20 ч). В тушках жуков определяли содержание адреналина по методу Ронина (8), модифицированному для работы с личинками и имаго насекомых (9); в срезанных частях листьев (сразу же после удаления жуков, а также через 1, 3 и 5 сут) — содержание гликоалкалоидов по методу Тукало (10) и качественный состав белков методом ДДС-Na-электрофореза (0,1 % додецилсульфат натрия) в полиакриламидном геле по Лэммли (11) с окрашиванием белковых фракций кумасси R-250.
Для оценки специфичности повреждений сравнивали результаты биохимического анализа листьев индивидуальных растений, поврежденных жуками, имеющими разные фенотипы (I опыт), и листьев, механически поврежденных (II опыт) посредством постепенного надрезания края листа ножницами на протяжении I опыта. Контролем служили, во-первых, неповрежденные листья индивидуальных растений, участвовавших в I и II опытах (контроль 1), во-вторых, — листья с неповрежденного растения (контроль 2). Для чистоты эксперимента на всех листьях в контроле и II опыте укрепляли капроновые садки без жуков.
Статистическую обработку данных проводили по t -критерию Стьюдента.
Результаты . Наиболее высоким содержанием гликоалкалоидов (600 мг/100 г сырой массы и более) характеризовались дикорастущие виды картофеля S. polyadenium , S. demissum и S. goniocalyx (табл. 1). У большинства сортов S. tuberosum (за исключением Зарево и Вятка) содержание гликоалкалоидов было почти в 4 раза ниже — не более 150 мг/100 г сырой массы. Содержание гликоалкалоидов в листьях растений подвидов S. andigenum , а также дикорастущих видов S. commersonii и S. spegazzini составляло в среднем около 450 мг/100 г сырой массы.
1. Содержание гликоалкалоидов в листьях интактных и пораженных колорадским жуком растений картофеля разных видов, подвидов и сортов в зависимости от продолжительности повреждающего воздействия (мг/100 г сырой массы)
|
Вид, подвид, сорт |
I опыт (листья, поврежденные имаго Контроль 1 (интактные листья с поврежден-ных II опыт (механически по- Контроль 2 (интактные колорадского жука) колорадским жуком растений) врежденные листья) растения) В р е м я в з я т и я п р о б ы, сут 1-е 3-и 5-е 3-и 1 5-е 1 5-е |
|
Solanum andigenum: ssp. argentinicum ssp. colombianum S. commersonii S. demissum S. goniocalyx S. polyadenium S. spegazzini S. tuberosum , сорт: Вятка Елизавета Жуковский Зарево Луговской Невский Петербургский Темп |
483 ± 21 503 ± 17 516 ± 14 476 ± 15 521 ± 18 – 477 ± 18 496 ± 15 516 ± 13 519 ± 17 – – 471 ± 16 475 ± 16 511 ± 17 506 ± 18 – 523 ± 17 511 ± 15 486 ± 16 505 ± 17 597 ± 14 618 ± 19 639 ± 20 – 630 ± 17 576 ± 18 612 ± 20 – – – – – 578 ± 18 600 ± 19 625 ± 19 631 ± 19 652 ± 20 – 641 ± 21 654 ± 24 638 ± 22 – 512 ± 18 – 489 ± 16 – – 515 ± 16 225 ± 7,9 231 ± 6,9 216 ± 8,0 – 208 ± 6,7 198 ± 5,9 212 ± 7,4 128 ± 5,2 120 ± 3,6 127 ± 4,0 – 117 ± 3,8 121 ± 5,3 121 ± 4,0 – 153 ± 5,3 144 ± 4,7 – 135 ± 4,3 140 ± 4,1 147 ± 5,3 308 ± 9,2 347 ± 11 364 ± 10 324 ± 10 366 ± 11 296 ± 9,8 311 ± 9,3 130 ± 7,5 137 ± 4,3 148 ± 4,4 134 ± 4,0 145 ± 4,4 121 ± 3,9 124 ± 4,0 113 ± 4,0 110 ± 2,8 120 ± 5,4 – 124 ± 3,4 115 ± 3,6 112 ± 3,9 140 ± 4,5 153 ± 4,2 162 ± 4,8 133 ± 4,2 158 ± 4,6 140 ± 4,2 138 ± 4,1 128 ± 4,5 135 ± 4,0 142 ± 5,3 130 ± 4,0 137 ± 3,1 108 ± 5,6 121 ± 4,6 |
П р и м е ч а н и е. Прочерк означает отсутствие данных. Описание вариантов опыта см. в разделе «Методика».
