Адаптационные особенности девясила германского (Inula germanica L.)
")
Автор: Тамахина А.Я., Назранов Х.М., Локьяева Ж.Р.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Биологические науки
Статья в выпуске: 11, 2016 года.
Бесплатный доступ
Цель исследования - изучение механизмов адаптации девясила германского (Inula germa-nica L.) к засухе и засоленности почвы на ос-нове анатомо-морфологических особенностей и локализации продуктов вторичного мета-болизма. По уровню транспирации девясил германский относится к настоящим ксеро-фитам, по отношению к засоленности - к со-ленепроницаемым гликофитам. Основным ме-ханизмом засухо- и солеустойчивости явля-ется регуляция водного обмена. Морфологи-ческими и биохимическими приспособлениями девясила германского к засухе являются: ксе-роморфная структура листа (отсутствие устьиц на верхней поверхности листьев, толстая складчатая кутикула, двойная об-кладка проводящих пучков); накопление кри-сталлов оксалата кальция и простых сахаров в мезофилле листа; лигнификация открытых коллатеральных пучков стебля; внутренняя секреция эфирных масел в эпидермисе ложно-язычковых цветков; накопление каучука в ли-стьях и сердцевине корня; синтез антрацен-производных и дубильных веществ в корнях и стеблях, инулина - в корнях. Характерной особенностью строения стебля девясила германского является лигнификация тканей центрального цилиндра. Клетки нижнего эпи-дермиса листьев имеют извилистые стенки, устьица аномоцитного типа, железистые и простые волоски, эфирномасличные желѐзки. Клетки верхнего эпидермиса многоугольные или слабоволнистые. Кутикула эпидермиса с обеих сторон листа толстая, складчатая. Кристаллы оксалата кальция локализованы в неспециализированных кристаллоносных клет-ках мезофилла в виде крупных многогранных сростков сферической формы. Клетки эпи-дермиса ложноязычковых цветков удлиненные с прямыми стенками; эпидермис покрыт тол-стой кутикулой; железистые ткани пред-ставлены внутренними желѐзками (желези-стые пятна) и наружными головчатыми же-лезистыми волосками. Полученные данные расширяют представления о механизмах за-сухо- и солеустойчивости девясила герман-ского. Установленные анатомо-морфологи-ческие и фитохимические особенности можно использовать в качестве диагностических признаков при идентификации вида Inula ger-manica L.
Засу-хоустойчивость, солеустойчивость, продук-ты вторичного метаболизма, лист, корень, стебель, ложноязычковый цветок
Короткий адрес: https://sciup.org/14084533
IDR: 14084533 | УДК: 581.5:581.19:581.4:581.8
Adaptation features of German elecampane (Inula germanica L.)
The aim of the work is the study of mechanisms of German elecampane (Inula germanica L.) adap-tation to drought and soil salinity on the basis of anatomical and morphological features and locali-zation of products of secondary metabolism. On the level of transpiration German elecampane is the present xerothyte, in relation to salinity of the soil is impenetrable for salts glycophyte. The main mechanism of drought resistance and salt-endurance is water metabolism regulation. Mor-phological and biochemical devices of German elecampane by the drought are xeromorphic struc-ture of a leaf (lack of stoma on the upper face of leaves, a thick folded cuticle, double facing of con-ductive bunches); the accumulation of crystals of Sodium oxalatum of calcium and prime sugars in the leaf mesophyll; the lignification of open collat-eral bunches of the stalk; internal secretion of es-sential oils in epidermis in semiflorets; the accu-mulation of rubber in leaves and core of the root; the synthesis of anthracene-derivatives and tan-ning agents in roots and stalks, inulin in roots. The feature of the stalk structure is the lignification of fabrics of the central cylinder. The cells of the lower epidermis of the leaves have sinuous walls, the stomata of anomotsital type, glandular and simple hairs, oil droplets. The cells of upper epidermis are polygonal or undulating. The cuticle on both sides of the leaf is thick, folded. The crystals of Sodium oxalatum of calcium are localized in unspecialized crystalliferous cells of mesophyll in the form of large polyhedral aggregates of crystals of spherical shape. The cells of epidermis of semiflorets are oblong with straight walls; the epidermis is covered with a thick cuticle; there are internal oil droplets and external capitate glandular hairs. The data ob-tained expand the understanding of the mecha-nisms of drought resistance and salt-endurance of German elecampane. The set of anatomical-morphological and phytochemical features can be used as diagnostic features in identifying species of Inula germanica L.
