Адаптационные возможности интродуцентов различного происхождения в условиях Ботанического сада г. Сургута

Автор: Турбина И.Н., Кукуричкин Г.М.
Журнал: Вестник Нижневартовского государственного университета @vestnik-nvsu
Рубрика: Экология растений
Статья в выпуске: 2, 2020 года.
Бесплатный доступ
Ботанический сад в г. Сургуте является самым северным в Западной Сибири и одним из самых северных ботанических садов в России, он является основным форпостом для продвижения новых агро- и биотехнологий на Крайний Север. В настоящее время коллекция декоративных растений Сургутского ботанического сада насчитывает около 200 видов и сортов травянистых растений, относящихся к 60 родам и 32 семействам, и около 210 видов, форм и культиваров древесных растений. Целью работы являлось изучение механизмов адаптации некоторых декоративных растений из коллекции ботанического сада с использованием современного портативного флавонид- и хлорофиллометра DUALEX. Объекты исследований - пять представителей травянистых растений из рода Iris L. - Iris hungarica Waldst. &Kit., Irisvariegata L., Iris sibirica L., Iris glaucescens Bunge, Iris sulfurea C. Koch. и восемь видов древесных растений из рода Acer L. - Acer pseudoplatanus L., Acer rubrum L., Acer trautvetteri Medw., Acer tataricum L., Acer ukurunduense Trautv...
Адаптация, биохимические показатели, ботанический сад, древесные растения, травянистые растения
Короткий адрес: https://sciup.org/14117224
IDR: 14117224 | УДК: 581.5:635.9 | DOI: 10.36906/2311-4444/20-2/08
Adaptation possibilities of introduced plants of various origin in conditions of Botanical garden at Surgut State University
The botanical garden in Surgut is the northernmost in Western Siberia and one of the northernmost botanical gardens in Russia. It is the major point for promoting new agricultural and biological technologies to the Far North. Currently, the collection of ornamental plants of the Surgut botanical garden comprises about 200 species and varieties of herbaceous plants belonging to 60 genera and 32 families, and about 210 species, forms and cultivars of woody plants. The research objective is to study the adaptation mechanisms of some ornamental plants from the collection of the botanical garden using the up-to-date portable flavonoid and chlorophyll meter DUALEX. The research objects are five species of herbaceous plants of genus Iris L. - Iris hungarica Waldst. &Kit., Iris variegata L., Iris sibirica L., Iris glaucescens Bunge, Iris sulfurea C. Koch. and eight species of woody plants of genus Acer L. - Acer pseudoplatanus L., Acer rubrum L., Acer trautvetteri Medw., Acer tataricum L...
Текст научной статьи Адаптационные возможности интродуцентов различного происхождения в условиях Ботанического сада г. Сургута
В последние годы приобрели актуальность исследования содержания растительных пигментов с использованием спектров отражения для получения количественных данных в пространственных и временных масштабах. За рубежом развиты исследования в области моделирования пигментов растений и их диагностических функций с использованием оптических методов дистанционного зондирования [16; 17; 18]. Пигменты растений (хлорофилл, флаваноиды, антоцианы) выполняют защитные функции, предохраняют растения от различных неблагоприятных воздействий [15]. Состояние пигментного комплекса влияет на устойчивость растительных организмов к стрессу. Стресс – это интегральный неспецифический ответ растения на повреждающее действие, направленный на выживание организма за счет мобилизации и формирования защитных систем. Устойчивость является конечным результатом адаптации [5].
Интродукционные исследования проводили в условиях Сургута, который расположен в среднетаежной зоне Западно-Сибирской равнины, характеризующейся резко континентальным климатом, с холодной продолжительной зимой и теплым коротким летом. По обеспечению влагой район находится в области достаточного увлажнения. Находясь на севере Азии, территория испытывает влияние влажных воздушных масс с Северного Ледовитого океана. Поэтому зима относительно сухая и морозная. В летний период доминируют северные ветра, однако в ясную погоду воздушные массы успевают прогреваться.
Ботанический сад в г. Сургуте является самым северным в Западной Сибири и одним из самых северных ботанических садов в России, он является основным форпостом для продвижения новых агро- и биотехнологий на Крайний Север, в т. ч. в арктические регионы, на принципах ступенчатой акклиматизации ценных видов и сортов растений.
