Адаптивные реакции надземной вегетативной сферы картофеля (Solanum tuberosum L.) сорта 'Башкирский' на густоту посадки
 сорта 'Башкирский' на густоту посадки")
Автор: Ишбирдин А.Р., Пур Д.А., Марданшин И.С.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 4, 2024 года.
Бесплатный доступ
Одним из подходов в оценке эффективности агротехнических приемов выращивания сельскохозяйственных культур является популяционный анализ с выявлением проявлений индивидуальных и популяционных стратегий жизни возделываемых растений и их сортовой специфичности. Исследования проводились в 2024 г. на базе Чишминского селекционного центра Башкирского НИИСХ. Объектом исследования был сорт ‘Башкирский'. Изучали морфологическую структуру надземной вегетативной сферы и ее изменчивость в вариантах с густотой посадки 17.8, 26.7 и 53.3 тыс. клубней/га. Изучили закономерности изменчивости 11 морфологических признаков: число побегов; высота растения (наиболее развитого побега); число листьев на наиболее развитом побеге; число пар крупных долей пластинки листа; длина и ширина листа, длина черешка наиболее развитого листа серединной формации; длина и ширина пластинки наиболее развитой доли листа, индексы листа и наиболее развитой доли листа. Однофакторный дисперсионный анализ выявил значимое влияние густоты посадки на большинство признаков, кроме индексов листа и боковой доли листа. Установили адаптивный характер изменчивости таких признаков, как число пар и размеры боковых долей пластинки листа, проявивших, соответственно, конвергентную и дивергентную онтогенетические тактики. Выявили структуру морфологической изменчивости с выделением групп индикаторных признаков. К генотипическим (таксономическим) индикаторам отнесены признаки с низкими показателями общей и согласованной изменчивости: высота растения, число листьев, длина листа, число пар долей листа, индекс доли листа, индекс листа. К биологическим индикаторам с низкой общей и высокой согласованной изменчивостью отнесены длина и ширина доли листа и ширина листа. Высокую общую и низкую согласованную изменчивость имеют экологические индикаторы, в наибольшей степени зависящие от внешних условий, - длина черешка и число побегов. Системные индикаторы не выявлены. Предстоит изучение влияния изменчивости надземной вегетативной сферы на структуру урожая картофеля.
Картофель, сорт ‘башкирский', густота посадки, изменчивость, онтогенетические тактики
Короткий адрес: https://sciup.org/147247239
IDR: 147247239 | УДК: 58.084.2 | DOI: 10.17072/1994-9952-2024-4-349-356
Adaptive responses of the aboveground vegetative sphere of potato (Solanum tuberosum L.) variety 'Bashkirsky' to planting density
One of the approaches to assessing the efficiency of agricultural practices in growing agricultural crops is population analysis with the identification of manifestations of individual and population life strategies of cultivated plants and their varietal specificity. The studies were conducted in 2024 at the Chishminsky Breeding Center of the Bashkir Research Institute of Agriculture. The object of the study was the ‘Bashkir' variety. We studied the morphological structure of the aboveground vegetative sphere and its variability in variants with a planting density of 17.8; 26.7 and 53.3 thousand tubers/ha. We studied the patterns of variability of 11 morphological traits: number of shoots; plant height (the most developed shoot); number of leaves on the most developed shoot; number of pairs of large lobes of the leaf blade, leaf length and width, petiole length of the most developed leaf of the middle formation; length and width of the blade of the most developed leaf lobe, indices of leaves and the most developed leaf lobe. One-way analysis of variance revealed a reliable effect of planting density on most traits, with the exception of leaf indices and the lateral leaf lobe. The adaptive nature of variability of such traits as the number of pairs and sizes of the lateral lobes of the leaf blade, demonstrating convergent and divergent ontogenetic tactics, respectively, was established. The structure of morphological variability with the allocation of groups of indicator traits was revealed. Traits with low rates of general and coordinated variability are classified as genotypic (taxonomic) indicators: plant height, number of leaves, leaf length, number of pairs of leaf lobes, leaf lobe index, leaf index. The length and width of a leaf lobe, leaf width are classified as biological indicators with low general and high coordinated variability. High general and low coordinated variability are characteristic of ecological indicators that depend most on external conditions - petiole length and number of shoots. No systemic indicators have been identified. The influence of variability in the aboveground vegetative sphere on the structure of potato yield remains to be studied.
