Абиотические и биотические события на рубеже франского и фаменского веков

Бесплатный доступ
Короткий адрес: https://sciup.org/149128974
IDR: 149128974
Текст статьи Абиотические и биотические события на рубеже франского и фаменского веков
Рубеж франского и фаменского веков сопряжен с глобальными гeологи-ческими и, как следствие, биотическими событиями, одним из которых является мaссовое вымирание организмов морской биоты. Как отметил А. С. Алексеев [1], особенно резко изменился морской бентос. На этом уровне установлено полное исчезновение некоторых родов аммоноидей, целых отрядов среди брахиопод, трилобитов, почти полное исчезновение рифовых построек, сложенных скелетами кораллов и строматопорат..Изменения отмечены и в континентальной биоте. Однако анализ последовательности миоспоровых комплексов в наиболее стратиграфически полных пограничных франско-фаменских разрезах свидетельствует о том, что изменения в растительных сообществах не были столь драматичными, как в морской биоте.
Нижняя граница фаменского яруса принята в разрезе Upper Coumiac Quarry (Франция) в основании конодонтовой зоны Palmatolepis triangularis [12]. Граница проходит по кровле вeрхнего горизонта Кельвассер, совпадающей с франско-фаменским вымиранием, одним из пяти наиболее крупных в фане-розое (о масштабах данного биособытия имеются различные мнения [1]). В Восточной Eвропе в самых стратиграфически полных разрезах (в Уметовско-Линевской, Припятской и Днепровско-Донецкой депрессиях, на Южном Тимане) граница франского и фаменского ярусов по миоспорам установлена в основании зоны Corbulispora vimi-neus — Geminospora vasjamica, соответствующей волгоградскому горизонту. Там, где на этом уровне наблюдается перерыв в осадконакоплении (центральные районы Русской плиты), граница проводится в кровле ливенского и в подошве задонского горизонтов [6].
АБИОТИЧЕСКИЕ СОБЫТИЯ
Для выявления возможных причин вымирания организмов на франско-фа-менском рубеже необходимо рассмотреть весь комплекс глобальных событий 2
этого врeмeни. Maссовое вымирание на обсуждаемом рубеже коррелируется с глобальной регрессией и бескислородным событием. Анализ эвстатических колебаний уровня Mирового океана, позднедевонского рифтогенеза и супер-плюмовых событий демонстрирует дестабилизацию условий в атмосфере и гидросфере и, как следствие, изменения экологической обстановки и известные биособытия [5].
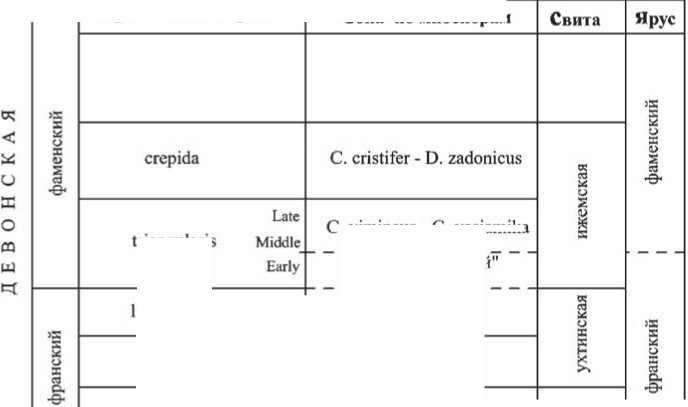
В настоящее время накопился значительный материал по характеристике колебаний уровня моря в это время в разных регионах Земли, приуроченных к различным тектоническим структурам. Изучение глубоководных разрезов Рейнских сланцевых гор в Западной Гeр-мании показало, что на самой франско-фаменской границе фиксируется начало регрессии, отмеченное хардграундом, в то время как верхний и нижний кель-вассерские горизонты соответствуют наиболее глубоководным фациям. Установлена определенная последовательность событий в разрезах Западной Европы и юго-запада США (рис. 1): коно-донтовая зона linguiformis — эвстатичес-кий подъем, приведший к аноксии, затем значительное эвстатическое опуска-ниe уровня моря — регрессия и массо-
Тимано-Печорская провинция
Зона по миоспорам
triangularis linguiformis
G. subsuta rhcnana
C. impcrpctuus
Рис. 1. Схема биостратиграфического расчленения пограничных франско-фаменских отложений подволгоградскии палинокомплекс
С. vimineus - G. vasjamika
вое вымирание; зона Early triangularis — продолжение регрессии; зона Middle triangularis — новая трансгрессия [19]. Восточнее, на территории Польско-Mо-равского шельфового бассейна, начало регрессии маркируется эрозионными несогласиями, хардграундами и брекчиями или поверхностями ненакопления. Начало последующей фаменской трансгрессии колеблется здесь от зоны Middle triangularis до зоны rhomboidea [17].