В настоящее время коллекция декоративных растений Сургутского ботанического сада насчитывает около 200 видов и сортов травянистых растений, относящихся к 60 родам и 32 семействам, и около 210 видов, форм и культиваров древесных растений, она сформирована по систематическому принципу (родовыми комплексами), что позволяет сравнивать между собой морфологические особенности видов в пределах рода и семейства. Все растения зимуют в открытом грунте, без укрытия.
В культуре ботанического сада насчитывается 15 видов рода Iris L. и 10 культиваров Iris sibirica L., полученные из ботанических садов Новосибирска, Пензы, Махачкалы, Пятигорской эколого-ботанической станции БИН РАН.
Этот травянистый многолетник исключительно многообразен по количеству видов, сортов, диапазону цветовой гаммы. В природе насчитывается более 200 видов ирисов, распространенных в умеренных и отчасти в субтропических широтах всех континентов северного полушария – на открытых, солнечных местах, лишь небольшая часть видов – растения теневых и даже заболоченных местообитаний [9]. Во флоре России встречается около 60 видов [1].
Род Acer L. в коллекции представлен 49 экземплярами 15 видов различного географического происхождения. Всего в природе насчитывается около 150 видов, которые распространены в Европе, в Передней и Средней Азии, в Гималаях, Восточной Азии, в Северной и Центральной Америке. Многие виды этого рода довольно быстро растут, неприхотливы в уходе и обладают более высокой аккумулирующей способностью тяжелых металлов (свинец, кадмий) в листьях взрослых растений по сравнению с другими породами [10].
Целью исследования являлось изучение механизмов адаптации некоторых декоративных растений из коллекции ботанического сада СурГУ с использованием современного портативного флавонид- и хлорофиллометра DUALEX.
Материалы и методы исследования
Объектами исследований являлись пять представителей травянистых растений из рода Iris L. семейства Iridaceae Juss.
Iris hungarica Waldst. &Kit. (syn. Iris versicolor L.) (И1) – ирис разноцветный, является представителем североамериканской флоры. Произрастает на сырых лугах и болотах, по берегам озер и рек, в кустарниках теплоумеренной и субтропической зон северо-востока Канады и США [13]. Корневище ползучее, 8–10 см толщиной, образует рыхлую дерновину. Листья прикорневые, утолщенные посередине, но без резко выступающей средней жилки, к основанию с красноватым оттенком, 1,5–2,5 см шириной, 35–45 см длиной. Этот вид был привезен в 2018 г. из Горного ботанического сада (г. Махачкала).
Iris variegata L. (И2) – ирис пестрый, произрастает на сухих каменистых склонах, среди кустарников, в редколесьях, на лесных полянках, по опушкам дубрав юга теплоумеренной зоны Средней Европы, Балкан. Корневище 1–2 см толщиной. Прикорневые листья широко- или линейно-мечевидные, 15–25 см шириной, прямые или слегка серповидно изогнутые, с заметной продольной ребристостью в средней части пластинки, равны или ниже цветоноса. Один из первых видов, использованных при создании культурных форм ириса [7].
Iris sibirica L. (И3) – ирис сибирский, произрастает в Европе, Предкавказье, Малой Азии и Западной Сибири. Обитает на пойменных и лесных лугах, в березовых колках. Плотнодерновин-ный поликарпик [14]. Стебли 60–135 см высоты. Листья линейные, заостренные, короче стебля. Отрастает в начале июня, цветение во второй половине июля, в течение двух недель, вегетирует до снежного покрова. В культуре неприхотлив, устойчив. В коллекции три экземпляра, с различной окраской венчика, полученных в 2018 г. из Центрального сибирского ботанического сада (г. Новосибирск).
Iris glaucescens Bunge (И4) – ирис сизоватый, произрастает в полынно-дерновиннозлаковых степях на солонцеватых песках, сухих каменистых и щебнистых склонах немногочисленными популяциями на юге Западной Сибири, в Казахстане, Монголии и Китае. Рыхлокустовой поликар-пик. Листья серповидно изогнуты, интенсивно сизые. В коллекции два экземпляра, поступивших в 2018 г. из Горного ботанического сада (г. Махачкала).