Текст научной статьи Адаптационные особенности девясила германского (Inula germanica L.)
Введение. Девясил германский ( Inula ger-manica L.) – евро-западноазиатский вид лесостепной и степной зоны [1, 2]. Надземная масса девясила германского обладает антиоксидантной, гепатопротекторной и противовоспалительной активностью и применяется в народной и ветеринарной медицине [3, 4]. Химический состав растения изучен слабо. В подземной фитомассе содержатся эфирное масло, сескви-терпеноиды (германин), алакалоиды, а в надземной фитомассе – сесквитерпеноиды и каучук [3]. В траве и листьях девясила германского количественно определены флавоноиды и дубильные вещества [5, 6].
По отношению к температурному и водному режиму девясил германский является степным эвксерофитом, а по отношению к кальцию – облигатным кальцефилом [1, 7]. В КабардиноБалкарской Республике (КБР) девясил германский имеет узкий ареал распространения – участки типичных степей на карбонатных черноземах на севере и северо-востоке республики [8]. В связи с этим научный интерес представляет изучение механизмов адаптации Inula germanica L. к засухе и засоленности почвы.
Цель исследования : изучение механизмов адаптации Inula germanica L. к засухе и засоленности почвы на основе анатомоморфологических особенностей и локализации ряда биологически активных веществ в органах и тканях.
Объект, условия и методы исследования. Фитомасса девясила германского заготовлена в период цветения-плодоношения растений (август-сентябрь) 2015 г. и цветения (первая декада августа) 2016 г. в степной зоне КБР (с.п. Ал-туд). Климат степной зоны сухой и жаркий с выраженным периодом летнего иссушения. Среднегодовая температура воздуха +11,6 ºС, среднегодовое количество осадков – 533 мм. Почвы района исследования – черноземы южные карбонатные – тяжелосуглинистые, слабощелочные (рНвод 7,8); в горизонте 0–25 см содержание гумуса – 3,5 %; емкость поглощения – 29 ммоль (экв.)/100 г почвы; содержание по- движного фосфора и обменного калия среднее (соответственно 24,5 и 240,2 мг/кг почвы); карбоната кальция высокое (3,8 %). Засоление почв содово-сульфатное: НCO3- – 0,036 %; SO42- – 0,015 %; Cl- – 0,003 %.
Локализацию и идентификацию продуктов вторичного метаболизма проводили на свежих поперечных срезах корней, верхней и нижней части стеблей, а также на листьях и ложноязычковых цветках. Для исследования отобрано 50 растений. Количество срезов корней с каждого растения 10, стеблей – 20. Срезы изучали в 5-полях зрения микроскопа при увеличении 300× и 600×. Для качественного анализа использовали общепринятые гистохимические методы определения инулина, моно- и дисахаридов, дубильных веществ, антраценпроизвод-ных, смол, эфирных масел, лигнина, оксалата кальция, каучука [9]. Статистическая обработка включала расчет средней арифметической величины морфометрических параметров (высота побега, количество стеблевых листьев, число соцветий на 1 растение; длина и ширина листа, диаметр корзинки) и доверительного интервала по критерию Стьюдента для вероятности 0,95.
Результаты исследования. В исследованном фитоценозе девясил германский образует редкие и немногочисленные куртины в окружении субдоминантов длиннокорневищных и дерновинных злаков (овсяница валисская, щетинник сизый, тонконог гребенчатый, кострец безостый, пырей ползучий, сорго алеппское, житняк ломкий) . При раскопках подземной фитомассы установлен преимущественно вегетативный способ размножения растений девясила германского. Длинные ползучие корневища растений располагаются в верхнем слое почвы (0–20 см) и сильно разветвляются.
Высота растений девясила германского – 52,5±10,2 см. Стебель прямостоячий, продольно-ребристый, вверху ветвистый, равномерно облиственный. Количество листьев 25,6±3,2. Листья продолговато-ланцетные, жесткие, длиной 4,1±0,6 см, шириной 1,3±0,2 см, остроконечные, сидячие, в основании сердцевидные, полустеблеобъемлющие, с мелкими зубчиками. С нижней стороны листа выделяются жилки. Жилкование перистое. Корзинки (4,3± 1,1 шт.) диаметром 0,91±0,13 см собраны в щиток. Цветки желтые; ложноязычковые цветки слегка превышают обертку. Обертка цилиндрическая, черепитчатая. Плоды – цилиндрические ребристые голые семянки с хохолком из много- численных длинных щетинок.