В центральной части Русской плиты почти повсеместно отсутствуют отложения зоны triangularis. В ее периферийных районах величина гиатуса в значительной мере варьируется. В Припятском прогибе регрессия, начавшаяся еще в позднем фране, сменяется трансгрессией на рубеже зон Early — Middle triangularis.
На юго-востоке Русской плиты и в прилегающих районах Южного Урала регрессия фиксируется начиная с зоны linguiformis, при этом максимальное ее проявление приходится на время, соответствующее зоне triangularis. На западном склоне Среднего Урала регрессия, сменившая позднефранскую трансфессию, отмечена в терминальной части зоны linguiformis. Maксимум регрессии — в базальной части зоны
Early triangularis. Начиная с зоны Middle triangularis относительный уровень моря повышался [3]. В Тимано-Печорской провинции регрессия началась с зоны linguiformis, а максимальное ее развитие охватывает интервал, соответствующий зонам Early — Middle triangularis.
Ретроспективный анализ [5] исследований по девонским эвстатическим колебаниям в Северной Eвразии (Русская плита, Кузбасс, Колыма) и других регионах Земли выявил четко проявленную регрессию, которая приходится на конец зоны linguiformis и зону Early triangularis, и сменившую ее трансгрессию (Middle triangularis).
Колебания уровня Mирового океана на франско-фаменской границе и вблизи нее относятся к тектоноэвстати-ческим, связываются с развитием су-перплюмового события и сопровождающего его рифтогенеза. Глобальная регрессия и сменившая ее трансгрессия были следствием изменения строения дна океанического бассейна, обусловленного импульсом развития супер-плюмового процесса [5]. Предполагается, что к кратковременным глобальным изменениям уровня моря, вызывающим парное явление регрессия— трансгрессия, могли привести достаточно мощные процессы рифтогенеза. Изучение позднедевонского рифтоге-неза показало его глобальный характер. Он фиксируется на всех существовавших в то время континентах. В пределах Восточно-Eвропейской платформы позднедевонский рифтогенез и магматизм прослеживаются очень широко. В Тимано-Печорском регионе базальтовый вулканизм и рифтинг были достаточно кратковременными (конец живе-та — начало франа).
К событиям, сопряженным с глобальными процессами рифтогенеза, магматизма и эвстатических колебаний уровня Mирового океана, можно отнести формирование черносланцевых комплексов, характерную минералогению и геохимические аномалии.
Mаксимальное развитие позднедевонских (вторая половина франского и фаменский век) черносланцевых формаций наблюдается в эпиконтинентальных бассейнах Лавруссии [11]. Наиболее крупным является черносланцевый пояс на юго-востоке США, включающий Аппалачский, Mичиганский, Ил-линойсский бассейны и южную континентальную окраину. В другой части континента Лавруссия — в Тимано-Пе- чорском регионе и Предуралье — черные сланцы широко развиты в домани-ковом горизонте франа. С черносланцевыми формациями связан комплекс стратиформных месторождений с широким спектром рудных металлов. Эти формации имеют глобальное распространение и фиксируют в осадочных слоях Земли импульсы интенсивной дегазации земного ядра, при которых углеродсодержащие газы мигрировали вверх.