Iris sulfurea C. Koch. (syn Iris inbricata Lindl.) (И5) – ирис черепитчатый, встречается на горных лугах, на достаточно увлажненных (проточное увлажнение) каменистых склонах, нередко по обрывистым берегам горных рек в юго-восточных районах Закавказья, в Северном Иране, на высоте 800–2000 м н.у.м. Плотнодерновинный поликарпик. Он имеет широкие, мечеобразные, желто-зеленые или светло-зеленые листья до 30–40 см в длину и от 2 до 3 см в ширину; тонкий стебель овальной формы, с ветвями. Три экземпляра этого вида в 2019 г. получены из Пятигорской эколого-ботанической станции БИН РАН [8].
Вторую группу объектов исследования составили восемь видов древесных растений из рода Acer L. семейства Sapindaceae Juss. различного географического происхождения [3; 4; 6; 11].
Acer pseudoplatanus L. (К1) – клен ложноплатановый, или белый, или явор. Распространен в широколиственных лесах Южной и Центральной Европы, юго-западной Украины и Кавказа. В естественных условиях – высокоствольное дерево. При интродукции в северных регионах часто принимает форму кустарника. Листья до 15–17 см в диаметре, с 5 крупными округлыми лопастями, с верхней стороны темно-зеленые, с нижней – сизовато-белые, обычно волосистые. Требователен к почвенно-климатическим условиям. 2 экземпляра получены из Санкт-Петербурга в 2012 г. и 5 – из Светлогорска (Калининградская область) в 2016 г. Регулярно обмерзают до уровня снега, но интенсивно возобновляются. 4 группа зимостойкости.
Acer rubrum L. (К2) – клен красный Североамериканское не крупное дерево. Растет в лесах по берегам рек и болот, в ущельях и на сухих пониженных участках. Среднетеневынослив. Листья простые 3–5-лопастные, крупные (5–15 на 3,5–10 см), осенью желтые, оранжевые до яркопурпурных. 1 экземпляр получен из Санкт-Петербурга в 2013 г. 2–3 группа зимостойкости.
Acer trautvetteri Medw. (К3) – клен Траутфеттера, или высокогорный. Естественно произрастает в высокогорьях Кавказа. Небольшое дерево с толстым, саблевидно изогнутым в нижней части стволом. Крона широкояйцевидная или шаровидная. Теневынослив. Листья простые, в очертании широкояйцевидные, 9–14 см длиной и 11–16 см шириной, глубоко пятилопастные. Осенью листья приобретают ярко-красно-карминовую окраску. 2 экземпляра получены в 2019 г. из Горного ботанического сада (Дагестан). Группа зимостойкости пока не известна.
Acer tataricum L. (К4) – клен татарский, или черноклен, растет в европейской части России, на Кавказе, в Малой Азии, Иране; на опушках и в окнах широколиственных лесов, чаще на выруб- ках, на высоких гривах в поймах рек. Небольшое дерево, обычно кустовидное. Листья цельные или слегка лопастные, 6–10 см длиной, яйцевидные, заостренные, по краю двоякопильчатые, сверху темно-зеленые, снизу светлее и по жилкам волосистые. Осенью от желтых, оранжево-желтых и пурпурных до коричневых. Теневынослив. Изредка встречается в озеленении и регулярно плодоносит. В коллекции Сургутского ботанического сада имеется несколько экземпляров, поступивших из Уфы в 2012 г., из Санкт-Петербурга в 2014 г., а также полученные из семян сургутской репродукции. 1 группа зимостойкости.
Acer ukurunduense Trautv. &C.A. Mey. (К5) – клен желтый, или укурундский. Ареал: Российский Дальний Восток, Япония, северо-восток Китая. Раскидистый кустарник или кустовидное дерево. Листья пальчато-пятилопастные, довольно жесткие, длиной 8–12 см и почти такой же ширины, опушенные, средняя лопасть значительно крупнее боковых. Осенью листва приобретает красивую яркую окраску, оттенки которой варьируются от светло-красных до коричневатооранжевых и фиолетовых. Хорошо переносит затенение и в природе обычно встречается в верхнем кустарниковом ярусе хвойных лесов. Самый морозостойкий из дальневосточных кленов. 5 экземпляров получено из Санкт-Петербурга в 2016 г. 2–3 группа зимостойкости.