2. Содержание адреналина у имаго колорадского жука, питавшегося на листьях разных видов, подвидов и сортов картофеля (мг%)
|
Вид, подвид, сорт |
Содержание адреналина |
|
Solanum andigenum: |
|
|
ssp. argentinicum |
7,54 ± 0,24 |
|
ssp. colombianum |
7,48 ± 0,23 |
|
S. commersonii |
11,60 ± 0,40 |
|
S. demissum |
14,90 ± 0,50 |
|
S. goniocalyx |
13,80 ± 0,40 |
|
S. polyadenium |
15,80 ± 0,50 |
|
S. spegazzini |
7,21 ± 0,23 |
|
S. tuberosum, сорт: |
|
|
Вятка |
4,18 ± 0,11 |
|
Елизавета |
3,16 ± 0,09 |
|
Жуковский |
3,28 ± 0,10 |
|
Зарево |
4,84 ± 0,16 |
|
Луговской |
3,15 ± 0,10 |
|
Невский |
3,12 ± 0,08 |
|
Петербургский |
3,24 ± 0,03 |
|
Темп |
3,41 ± 0,10 |
Известно, что у насекомых адреналин выделяется в гемолимфу из corpus cardiacum в состоянии стресса (например отравление ДДТ), оказывая определенный физиологический эффект на нервную систему, чем способствует смене гиперактивности на состояние прострации. В то же время адреналин насекомых участвует в регуляции углеводного обмена, контролируя распад гликогена в мышцах, а также концентрацию свободной трегалозы в гемолимфе (6).
Основываясь на этих данных, следует полагать, что наименее пригодными для питания колорадского жука, а значит, и наиболее устойчивыми к нему оказались представители видов S. polya-denium, S. demissum и S. goniocalyx, а наименее устойчивыми — все исследованные сорта S. tuberosum, за исключением сорта Зарево (табл. 2). Питание листьями растений сорта Зарево вызывало у колорадских жуков увеличение содержания адреналина (55 %) в значительно большей степени, чем у тако- вых, заселявших другие сорта картофеля. В листьях растений этого сорта выявлено также высокое содержание гликоалкалоидов (311 мг/100 г сырой массы). Доказательством
взаимосвязи между содержанием в листьях гликоалкалоидов и интенсивностью выделения адреналина у колорадских жуков может служить высокая устойчивость сорта Зарево к последним.
Тесная положительная корреляция между содержанием гликоалкалоидов в листьях растений картофеля и содержанием адреналина у имаго колорадского жука ( r = 0,92) выявлена и для других образцов картофеля, включая дикорастущие виды и примитивный культурный вид S. andigenum (рис. 1).
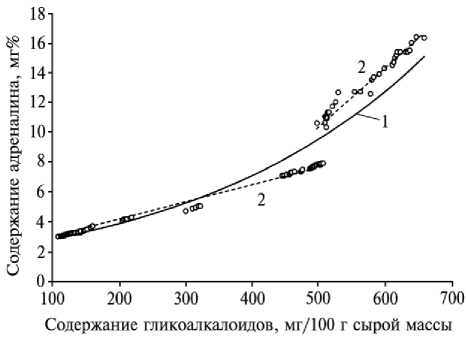
Рис. 1. График корреляционной зависимости между содержанием адреналина у имаго колорадского жука и количеством гликоалкалоидов в листьях картофеля: 1 — экспонента (линия тренда), 2 — отрезки экспоненциальной зависимости, близкие к прямолинейным.
Следовательно, гликоалкалоиды листьев картофеля обеспечивают защиту растений от повреждения колорадским жуком. Поскольку при увеличении содержания гликоалкалоидов в листьях количество адреналина у колорадского жука возрастает, этот признак можно использовать в качестве критерия оценки устойчивости растений к листогрызущим насекомым.
Исследование динамики содержания гликоалкалои- дов при повреждении листьев некоторых образцов культурного картофеля позволило нам получить еще одно экспериментальное подтверждение непре- менного участия этих соединений в формировании устойчи вости растений к насекомым-листоедам. У ряда исследованных сортов (Зарево, Лу-говской, Петербургский, Темп) отмечено прогрессирующее увеличение содержания гликоалкалоидов, наблюдаемое иногда уже через 24 ч после подсаживания жука, а через 5 сут оно достигало 17-18 % от первоначального уровня (рис. 2).