Стебель девясила германского имеет пучковый тип строения с заложением межпучкового камбия. В верхней части стебель покрыт эпидермисом с простыми многоклеточными волосками. В средней и нижней части стебля эпидермис сменяется многослойной перидермой. Между ксилемой и флоэмой лежит камбиальная
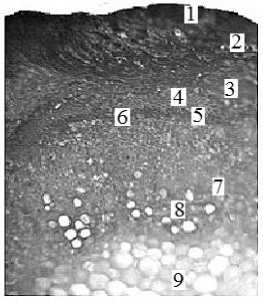
а зона, состоящая из прямоугольных клеток, расположенных правильными радиальными рядами. Участки межпучкового камбия примыкают к пучковому камбию, образуя сплошной камбиальный слой. Межпучковый камбий дифференцируется в паренхиму сердцевинных лучей. Сердцевина состоит из рыхло расположенных паренхимных клеток (рис. 1, а).
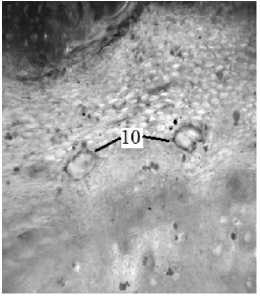
б
Рис. 1. Поперечный срез стебля (а) и корня (б): 1 – перидерма; 2 – первичная кора;
3 – склеренхима перицикла; 4 – флоэма; 5 – пучковый камбий; 6 – межпучковый камбий;
7 – сердцевинный луч; 8 – ксилема; 9 – паренхима сердцевины; 10 – схизогенное вместилище. Окраска резорцином (а) и суданом III (б). Увел. 300×
По результатам окрашивания резорцином с соляной кислотой установлено, что лигнифика-ции подвергаются элементы ксилемы, флоэмы и сердцевинных лучей по всей длине стебля, что обеспечивает механическое укрепление ткани и защиту клеток от химических, физических и биологических воздействий.
В стеблях и корнях содержатся антраценпро-изводные и дубильные вещества. Желтое окрашивание гидроксидом калия свидетельствует о том, что антраценпроизводные вещества представлены восстановленными мономерами – производными антранола, антрона, оксиантрона и их производных. Антраценпроизводные, дубильные вещества и инулин локализованы в коровой паренхиме и сердцевине корня. Антрацен-производные выполняют функцию защиты от паразитов, играют определенную роль в окислительно-восстановительных процессах растений, а также способствуют накоплению полисахаридов. Дубильные вещества благодаря большому количеству фенольных гидроксилов обладают выраженными бактериостатическими и фунгицидными свойствами; участвуют в окислительно-восстановительных процессах, являясь переносчиками кислорода в растениях. Инулин выполняет функцию энергетического запаса в корнях [10].
Смолы и эфирные масла накапливаются в схизогенных вместилищах корней и корневищ (рис. 1, б), а также в млечном соке листьев. Биологическая функция смол связана с защитой растения от фитопатогенов в условиях водного стресса [10].
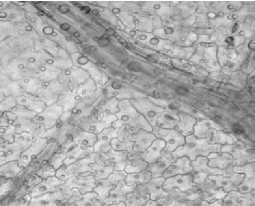
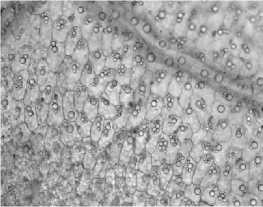
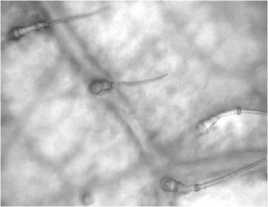
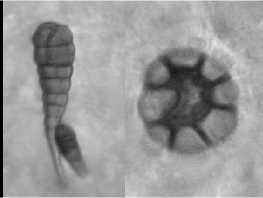
Листья девясила германского имеют ксеро-морфную структуру. Клетки нижнего эпидермиса извилистые (рис. 2, а), с верхнего – многоугольные или слабоволнистые (рис. 2, б). Расположение устьиц на нижней поверхности листа равномерно рассеянное; устьица аномоцитного типа с 4 околоустьичными клетками. На нижнем эпидермисе и жилках рассеянно расположены кроющие трихомы – простые 3-5-клеточные волоски с расширенным основанием (рис. 2, в), железистые волоски с одноклеточной ножкой и многоклеточной головкой и реже – сидячие эфирномасличные железки (рис. 2, г).