Acer spicatum Lam. (К6) – клен колосистый, является самым северным из видов клена, произрастающих в Северной Америке. Растет на опушках и в подлеске смешанных лесов, довольно хорошо переносит сильное затенение. В естественных условиях – кустарник или небольшое дерево. Листья простые, широко яйцевидные, 7–10 см длиной и 6–9 см шириной, со слабо сердцевидным основанием, неглубоко трех- или пятилопастные, толстые, мягкие, снизу опушены. Осенью листья окрашиваются в желто-оранжевые тона. 3 экземпляра получены из Уфы в 2012 г. 2 группа зимостойкости.
Acer mandshuricum Maxim. (К7) – клен маньчжурский, в природе растет в смешанных лесах на юге Российского Дальнего Востока, в Корее, Японии, Китае. Растет в долинных широколиственных и смешанных лесах, где местами господствует во 2 ярусе. Светолюбив. Дерево, часто кустовидное, с тройчатыми листьями, листочки длиной до 8 см и до 2,5 см шириной на длинных красноватых черешках. Осенью листья становятся пурпурно-красными. В коллекции Сургутского ботанического сада имеется два экземпляра, поступивших из Санкт-Петербурга в 2014 г. и Новосибирска в 2018 г. 2-3 группы зимостойкости.
Acer ginnala Maxim. (К8) – клен гиннала, или приречный, произрастает в широколиственных лесах Российского Дальнего Востока, в Японии и Китае. Деревья 5–7 м высотой, с трехлопастными листьями, центральная лопасть более длинная, боковые – короче, горизонтально распростертые или несколько вверх направленные, нередко почти цельные, яйцевидной формы. Осенью листья становятся малиново-красными. 3 экземпляра получены из Уфы в 2012 г. 1 группа зимостойкости. Цветет и плодоносит.
Для оценки повреждаемости древесных растений низкой температурой использовалась 5-балльная шкала С.Я. Соколова [12]: 1 – растение вполне зимостойко; 2 – у растения отмерзают концы побегов; 3 – отмерзают крупные ветви; 4 – отмерзает вся надземная часть до уровня снегового покрова (или почвы); 5 – растение не зимует, вымерзает с корнем.
Определение биохимических показателей (содержание флавонолов – Flv, мг/см², хлорофилла – Chl, мг/см² и индекса азотного баланса – Nbi) проводили с помощью инновационного аппарата DUALEX (Франция). Индекс азотного баланса растений Nbi (Nitrogen Balance Index) представляет собой соотношение количества хлорофилла и флавонолов. Выборку листьев делали с нескольких близко растущих растений, для древесных на уровне поднятой руки, с максимального количества доступных веток, направленных условно на север, запад, восток и юг.
Статистическая обработка данных осуществлялась при помощи программных пакетов Excel MS Office-2016 и Statistica 10. Соответствие структуры данных закону нормального распределения оценивалось на основе критерия Шапиро–Уилка (для выборок n < 30). Производилась идентификация показателей Nbi, Chl, Flv на соответствие закону нормального распределения. Закон Гаусса подтвердился, поэтому дальнейшие исследования зависимостей производились методами параметрической статистики. Для сравнения двух независимых ненормально распределенных выборок использовали U-тест Манна-Уитни (Mann-Whitney U-test), который сравнивает не средние значения выборок, а суммы рангов по каждой из них. Сравнение двух независимых групп осуществлялось с использованием параметрического критерия t-тест Стьюдента для оценки наличия различий между ними. Для сравнения нескольких независимых выборок видов клена по одному признаку использовали непараметрический дисперсионный анализ Крускала–Уоллиса и медианный тест (ANOVA Kruskal-Wallisand Median test).