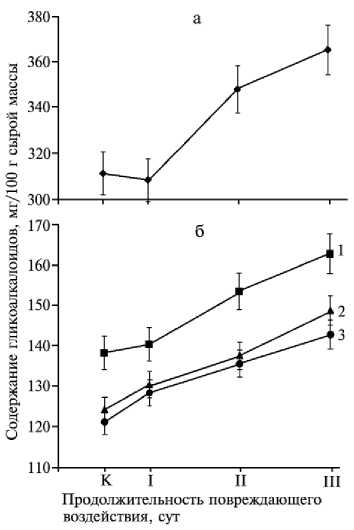
Рис. 2. Содержание гликоалкалоидов в листьях различных сортов картофеля ( Solanum tuberosum ) после повреждающего воздействия колорадского жука в течение 1 (I), 3 (II) и 5 (III) сут: а — сорт Зарево; б — сорта Петербургский (1), Луговской (2) и Темп (3); К — контроль (содержание гликоалкалоидов в интактном растении).
Следует отметить, что накопление гликоалкалоидов в поврежденных колорадским жуком листьях картофеля можно было зарегистрировать, используя в качестве контроля (норма) только полностью интактные растения (контроль 2). В неповрежденных листьях растений, заселенных жуком (контроль 1), содержание гликоалкалоидов так же, как и в I опыте, не оставалось постоянным, а возрастало по ходу эксперимента практически синхронно (см. табл. 1). Вместе с тем механическое повреждение, имитировавшее погрызы жука (II опыт), не оказывало влияния на содержание гликоалкалоидов.
Итак, на примере сортов картофеля Зарево, Луговской, Петербургский и Темп показано, что гликоалкалоиды могут служить не только конституционным, но и факультативным (динамическим) защитным фактором, который активируется в состоянии стресса растения и к тому же специфичен в отношении характера нанесенного повреждения. По всей видимости, индуктором образования гликоалкалоидов в клетках листьев является не просто их механическое повреждение, которое само по себе встречается весьма редко, а какое-то биологически активное вещество, выделяемое колорадским жуком при питании и, вероят нее всего, являющееся обычным компонентом слюны насекомого. Нам пока не известно, каким именно образом слюна колорадского жука может индуцировать выде- ление стероидных гликоалкалоидов при повреждении листьев, однако механизм регуляции содержания этих метаболитов в растении можно представить вполне определенно. В частности, известно, что биосинтез стеролов и изопреноидов в основном лимитируется действием ключевого метаболита — мевалоновой кислоты — и находится под контролем 3-гидрокси-3-метилглутарил-КоА-редуктазы (ГМГР, КФ 1.1.1.34). Сравнительно недавно было показано, что регуляция активности этого фермента происходит на уровне транскрипции, так как мРНК образуется только в ответ на повреждение клеток, стресс (например закисление межклеточного пространства или фарнезол-индуцированная гибель клеток в суспензионной культуре Nicotiana tabacum L.) или здесь действуют какие-либо «сигнальные» факторы, осуществляющие внешнюю регуляцию метаболизма (например свет при синтезе пигмента шиконина у Lithospermum erythrorhizon Sieb. et Zucc.) (12, 13). У картофеля ГМГР кодируется целым семейством генов, индукция экспрессии которых происходит строго дифференцированно в зависимости от того, каким фактором она была вызвана — фитопатогеном, повреждением тканей листа или обработкой элиситором (арахидоновая кислота) (14).
Вполне возможно, что и в нашем случае какие-то компоненты, выделяемые слюнными железами насекомого, индуцировали специфическую экспрессию одного из множества изоферментов ГМГР, который катализировал образование мевалоновой кислоты, служащей источником углерода при биосинтезе стероидной группы гликоалкалоидов, что способствовало повышению содержания последних в клетке. При этом надо полагать, что у растений в этой цепочке должен присутствовать как минимум еще один «фактор», осуществляющий информационную связь между поврежденными и интактными тканями. Распространение этого «фактора» должно осуществляться посредством как ближнего (внутри листа), так и дальнего транспорта (между отдельными листьями), так как только это могло быть причиной почти синхронного увеличения содержания гликоалкалоидов как в поврежденных, так и в интактных листьях, расположенных на том же поврежденном растении (контроль 1).