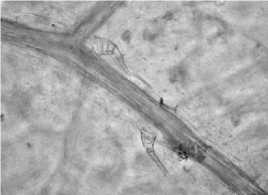
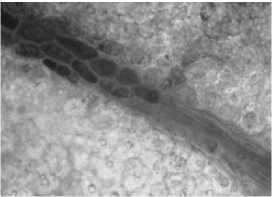
Кутикула складчатая, в нижней части листа более толстая, что обеспечивает защиту от тепловых повреждений. Центральная проводящая жилка хорошо выражена. Проводящие пучки окружены склеренхимой, выполняющей механическую функцию, и двухслойной обкладочной паренхимой. Обкладки пучков в листьях состоят из клеток, удлиненных в направлении, параллельном ходу пучка, содержат хлоропласты.
С нижней стороны листа обкладочные клетки мелкие (рис. 2, д), с верхней стороны – более крупные (рис. 2, е). Обкладки простираются до окончания жилок и окружают конечные трахеиды. Кутикуляризация клеток эпидермы и защита проводящих пучков листа слоями обкладочных клеток являются ксерофитными признаками растений [11].
По краю листа расположены 3–5-клеточные железистые волоски с толстой кутикулой и одноклеточной заостренной легко обламывающейся конечной клеткой (рис. 2, ж). Эфирное масло выделяется наружу при отпадании конечной клетки волоска. Капли эфирного масла обнаружены и в протопласте клеток эпидермиса.
Основными вторичными метаболитами листьев девясила германского являются эфирные масла, смолы, моно- и дисахариды, кристаллы оксалата кальция и каучук, играющие важную роль в формировании засухоустойчивости растения. Так, испарение эфирных масел листьями обуславливает уменьшение теплопроница-емости воздуха, что способствует предохранению растения от чрезмерного нагревания и переохлаждения, а также обеспечивает регуляцию транспирации [12]. Синтез сахаров-осмолитов является механизмом поддержания высокого осмотического давления в клетках в условиях почвенного засоления и засухи [13].
а
б
в
г
д
е
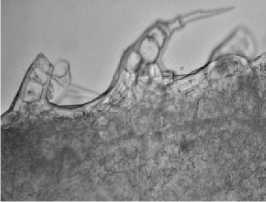
ж
Рис. 2. Лист девясила германского: а – клетки нижнего эпидермиса; б – клетки верхнего эпидермиса; в – кроющие трихомы нижнего эпидермиса; г – эфирно-масличные железки; д – жилка на нижней поверхности; е – жилка на верхней поверхности; ж – краевые волоски.
Окраска Суданом III (г). Увел. 600 х
В клетках мезофилла листьев, вокруг жилок, по краю листа локализованы многочисленные неспециализированные кристаллоносные клетки с друзами оксалата кальция в виде крупных многогранных сростков коротких, иногда остроконечных кристаллов, имеющих в очертании более или менее сферическую форму (рис. 3, а).
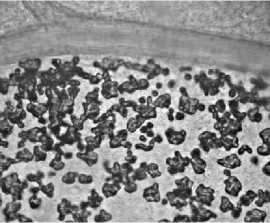
а

б
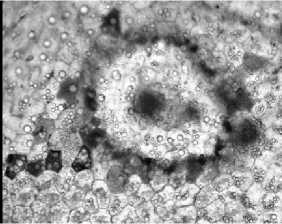
в
Рис. 3. Кристаллы оксалата кальция в кристаллоносных клетках (а), глобулы каучука в млечниках (б) и в местах повреждения (в) листа. Окраска суданом III (б-в).
Увел. 600 * (а, в), 300 * (б)
Кристаллы оксалата кальция активно вовлекаются в процессы детоксикации избытка поступившего в растения кальция [14], участвуют в осморегуляции [15], обеспечении жесткости тканей [16], защите растений от патогенов и травоядных [17], детоксикации ионов тяжелых металлов [18], формировании ионного гомеостаза [19].