Результаты и их обсуждение
Диапазон азотного баланса у травянистых растений составил от 32,2 до 39,7, а содержание хлорофилла – от 48,1 до 58,6мг/см2, с максимальным значением этих двух показателей у Iris hun-garica (рис. 1). Это связано с действием стресса, вызванного нарушением водного режима растительного организма ириса разноцветного, который предпочитает довольно увлажненные местообитания, а в культуре недостаточное водоснабжение привело к нарушению экологических требований растений, что согласуется с литературными источниками [2]. Показатели содержания (Nbi,
Chl) в листьях травянистых растений варьировали больше, чем содержание флавонолов, которое
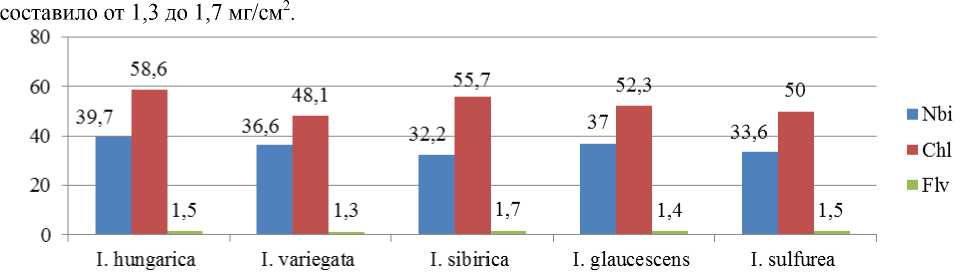
Рис. 1. Сравнительный анализ средних значений показателей (Nbi, Chl, Flv) в листьях травянистых растений рода Iris (август)
Аналогично у древесных растений максимальные значения (Nbi, Chl) отмечены у Acer pseudoplatanus, что является реакцией адаптации к условиям произрастания и указывает на повышение защитных функций растения (рис. 2). Стресс вызван климатическими факторами, так как явор регулярно обмерзает до уровня снега, а затем интенсивно возобновляется за вегетационный период.
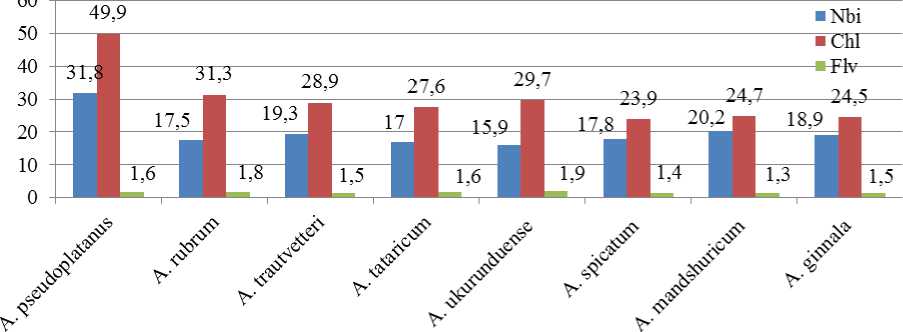
Рис. 2. Сравнительный анализ средних значений показателей (Nbi, Chl, Flv) в листьях древесных растений рода Acer (август)
Значение индекса азотного баланса (NBI) у остальных изученных видов изменялось незначительно – от 15,9 ( Acer ukurunduense ) до 19,3 ( Acer trautvetteri ). Данный показатель зависит от многих экологических факторов, а в нашем случае минимальное значение (NBI) клена укурунд-ского является адаптационной реакцией к условиям достаточного освещения, так как он произрастает на открытом месте, без затенения.
Содержание хлорофилла варьировало от 23,9 мг/см2 ( Acer spicatum ) до 31,3 мг/см2 ( Acer rubrum ), т. е. всего в 1,3 раза в пределах видов группы, произрастающих на одной экспозиции (в одинаковых экологических условиях).
Содержание флавонолов в листьях древесных растений было невелико и варьировало от 1,3 мг/см2 у Acer mandshuricum до 1,9 мг/см2 у Acer ukurunduense .
Таким образом, исследованные группы неодинаково реагируют на условия среды произрастания. Пигментный состав листьев декоративных растений свидетельствует об их относительно высоком физиологическом статусе.
В полученных результатах достоверными были различия по содержанию хлорофилла и флавонолов при сравнении средних значений двух независимых групп декоративных растений, о чем свидетельствуют данные, приведенные в таблице.