Текст научной статьи Адаптивные реакции надземной вегетативной сферы картофеля (Solanum tuberosum L.) сорта 'Башкирский' на густоту посадки
Агротехника возделывания картофеля предполагает применение широкого спектра методов повышения урожайности и управления структурой урожая (товарность, производство семенного материала): предпосевная обработка почвы, сроки посадки, способы и глубина посадки, качество семенных клубней, предпосевная обработка клубней, густота посадки, применение средств защиты растений и удобрений, а также такие не совсем традиционные приемы, как предварительное проращивания клубней, декапитация растений, удаление завязавшихся плодов [Иванова, 2011; Васильев, 2012; Владимиров, Гайнутдинов, Аппаков, 2012; Табаков, Самаркина, Шашкаров, 2013; Митрофанов и др., 2016; Дыйканова, 2017 и мн. др.]. Практически все эти агротехнические приемы прямо или косвенно направлены на оптимизацию роста и развития надземной части растений, продления сроков и эффективности фотосинтетической активности, определяемых числом и высотой побегов, площадью ассимиляционной поверхности. На развитие надземных органов картофеля и структуру урожая влияют и постоянно действующие факторы (тип почвы и ее механический состав), а также климатические и погодные условия. В связи с этим большое значение приобретают многофакторные исследования [Васильев, Бобоев, 2012], среди которых и изучение индивидуальных и популяционных стратегий жизни картофеля и их сортовой специфичности [Ишбирдин, Кливаденко, Ишмуратова, 2011; Каримова, Аташ Пур, 2024].
Объекты и методы исследований
Исследования проводились в 2024 г. на базе Чишминского селекционного центра Башкирского НИИСХ. Почвы – чернозем оподзоленный, тяжелосуглинистого механического состава. Содержание гумуса в пахотном слое – 7.4%, рН – 6.9. Объектом исследования был сорт ‘Башкирский’, созданный селекционерами Башкирского НИИ СХ и Федерального исследовательского центра картофеля имени А.Г. Лорха скрещиванием сортов ‘Белоусовский’ и гибрида 289/82-3 ФИЦ картофеля. Сорт включен в Государственный реестр селекционных достижений в 2007 г. по Уральскому региону.
Посадка картофеля проводилась 23 мая под лопату в предварительно нарезанные гребни культиватором КОН-2,8. Гребни формировали с междурядьем 75 см, расстояние между растениями – 75, 50 и 25 см соответственно, густота посадки составляла 17.8, 26.7 и 53.3 тыс. клубней/га. Измерения проведены 17 июля. Погодные условия для формирования растений до этого срока были благоприятны как для закладки, так и для накопления урожая, характеризовались умеренно положительными температурами и достаточным уровнем осадков, характерными для данной агроклиматической зоны. Удобрения вносили методом разбрасывания перед окучиванием в дозе 800 кг/га (N16P16K16). Измеряли 9 признаков морфологической структуры надземных частей растения: число побегов; высоту растения (наиболее развитого побега); число листьев на наиболее развитом побеге; число пар крупных долей листа, длину и ширину листа, длину черешка наиболее развитого листа серединной формации; длину и ширину пластинки наиболее развитой доли листа. Для оценки формы рассчитывали индексы листа и наиболее развитой доли листа как отношение длины к ширине структурной единицы. Уровень общей изменчивости оценивали по шкале С.А. Мамаева [1970]; структуру изменчивости морфологических признаков выявляли по методике Н.С. Ростовой [2002]; оценку усредненного виталитета растений в варианте опыта, построение экоклина по благоприятности условий роста проводили по методикам А.Р. Ишбирдина и М.М. Ишмуратовой [2004]; онтогенетические тактики (характер изменения вариабельности признака на экоклине по направлению ухудшения условий роста) определяли по методике Ю.А. Злобина [1989] с изменениями [Методика…, 2020]. Для оценки значимости влияния факторов на формирование признака оценивалась достоверность различий средних по критерию Стьюдента и однофакторный дисперсионный анализ. Данные обрабатывались в программах Microsoft Office Excel 2007 и STATISTICA 12.