Показательны результаты исследования изотопного состава углерода. В непрерывных разрезах, изученных в самых разных регионах мира (в Западной и Восточной Eвропе, США, в Канаде, Австралии, Северной Африке), четко выделяются две положительные аномалии 13С, по стратиграфическому положению соответствующие двум кельвас-серским горизонтам. Причем аномалии фиксируются во всех разрезах, как в содержащих темноцветные кельвас-серские породы (в собственно кельвас-серских горизонтах), так и в чисто карбонатных, без признаков аноксийных обстановок. Предполагаются ситуации изотопного фракционирования углерода в мантии. Это могло быть в эпохи активного тектоногенеза. По-видимо-му, в процессе развития суперплюмо-вого события было два импульса восходящих флюидных потоков, достигших земной поверхности [17, 5]. Первый (основание зоны Late rhenana) флюидный поток, достигнув поверхности Земли, вызвал дестабилизацию условий в атмосфере и гидросфере (изменение климата, характера морских течений, стратификации океанских вод и др.) и, как следствие, изменение экологической обстановки и биособытия. Нестабильность экологических условий сохранялась длительное время и сопровождалась флуктуациями аноксийных обстановок, колебаниями уровня моря и др. Второй (терминальная часть зоны linguiformis), наиболее мощный импульс флюидных потоков, вероятно связанный с фазой рифтогенеза и началом глобальной регрессии, привел к массовому вымиранию морских организмов на франско-фаменском рубеже. Эти флюидные потоки привнесли в экзосферу не только обогащенный тяжелым изотопом углерод, но и ряд других элементов, влияющих на развитие биосферы. При опробовании непрерывных франско-фаменских разрезов отмечаются геохимические аномалии фосфора, рудных элементов [10]. К этому же периоду приурочен расцвет кремнистой биоты, свидетельствующий об обогащении морской воды кремнекисло-той [17].
Анализ возможных причин фран-ско-фаменского вымирания свидетельствует о продолжительном процессе изменения обстановок, на который влияло множество факторов, имеющих земные причины (в настоящее время точка зрения об импактном событии на франско-фаменской границе как первопричине массового вымирания не доказана).
БИОТИЧЕСКИЕ СОБЫТИЯ
Канадский геолог Д. Mакларен одним из первых обратил внимание на значительные биотические изменения на границе франского и фаменского ярусов [15]. По данным Д. Mакги [14], на этом рубеже уровень вымирания морских животных составил 3/4 видов, 1/2 родов и более 1/5 семейств.
Большим вкладом в оценку масштабов этого биособытия и его достаточно точной корреляции в различных регионах Земли стала разработка стандартных зон девона по конодонтам [19]. В результате изучения конодонтов из франско-фаменских отложений в США, Канаде, Бельгии, Германии, Франции, Нахичевани, Южном Китае и Австралии установлено массовое вымирание организмов на границе конодонтовых зон gigas (ныне linguiformis) и triangularis.
В пределах Рейнских сланцевых гор выделяют два горизонта черных кель-вассерских известняков и известковистых сланцев: нижний, основание зоны Late rhenana, и верхний в терминальной части зоны linguiformis. “Кельвассерс-кий кризис” начинается ниже зоны Late rhenana с исчезновения планктонной группы стилиолинид (гладкораковинных тентакулитов) и большей части семейств трилобитов. Промежуток времени, соответствовавший зонам Late rhenana – linguiformis, характеризовался глобальным диахронным исчезновением биогермных коралловых и стро-матопоратовых рифов во всех бассейнах мира. Э. Шиндлер [18] показал значительную продолжительность “кель-вассерского кризиса” и установил ступенчатый характер исчезновения различных таксонов организмов. По наблюдениям П. Коппера [10], связь между появлением черносланцевых горизонтов и исчезновением рифов отсутствует.
На западном склоне Южного Урала в большинстве разрезов к франско-фа-менской границе приурочен стратиграфический перерыв, известно только несколько полных разрезов, характеризующих различные фациальные обстановки. [16]. Существенные изменения в комплексе брахиопод происходят внутри брахиоподовых ракушняков (Early triangularis). На этом рубеже исчезает часть родов , составляющих основу зоны Late rhenana и зоны linguiformis. Некоторые роды — Schizophoria, Cyrto-spirifer, Athyris — продолжают существовать, но представлены другими видами. Изменения, происходившие выше, на границе зон Middle — Late triangularis, уступают по своей значимости нижнему рубежу; они отмечаются в основном на уровне видов.
В пределах западного склона Среднего Урала [3], так же как в ряде других районов мира, в самых верхах зоны

Рис. 2. Верхняя часть ухтинской свиты (обн. 19, р. Ижма)
linguiformis отсутствует сам вид Palmatolepis linguiformis, но увеличивается количество франских икриоду-сов. Возможно, это связано с обмелением бассейнов седиментации. Здесь (разрез “Баронская”, характеризующий отложения глубокого склона), как и в разрезах Южного Урала, непосредственно на франско-фаменской границе не фиксируются резкие литологические изменения, хотя по фауне эта граница устанавливается четко.