Таблица
Результаты выполнения t -теста для независимых выборок
|
Показатель, вид |
Mean |
t-value |
df |
p |
Valid N |
Std. dev. |
F-ratioVariances |
p Variances |
|
Травянистые растения |
||||||||
|
Chl И1–И2 |
59,44 |
2,34 |
8 |
0,04 |
10 |
2,92 |
12,82 |
0,03 |
|
48,1 |
10,45 |
|||||||
|
Flv И1– И2 |
1,49 |
3,32 |
8 |
0,01 |
10 |
0,10 |
3,66 |
0,24 |
|
1,32 |
0,05 |
|||||||
|
Flv И1– И3 |
1,49 |
-3,48 |
8 |
0,008 |
10 |
0,10 |
1,48 |
0,70 |
|
1,73 |
0,12 |
|||||||
|
Flv И2– И3 |
1,32 |
-6,97 |
8 |
0,0001 |
10 |
0,05 |
5,41 |
0,13 |
|
1,73 |
0,12 |
|||||||
|
Flv И2– И5 |
1,32 |
-6,09 |
8 |
0,0003 |
10 |
0,05 |
2,39 |
0,40 |
|
1,49 |
0,03 |
|||||||
|
Flv И3– И4 |
1,73 |
5,26 |
8 |
0,0008 |
10 |
0,12 |
4,09 |
0,20 |
|
1,41 |
0,06 |
|||||||
|
Flv И3– И5 |
1,73 |
4,31 |
8 |
0,003 |
10 |
0,12 |
12,94 |
0,03 |
|
1,49 |
0,03 |
|||||||
|
Древесные растения |
||||||||
|
Flv К1– К6 |
1,58 |
2,66 |
22 |
0,01 |
24 |
0,13 |
2,82 |
0,09 |
|
1,38 |
0,22 |
|||||||
|
Flv К7– К8 |
1,27 |
-3,88 |
38 |
0,0004 |
40 |
0,24 |
1,68 |
0,27 |
|
1,53 |
0,18 |
|||||||
|
Chl К2– К8 |
31,30 |
4,75 |
30 |
0,00005 |
32 |
5,20 |
3,04 |
0,03 |
|
24,46 |
2,99 |
|||||||
|
Chl К3– К8 |
28,88 |
4,86 |
30 |
0,00003 |
32 |
1,23 |
5,87 |
0,005 |
|
24,46 |
2,99 |
|||||||
|
Chl К5– К8 |
29,71 |
3,40 |
30 |
0,002 |
32 |
5,76 |
3,72 |
0,01 |
|
24,46 |
2,99 |
|||||||
|
Chl К2– К4 |
31,30 |
2,84 |
30 |
0,008 |
32 |
5,20 |
6,04 |
0,0007 |
|
27,60 |
2,12 |
|||||||
|
Chl К2– К6 |
31,30 |
2,96 |
22 |
0,007 |
24 |
5,20 |
1,73 |
0,38 |
|
23,94 |
6,84 |
|||||||
Примечание : Mean – средние значения для каждой группы; t - value – значение рассчитанного программой t -критерия Стьюдента; df – число степеней свободы; p – вероятность ошибки (критический уровень значимости принят равным р < 0,05); Valid N – объем выборки; Std. dev. – стандартное отклонение выборки; F-ratio, Variances – значение F -критерия Фишера, с помощью которого проверяется гипотеза о равенстве дисперсий в сравниваемых выборках; P, Variances – вероятность ошибки для F- теста Фишера.
При сравнении суммы рангов показателя Chl у двух независимых ненормально распределенных групп Acer pseudoplatanus и Acer mandshuricum выявлено наличие статистически значимой разницы между сравниваемыми выборками при P = 0,001 (рис. 3a). При анализе данных показателя Flv четырех видов: Acer rubrum, A. trautvetteri, A. tataricum, A. ukurunduense – и совокупностей показателя NBI пяти видов: Acer rubrum, A. trautvetteri, A. tataricum, A. mandshuricum, A. ginnala – наблюдаются различия между сравниваемыми группами, при P < 0,05 (рис. 3b, c).
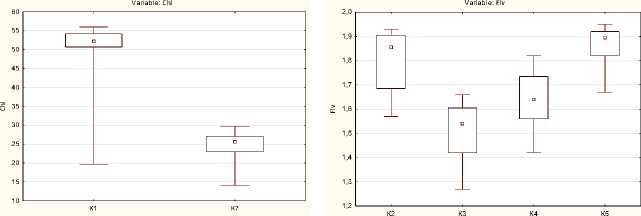
Вид
а
Вид b
* Примечание : Median – медиана; Perсentile – процентили 25–75%; min – минимальные значения показателя; max – максимальные значения показателя
Variable: Nbi
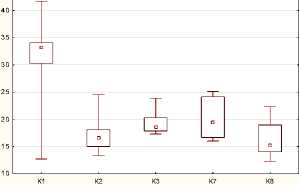
Вид
Рис. 3. Диаграмма сравнения нескольких независимых групп по одному показателю: a – Chl, b – Flv, c – Nbi
Выводы
Все изученные интродуценты находятся в удовлетворительном физиологическом состоянии, о чем говорит незначительное варьирование биохимических показателей в пределах изученных групп декоративных растений.