При анализе качественного состава и электрофоретических спектров белков из листьев интактных и поврежденных растений картофеля нами установлено, что у всех образцов, характеризовавшихся увеличением содержания гликоалкалоидов в ответ на повреждение, уже через 1 сут после начала эксперимента появлялась новая фракция белков, молекулярная масса которых составляла около 93 кД (рис. 3). Такое сходство молекулярной массы вновь образованного белка у всех исследованых образцов картофеля, в том числе и дикорастущих видов, позволяет сделать предположение о структурной гомологии этого белка и, возможно, общем характере механизма формирования устойчивости, по крайней мере, для представителей рода Solanum .
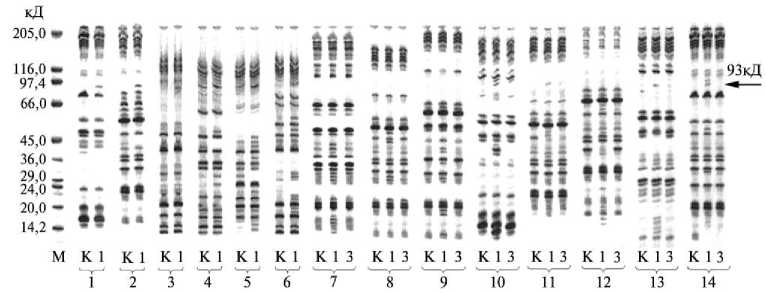
Рис. 3. Электрофореграмма денатурированных белков из листьев растений различных видов и сортов картофеля при повреждении колорадским жуком (10 % ПААГ, 0,1 % додецилсульфата натрия) : 1 — Sola-num andigenum ssp. argentinicun ; 2 — S. andigenum ssp. colombianum ; 3 — S. commersonii ; 4 — S. demissum ; 5 — S. goniocalyx ; 6 — S. spegazzini ; 7, 8, 9, 10, 11, 12, 13 и 14 — S. tuberosum соответственно сорта Вятка, Елизавета, Жуковский, Зарево, Луговской, Невский, Петербургский и Темп; М — маркер молекулярной массы, К — контроль (интактные растения), 1 и 3 — повреждающее воздействие в течение соответственно 1 и 3 сут.
Сопоставив данные об изменении содержания гликоалкалоидов и качественного состава белков, можно предположить, что белком, синтезированным de novo в листьях поврежденных растений, как раз и является ГМГР, лимитирующая синтез гликоалкалоидов на стадии образования мевалоновой кислоты. Еще одним косвенным подтверждением этого предположения является сообщение Choi с соавт. о дифференциальной активации двух картированных ГМГР-генов ( hmg -1 и hmg -2) S. tuberosum в зависимости от внешних условий, причем повреждение растения индуцировало образование только hmg -1-транскриптов (15). Длина hmg -1-тран-скрипта, по данным тех же авторов, составляет приблизительно 2,5 тыс. пар нуклеотидов. Нетрудно подсчитать, что молекулярная масса белкового продукта, синтезируемого на мРНК такой длины, должна составлять как раз около 90 кД.
Далее у всех без исключения экспериментальных образцов картофеля отмечено образование одного-двух, а чаще сразу нескольких низкомолекулярных белков — от 15 до 24 кД. В целом белки этой группы отличались от белка с молекулярной массой 93 кД значительно большим разнообразием, но вместе с тем у них наблюдается определенная видоспецифичность. У отдельных сортов S. tuberosum белки с молекулярной массой 15-24 кД были более или менее однородными (см. рис. 3). Вполне возможно, что эти белки являются оксипролин- и пролинбогатыми гликопротеинами (ПБГП), физиологическая роль которых до конца еще не выяснена. Предполагают, что ПБГП представляют собой, с одной стороны, специфические агглютинины, синтезируемые в ответ на чужеродный белок, а с другой — являются компонентами кле- точных мембран, принимающих участие в регуляции проницаемости и хеморецепции клеток (16).
Таким образом, повреждение растений картофеля (род Solanum ) колорадским жуком индуцирует синтез гликоалкалоидов и вызывает прогрессирующее увеличение их концентрации во всех листьях; механическое повреждение листьев не отражается на содержании гликоалкалоидов. Показано, что содержание адреналина у колорадского жука тесно коррелирует с содержанием гликоалкалоидов в поврежденных листьях. У всех исследованных образцов картофеля, которые характеризовались высоким содержанием гликоалкалоидов в листьях при повреждении растений колорадским жуком, происходит образование de novo высокомолекулярного белка (93 кД), а также целого ряда низкомолекулярных белков (15-24 кД). По нашему мнению, выделение гликоалкалоидов в листьях и кожуре клубней поврежденных колорадским жуком растений картофеля служит одним из факторов системной индуцированной устойчивости последних к листогрызущим насекомым, специфическим в отношении характера наносимого повреждения. Содержание гликоалкалоидов в листьях картофеля, а также содержание адреналина у имаго колорадского жука, заселяющего последние, могут служить критерием оценки устойчивости образцов различных видов и сортов картофеля к листогрызущим насекомым.