Результаты и их обсуждение
Фенотипическая изменчивость является общим свойством живых организмов меняться в определенных генотипом пределах (норма реакции). В отличающихся экологических условиях размах вариации реализованных отрезков в пределах нормы реакции может быть разным. Как правило, у растений увеличение размаха вариации со смещением в направлении увеличения изменчивости признака наблюдаются, во-первых, при неблагоприятных условиях и могут трактоваться как динамические проявления поиска жизнеспособной морфологической структуры организма (с последующим закреплением адаптивных форм с уменьшением уровня варьирования) или, во-вторых, как проявление разрушения целостности морфологической структуры в условиях крайнего стресса. Второе сопровождается снижением показателей жизненности организма – размеров, биомассы, способностей к воспроизводству.
Уровни общей изменчивости (коэффициент вариации (CV), %) признаков морфологической структуры надземных побегов картофеля сорта ‘Башкирский’ колеблются в широких пределах - от очень высокого до низкого. Очень высокий уровень изменчивости по вариантам опыта имеет признак длина черешка (40–48%); высокий – число побегов (28–29%); от среднего до высокого – индекс листа (13–24%); средний – ширина листа (14–18%), ширина доли листа (13–19%), длина доли листа (13–20%); от низкого до среднего – число пар долей (8–14%); низкий – индекс доли (12%), число листьев (8–11%), длина листа (10–12%), высота побега (10–12%). Признаки с очень низким уровнем изменчивости (менее 7%) не отмечены. В этом ряду можно предполагать снижение роли признака в адаптивном морфогенезе, однако это справедливо только в случаях одновременного снижения согласованной изменчивости (усредненного коэффициента детерминации признака с прочими признаками морфологической структуры – R2 ch ) с переходом признака в ряд генеотипических индикаторов. Индикационную роль признака в сложении морфологической структуры растения и изменение этой роли в условиях адаптации к стрессу можно оценить, анализируя структуру морфологической изменчивости организма, которая представляет координацию соотношений общей и согласованной изменчивости всех признаков морфологической структуры [Ростова, 2002].
На рисунке 1 представлена структура морфологической изменчивости надземных вегетативных органов картофеля. К генотипическим (таксономическим) индикаторам относятся признаки с низкими показателями общей и согласованной изменчивости: высота растения, число листьев, длина листа, число пар долей листа, индекс доли листа, индекс листа. Эти слабо взаимосвязанные в развитии признаки в наибольшей степени детерминируются генотипом и не играют большой роли в адаптации растений к неблагоприятным условиям. Низкий уровень общей изменчивости при высокой согласованной изменчивости характерен для биологических индикаторов, определяющих образ растения: длины и ширины доли листа и ширины листа. Эти признаки также не играют значительной роли в адаптивном морфогенезе растений. Высокую общую и низкую согласованную изменчивость имеют экологические индикаторы, в наибольшей степени среди всех признаков зависящие от внешних условий (длина черешка и число побегов). В большей степени такая индикаторная роль присуща признаку число побегов. Более широкий разброс показателей согласованной изменчивости отмечен для длины черешка с тенденцией перехода признака в ранг эколого-биологических (системных) индикаторов – с высокими показателями общей и согласованной изменчивости - в варианте 75x75 см. Причем, при наибольшей плотности посадки средняя длина черешка уменьшается в 1.5 раза и в 8.2 раза уменьшается показатель согласованной изменчивости, т.е. при увеличении плотности посадки отмечается автономизация в развитии признака. Для числа побегов с уменьшением площади питания и увеличением конкуренции за свет отмечается обратная тенденция – усиление роли признака как эколого-биологического индикатора, определяющего общее состояние системы. Среднее число побегов можно считать показателем благоприятности условий роста и развития растений картофеля сорта ‘Башкирский’ – с уменьшением корневой конкуренции за минеральное пита- ние и улучшением светового режима происходит увеличение среднего числа побегов на растение в 1,4 раза (2.4 – 2.9 – 3.3).