Максимальные значения показателей Nbi – 39,7; Chl – 58,6 мг/см2 (Iris hungarica ) и Nbi – 31,8; Chl – 49,9 мг/см2 ( Acer pseudoplatanus ) являются реакцией адаптации к условиям произрастания и указывают на формирование защитных функций растений.
Минимальное значение показателя Nbi – 15,9 у Acer ukurunduense является адаптационной реакцией к условиям достаточного освещения.
Исследование выполнено при финансовой поддержке Департамента образования и молодежной политики Ханты-Мансийского автономного округа – Югры (приказ № 1281 от 25.08.17 г.).
Список литературы Адаптационные возможности интродуцентов различного происхождения в условиях Ботанического сада г. Сургута
- Алексеева Н. Б. Род Iris L.(Iridaceae) в России // Turczaninowia. 2008. Т. 11. № 2. С. 5-70.
- Архангельская Г. П., Мухаев Б. А. Особенности адаптации ильмовых к почвенной засухе//Повышение устойчивости и долговечности защитных лесных насаждений. Волгоград, 1980. С. 72-81.
- Булыгин Н. Е. Дендрология. Л.: Агропромиздат, 1991. 352 с.
- Интродукция древесных растений в Сибири / науч. ред. И. Ю. Коропачинский. Новосибирск: Гео, 2017. 716 с.
- Кузнецов В. В., Дмитриева Г. А. Физиология растений. М.: Абрис, 2011. 783 с.
- Мамаев С. А., Дорофеева Л. М. Интродукция клена на Урале. Екатеринбург: УрО РАН, 2005. 104 с.
- Мамонтова Е. Н. Коллекция ирисов природной флоры, интродуцируемых в ботаническом саду Самарского госуниверситета // Самарская Лука, 2007. Т. 16. № 3(21). С. 518-531.
- Михеев А. Д. Iris L. // Конспект флоры Кавказа. СПб., 2006. Т. 2. С. 104-112.
- Павлова Н. С. Касатик - Iris L. // Флора Российского Дальнего Востока. Владивосток, 2006. С. 277-279.
- Панин С. И., Колесниченко Е. Ю, Морозова Т. С., Соловьева В. И. Оценка аккумуляции тяжёлых металлов древесными растениями полезащитной лесополосы и полевыми культурами // Вестник Московского государственного областного университета. Серия: Естественные науки. 2014. № 1. С. 75-80.
- Рязанова Н. А., Путенихин В. П. Клены в Башкирском Предуралье: биологические особенности при интродукции. Уфа: Гилем, 2012. 224 с.
- Соколов С. Я., Связева О. А. География древесных растений. М.-Л.: Наука, 1965. 266 с.
- Ткаченко К. Г. Виды рода Iris L. в коллекциях-экспозициях живых растений альпинария ботанического сада Петра Великого Ботанического института РАН // Вестник Удмуртского университета. Биология. Науки о Земле, 2013. Вып.3. С. 35-43.
- Фомина Т. И. Биологические особенности декоративных растений природной флоры в Западной Сибири. Новосибирск: Гео, 2012.179 с.
- Bueno J. M., Sáez-Plaza P., Ramos-Escudero F., Jiménez A. M., Fett R., Asuero A. G. Analysis and antioxidant capacity of anthocyanin pigments. Part II: chemical structure, color, and intake of anthocyanins // Critical Reviews in Analytical Chemistry. 2012. V. 42. № 2. P. 126-151.
- DOI: 10.1080/10408347.2011.632314
- Chen M., Blankenship R. E. Expanding the solar spectrum used by photosynthesis // Trends in plant science. 2011. V. 16. № 8. P. 427-431.
- DOI: 10.1016/j.tplants.2011.03.011
- Quemada M., Gabriel J. L., Zarco-Tejada P. Airborne hyperspectral images and ground-level optical sensors as assessment tools for maize nitrogen fertilization // Remote sensing. 2014. V. 6. № 4. P. 2940-2962.
- DOI: 10.3390/rs6042940
- Ritz T., Damjanović A., Schulten K., Zhang J. P., Koyama Y. Efficient light harvesting through carotenoids // Photosynthesis Research. 2000. V. 66. № 1-2. P. 125-144. :1010750332320
- DOI: 10.1023/A