Л И Т Е Р А Т У Р А
-
1. К е ф е л и В.И. Природные ингибиторы роста. Физиол. раст., 1997, 44, 3: 471-480.
-
2. П а с е ш н и ч е н к о В.А. Регуляция терпеноидного биосинтеза в растениях и его связь с биосинте
зом фенольных соединений. Физиол. раст., 1995, 42, 5: 787-804.
-
3. В и л к о в а Н.А., Г у с е в а Н.Н. Проблемы иммунитета растений к вредителям и болезням. Вест. РАСХН, 1994, 6: 11-13.
-
4. B u s h w a y R.J., S a v a g e S.A., F e r g u s o n B.S. Inhibition of acetyl cholinesterase by solanaceous glycoalkaloids and alkaloids. Amer. Potato J., 1987, 64, 8: 409-413.
-
5. R o d d i c k J.G., R i j n e n b e r g A.L., O s m a n S.F. Synergistic interaction between potato glycoalkaloids α α -solanine and α α -chaconine in relation to destabilisation of cell membranes: ecological implication. J. Chem. Ecol., 1988, 14, 3: 889-902.
-
6. С в и д е р с к и й В.А. Основы нейрофизиологии насекомых. М., 1986.
-
7. Ф а с у л а т и С.Р. Анализ структуры популяций колорадского жука и его значение для разработки
зональных систем защиты картофеля. Бюл. ВИЗР. СПб, 1987, 63: 38-43.
-
8. Р о н и н Б.С., С т а р о б и н е ц Г.М. Руководство к практическим занятиям по методикам клинических лабораторных исследований. М., 1989.
-
9. Методические рекомендации по изучению и оценке форм картофеля на устойчивость к колорадскому жуку /Под ред. Н.А. Вилковой. М., 1993.
-
10. Т у к а л о Е.А., Ц а р и к Г.Н. Ускоренный метод количественного определения гликоалкалоидов картофеля. Науч. докл. высшей школы (сер. Биологические науки), 1970, 12: 115-117.
-
11. L a e m m l i U.K. Cleavage of structural proteins during the assembly of the head of the bacteriophage T4. Nature, 1970, 227, 5259: 680-685.
-
12. H a m m e r l i n A., B a c h T.J. Farnesol-induced cell death and stimulation of 3-hydroxy-3-methylglutaril-coenzyme A reductase activity in tobacco cv Bright Yellow-2 cells 1,2. Plant Physiol., 2000, 123, 8: 1257-1268.
-
13. L a n g e M.B., S e v e r i n K., B e c h t h o l d A. e.a. Regulatory role of microsomal 3-hydroxy-3-methylglutaryl-coenzyme A reductase for shikonin biosynthesis in Lithospermum erythrorhizon cell suspension cultures. Planta, 1998, 204, 2: 234-241.
-
14. Y a n g Z., P a r k H., L a c y G.H. e.a. Differential activation of potato 3-hydroxy-3-methylglutaryl coenzyme A reductase genes by wounding and pathogen challenge. Plant Cell, 1991, 3, 8: 397-405.
-
15. C h o i D., B o s t o c k R., A v d i u s h k o S. e.a. Lipid-derived signals that discriminate wound- and pathogen-responsive isoprenoid pathways in plants: Methyl jasmonate and the fungal elisitor arachidonic acid induce different 3-hydroxy-3-methylglutaryl-coenzyme A reductase genes and antimicrobial isoprenoids in Solanum tuberosum L. Proc. Natl. Acad. Sci. USA, 1994, 91, 3: 2329-2333.
-
16. Л о м о в а Л.А. Исследование экспрессии генов защитных белков в клубнях картофеля при ответе на стресс и после обработки индукторами устойчивости. Автореф. канд. дис. Новосибирск, 1995.
Московский педагогический государственный университет , 129278, Москва, ул. Кибальчича, 6