В Южном Китае [13] охарактеризованы относительно глубоководные разрезы рифтогенных депрессий наряду с мелководными разрезами межрифтовых пространств. Отмечены две фазы вымирания, аналогичные ступенчатому вы- 4
миранию в Германии. В первую фазу во второй половине зоны linguiformis, непосредственно у нижней границы черносланцевого горизонта, исчезает бентосная фауна (кораллы, брахиоподы, бентосные остракоды). Во вторую фазу на франско-фаменской границе вымирает пелагическая фауна и наблюдается расцвет кремнистых организмов. Биотические события сопровождаются резким повышением содержания некоторых металлов в осадочных породах. Предполагается, что причиной массового вымирания послужило аноксическое событие (скомбинированное с накоплением токсичных металлов), вызванное усилением рифтинга [13].
Однако далеко не во всех регионах Земли в это время наблюдались анок-сийные или гипоксийные обстановки. Так, Т. Беккер и M. Хауз, детально изучившие разрезы Canning Basin в Австралии, специально указывают на отсут- ствие этих обстановок на франско-фа-менской границе [8].
Анализ событий вымирания организмов в глобальном масштабе на фран-ско-фаменском рубеже свидетельствует о достаточно сложном, ступенчатом характере исчезновения отдельных групп фауны, отмеченного по крайней мере в двух последних конодонтовых зонах франского яруса (L. rhenana и linguiformis). Вместе с тем максимум вымирания, выраженный в исчезновении очень большого количества видов морских организмов, и затем появление новых видов приурочены именно к франско-фаменскому рубежу.
В отличие от неплохой изученности морской биоты, к настоящему мо- менту очень мало опубликовано данных об изменениях в континентальной биоте, происходивших на франско-фа-менском рубеже, в частности в растительных сообществах. Mакроостатки девонских растений и особенно их репродуктивные органы встречаются крайне редко, тогда как только они могут служить биологической основой для реконструкций филогении и эволюции первых наземных растений. Поэтому уникальным объектом исследований являются миоспоры, которые хорошо сохраняются в ископаемом состоянии. Изменения их морфологии и размеров в геологическом разрезе отражают эволюцию растений и четко фиксируют смену флористических сообществ в истории Земли.
С целью изучения миоспор из пограничных франско-фаменских отложений начиная с 1993 г., а в 2006 г. совместно с английским палинологом Д. Mаршаллом (рис. 2) проводились полевые экспедиционные работы на Южном Тимане. Здесь девонские отложения отличаются богатством и разнообразием фаунистических и микро-флористических остатков и поэтому являются классическим объектом для палеонтологических исследований. На Южном Тимане аналогом ливенского горизонта является сульфатная толща в объеме верхней части ухтинской свиты. Свита коррелируется с конодонто-выми зонами Upper и Uppermost gigas. Она подразделяется на две подсвиты: нижнюю, сложенную известняками до-ломитизированными, доломитами, мергелями с прослоями глин и песчаников, и верхнюю, представленную ангидритами и гипсами с прослоями глин и доломитов. В глубоководных аналогах данных отложений, образованных мергельно-глинистыми породами, определены конодонты зоны Palmato-lepis linguiformis [6].
В гипсоносных отложениях верхней части ухтинской свиты (рис. 2; Ухтинский район, пос. Крутой Кут, левый берег р. Ухта; Сосногорский район, пос. Сосновка, правый берег р. Ижма, напротив устья р. Ухта) установлен пали-нокомплекс с видом-индексом ливенского горизонта — Grandispora subsuta (Nazarenko) Obukh. В комплексе доминируют споры Cristatisporites deli-quescens (Naumova) Arkh ., Membra-baculisporis radiatus (Naumova) Arkh ., Auroraspora speciosa (Naumova) Obukh. и зафиксировано появление Geminospora vasjamica (Tchib.) Obukh.

et Nekr., вида-индекса фаменской па-линозоны C. vimineus—G. vasjamica. Ранее на этом стратиграфическом уровне виды G. subsuta и G. vasjamica в Тимано-Печорской провинции не отмечались.
Выше сульфатной толщи ухтинской свиты, отнесенной к ливенскому горизонту, на Южном Тимане залегает ижемская свита. Возраст ижемской свиты определен как раннефаменский, ее сопоставляют с волгоградским, задонским и елецким горизонтами Центрального девонского поля и конодонтовы-ми зонами triangularis—crepida [2].