Проводящие пучки листьев сопровождают членистые млечники, в латексе которых суспендированы частицы каучука (рис. 3, б). Глобулы каучука сероватого и желтовато-бурого цвета различных размеров хорошо заметны при осветлении листьев жавелевой водой. Установлено наличие глобул каучука в сердцевине корней. Терпеноиды, составляющие основу каучука, играют важную роль в процессах обмена веществ, некоторые из них являются гормонами и регуляторами жизнедеятельности растений [10]. Основная функция каучука – герметизация повреждений тканей листа (рис. 3, в).
Клетки эпидермиса ложноязычковых цветков удлиненные, с прямыми стенками, покрыты толстой кутикулой. Для цветков характерна внутренняя и наружная секреция эфирного масла. Внутренние железки – одиночные клетки и железистые пятна – расположены в эпидермальной ткани (рис. 4, а). Испарение эфирных масел при отслаивании кутикулы является одним из механизмов регуляции транспирации растения. Наружная секреция осуществляется рассеянно расположенными головчатыми железистыми волосками с одно- или многоклеточной ножкой и одно- или многоклеточной головкой (рис. 4, б).
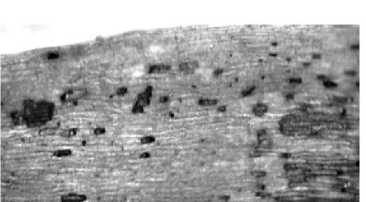
а
Рис. 4. Эфирномасличные железки ложноязычкового цветка: а – железистые пятна эпидермиса; б – железистые волоски. Окраска метиленовым синим. Увел. 300× (а), 600× (б)
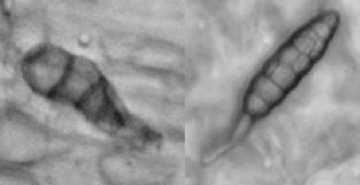
б
Выводы. Исследованы анатомические, морфологические и биохимические особенности Inula germanica L. в связи с адаптацией к произрастанию в степных фитоценозах.
Характерной особенностью строения стебля девясила германского является значительная степень лигнификации тканей центрального цилиндра, включая сердцевинные лучи. Клетки нижнего эпидермиса листьев имеют извилистые стенки, устьица аномоцитного типа, железистые волоски по краям листа и простые волоски по всей поверхности. Клетки верхнего эпидермиса многоугольные или слабоволнистые. Кутикула эпидермиса с обеих сторон листа толстая складчатая. Многочисленные кристаллы оксалата кальция локализованы в не- специализированных кристаллоносных клетках мезофилла в виде крупных многогранных сростков кристаллов, имеющих в очертании сферическую форму. Клетки эпидермиса ложноязычковых цветков удлиненные, с прямыми стенками; эпидермис покрыт толстой кутикулой; железистые ткани представлены внутренними железками (железистые пятна) и наружными головчатыми железистыми волосками.
По уровню транспирации девясил германский относится к настоящим ксерофитам (эвксерофитам), о чем свидетельствуют следующие анатомо-морфологические признаки: сильно разветвленная корневая система, расположенная в почве на небольшой глубине, небольшие жесткие листья, толстая кутикула эпидермиса листьев и цветков, двойная обкладка проводящих пучков. Продукты вторичного метаболизма (каучук, эфирные масла, ан-траценпроизводные, инулин, лигнин, смолы, дубильные вещества, кристаллы оксалата кальция) имеют адаптивное значение и играют защитную роль в условиях продолжительной засухи и высоких температур. По отношению к засоленности почвы девясил германский является соленепроницаемым гликофитом, по обитанию преимущественно на определенном субстрате – кальцексерофитом.
В связи с существованием тесной положительной корреляции между солеустойчивостью и засухоустойчивостью растений девясил германский, не имея эволюционно сложившихся механизмов устойчивости к засолению почвы, приспосабливается к ним за счет эффективных механизмов засухоустойчивости, важнейшим из которых служит способность к регуляции водного обмена.
Полученные данные расширяют представления о механизмах засухо- и солеустойчивости девясила германского . Установленные анатомоморфологические и фитохимические особенности можно использовать в качестве диагностических признаков при идентификации вида Inula germanica L.
Список литературы Адаптационные особенности девясила германского (Inula germanica L.)