В базальной части обн. 20 (стратотипа ижемской свиты), в котором глины темно-серые и зеленовато-серые переслаиваются с известняком светлосерым тонкоплитчатым (рис. 3), выделен палинокомплекс. Он отличается от известных ранее составом таксонов и их количественным соотношением. Eго особенностью является наличие видов Auroraspora sp . 1, Geminospora notata (Naumova) Obukh. var. microspinosus Tchib., Archaeozonotriletes famenensis Naumova var. minutus Naumova, Samarisporites lepidus Arkh., а также единичных спор, принадлежащих к родам, характерным для каменноугольных отложений — Tumulispora Staplin et Jansonius и Vallatisporites Hacq. В комплексе содержится значительное количество (до 60 %) франских видов миоспор. Другой его особенностью является обилие вариететов видов Membrabaculisporis radiatus (Naumova) Arkh . и Auroraspora speciosa (Naumova) Obukh., причем они значительно крупнее, чем исходные формы.
Новый палинокомплекс более близок к франским, чем к фаменским комплексам. В нем доминируют “франс-кие” таксоны, тогда как палинокомплекс перекрывающего его волгоградского горизонта фамена отличается почти полным отсутствием характерных фран-ских видов (однако до 40 % «фоновых» франских таксонов продолжали существовать и в фаменское время). Особенностью этого палинокомплекса являются также эндемизм и гигантизм миоспор, что, по-видимому, может свидетельствовать об островном характере флоры. Продуценты этой миоспоровой ассоциации, вероятно, произрастали в быстро меняющихся климатических условиях, например в условиях повышения гумидности климата при относительно постоянном температурном режиме.
Таким образом, благодаря четким отличительным признакам рассмотренного выше палинокомплекса возможно выделение нового биострати-графического подразделения. В шкале девонской зональности по миоспорам растений предлагается установить подволгоградские слои со стратотипом в разрезе ижемской свиты. Эти слои заполняют часть фиксирующегося на Русской платформе предфаменского стратиграфического перерыва и увеличивают объем франского яруса. По-видимому, новое биостратиграфиче-ское подразделение соответствует стратиграфическому интервалу, охарактеризованному в верхнедевонских разрезах Канады миоспоровой зоной Vallatisporites preanthoideus—Archae-ozonotriletes famenensis (конодонто-вым зонам linguiformis — нижней triangularis), и, возможно, нижней части зоны Membrabaculisporis radiatus—

Рис. 3. Базальные слои ижемской свиты (обн. 20, р. Ижма)
Cymbosporites boafeticus, установленной в восточной Померании (Польша) [9, 16].
Новый палинокомплекс демонстрирует постепенный характер изменений во флористических сообществах на уровне, коррелируемом с событием Kellwasser. Процесс вымирания позднефранских видов сопровождался появлением новых видов и родов миоспор, типичных для фаменских и даже каменноугольных палинокомп-лексов.
Обобщая результаты исследований по абиотическим и биотическим событиям на франско-фаменском рубеже, можно отметить, что изменения как в морской, так и в континентальной биотах имели сходный характер и подчинены были естественному эволюционному процессу. На скорость этих процессов, по-видимому, опосредованно влияли геологические события.
Список литературы Абиотические и биотические события на рубеже франского и фаменского веков
- Алексеев А.С. Типизация фанерозойских событий массового вымирания организмов // Вестн. Моск. ун-та. Сер. геол., 2000. № 5. С. 6-14.
- Безносов П.А., Хипели Д.В., Кузьмин А.В. и др. Литология, остатки позвоночных и конодонты ижемской свиты в стратотипе // Геология и минеральные ресурсы европейского северо-востока России: Материалы XIV Геол. съезда Республики Коми. Т. IV. Сыктывкар: Геопринт, 2004. С. 220-224.
- Бикбаев А.З., Снегирева М.П. Граница франского и фаменского ярусов на Среднем Урале по конодонтам // Девонские наземные и морские обстановки: от континента к шельфу (проект 499 МПГК / Международная комиссия по стратиграфии девона): Материалы Междунар. конф. Новосибирск, 2005. С. 35-36.
- Веймарн А.Б., Абрамова А.Н., Артюшкова О.В. и др. Корреляция разрезов фаменского яруса Южного Урала // Бюл. МОИП. Отд. геол., 2002. Т. 77. Вып. 1. С. 32-42.
- Веймарн А.Б., Корнеева С.А. Глобальные геологические события на рубеже франского и фаменского веков // Бюл. МОИП. Отд. геол., 2007. Т. 82. Вып. 1. С. 48-68.