- Губанов И.А., Киселева К.В., Новиков В.С. Дикорастущие полезные растения. -М.: Изд-во МГУ, 1993. -300 с.
- Флора Европейской части СССР/отв. ред. Н.Н. Цвелев. -СПб.: Наука, 1994. -Т. VII. -371 с.
- Растительные ресурсы СССР. Цветковые растения, их химический состав, использо-вание. Семейство Asteraceae (Compositae). -СПб.: Наука, 1993. -352 с.
- Шагиахметов Ю.С. Фармакологическая характеристика препаратов растений рода Inula и их применение в ветеринарной ме-дицине: автореф. дис.. д-ра вет. наук. -Троицк, 2000. -36 с.
- Яницкая А.В., Митрофанова И.Ю. Разра-ботка методики количественного определе-ния флавоноидов в траве девясила герман-ского//Научные ведомости. Сер. «Медици-на. Фармация». -2013. -№ 18 (161). -Вып. 23. -С. 211-214.
- Количественное определение дубильных веществ травы девясила германского/А.С. Рабичева, А.В. Яницкая, В.Н. Емцева //Итоги научно-исследовательской деятельности 2015 г.: изобретения, методи-ки, инновации: мат-лы VI Междунар. науч.-практ. конф. -М.: Перо, 2015. -С. 441-444.
- Кухарева Л.В., Пашина Г.В. Полезные травя-нистые растения природной флоры: справоч-ник по итогам интродукции в Белоруссии. -Минск: Наука и техника, 1986. -215 с.
- Тамахина А.Я., Гадиева А.А., Локьяева Ж.Р. Эколого-фитоценотическая характеристика девясила германского (Inula germanica L.) в степных фитоценозах Кабардино-Балкарской Республики//Современные проблемы науки и образования. -2016. -№ 4. -URL: www.science-education.ru/ru/issue/view?id=141.
- Прозина М.Н. Ботаническая микротехника. -М.: Высш. шк., 1960. -205 с.
- Основы биохимии вторичного обмена рас-тений: учеб.-метод. пособие/Г.Г. Борисо-ва, А.А. Ермошин, М.Г. Малева . -Екатеринбург: Изд-во Урал. ун-та, 2014. -128 с.
- Седельникова Л.Л. Анатомическое строе-ние листа у Hemerocallis hybrid//Вестн. КрасГАУ. -2016. -№ 7. -С. 55-61.
- Эфирные масла//Большая Советская эн-циклопедия (в 30 т.)/Гл. ред. А.М. Прохо-ров. -3-е изд. -М.: «Советская Энциклопедия, 1978. -Т. XXX. -С. 320-321. -632 с.
- Стрессорный ответ клеток Nicotiana syl-vestris L. на засоление и высокую темпера-туру. 1. Аккумуляция пролина, полиаминов, бетаинов и сахаров/Н.И. Шевякова, Б.В. Рощупкин, Н.В. Парамонова //Физиология растений. -1994. -Т. 41, № 4. -С. 558-565.
- Zindler-Frank E., Honow R., Hesse A. Calcium and oxalate content of the leaves of Phaseolus vulgaris at different calcium supply in relation to calcium oxalate crystal formation//J. Plant Physiol. -2001. -V. 158. -P. 139-144.
- Ruiz L.P., Mansfield T.A. A postulated role for calcium oxalate in the regulation of calcium ions in the vicinity of stomatal guard cells//New Phytol. -1994. -V. 127. -P. 473-481.
- Hudgins J.W., Krekling Т., Franceschi V.R. Distribution of calcium oxalate crystals in the secondary phloem of conifers: a constitutive defense mechanism?//New Phytologist. -2003. -V. 159. -P. 677-690.
- Dias В.В.A., Cunha W.G., Morais L.S. et al. Expression of an oxalate decarboxylase gene from Flammulina sp. in transgenic lettuce (Lactuca sativa) plants and resistance to Sclerotinia sclerotiorum//Plant Pathology. -2006. -V. 55. -P. 187-193.
- Мазен А.М.А. Связь отложений оксалата кальция в листьях джута (Corchorus olitori-us) с накоплением токсичных металлов//Физиология растений. -2004. -Т. 51, № 2. -С. 314-319.
- Попова Н.Ф. Роль оксалата в формирова-нии ионного гомеостаза в листьях Amaran-thus cruentus L.: автореф. … канд. биол. наук. -СПб., 2009. -18 с.