B11
C11
A11
> о
A1 B1 C1
B7
B8
C7 B6 B9
A9 A8A6
A4
B10 B2 A10 C10 B5 C8 A7
A3C2 C4 C5A2 A5
B3C3
0,00 0,02 0,04 0,06 0,08 0,10 0,12 0,14 0,16 0,18 0,20 0,22 0,24 0,26 0,28 0,30
R 2ch
Рис. 1. Структура морфологической изменчивости растений картофеля сорта ‘Башкирский’ при разной плотности посадки
По оси ординат – общая изменчивость признака (коэффициент вариации (CV, %), по оси абсцисс – согласованная изменчивость признака (усредненный коэффициент детерминации, R 2ch ); А - 75 x 75 см, В - 75 x 50 см, С - 75 x 25 см; признаки: 1 - число побегов, 2 - высота растения, 3 - число листьев на побеге, 4 - число пар долей листа, 5 – длина листа, 6 – ширина листа, 7 – индекс листа, 8 – длина доли листа, 9 – ширина доли листа, 10 – индекс доли листа, 11 – длина черешка
[The structure of morphological variability of the ‘Bashkir’ variety potato plants at different planting densities
On the ordinate axis – the total variability of the trait (coefficient of variation (CV, %), on the abscissa axis – the consistent variability of the trait (average coefficient of determination, R2ch); A - 75 x 75 cm, B - 75 x 50 cm, C - 75 x 25 cm; signs: 1 – the number of shoots, 2 – plant height, 3 – the number of leaves on the shoot, 4 – the number of pairs of leaf lobes, 5 – leaf length, 6 – leaf width, 7 – leaf index, 8 – leaf lobe length, 9 – leaf lobe width, 10 – leaf lobe index, 11 – petiole length]
Ранжирование признаков по силе влияния изучаемого фактора (доля факториальной дисперсии в общей дисперсии, η²) проводили по результатам однофакторного дисперсионного анализа. По показателю силы влияния густоты посадки на признаки морфологической структуры надземной части растения они составили следующий ряд: высота растения (31%) – длина черешка (20%) – число пар долей листа (17%) – число побегов (16%) – число листьев на побеге (14%) – ширина листа (11%) – длина листа (11%) – длина доли листа (7%). По результатам дисперсионного анализа влияние изучаемого фактора на ширину доли листа, индекс листа и индекс доли листа статистически не значимо.
Результаты попарного сравнения и статистической значимости различий признаков по вариантам приведены в таблице.
Характеризующие форму (вытянутость) листа и его структурных единиц (доли листа) расчетные показатели – индексы – значимо не различаются в вариантах с разной плотностью посадки. Все мерные и счетные признаки морфологической структуры имеют статистически значимые различия по вариантам опыта. Причем число побегов и высота растения значимо отличаются по всем парам вариантов, тогда как определяющие площадь листа признаки, такие как длина и ширина листа, число пар долей листа, длина и ширина доли листа не имеют статистически значимых различий между вариантами 75x75 см и 75x25 см. Число листьев и длина черешка не имеют статистически значимых различий между вариантами 75x50 см и 75x25 см, при этом число листьев в разреженных посадках меньше, а длина черешка больше, чем в загущенных вариантах опыта. Возможно, что эти признаки могут быть определяющими в сложении листовой мозаики и оптимизации использования света растениями.
Средние значения и изменчивость признаков и достоверность их различия по вариантам опыта [Average values and variability of traits and the reliability of their differences according to experimental variants]
Признаки морфологической структуры
Варианты и пары вариантов опыта
|
2- в |
Средние значения признака
|
75 |
3.3 |
42.8 |
11.6 |
4.3 |
23.0 |
16.3 |
1.4 |
7.4 |
4.2 |
1.8 |
1.2 |
|
50 |
2.9 |
38.4 |
12.5 |
4.0 |
21.7 |
14.5 |
1.6 |
6.7 |
3.9 |
1.8 |
0.8 |
|
25 |
2.4 |
45.7 |
12.6 |
4.5 |
23.9 |
16.4 |
1.5 |
7.4 |
4.2 |
1.8 |
0.8 |
|
Достоверность различия пар выборок по средним значениям признака |
|||||||||||
|
75 и 50 |
* |
*** |
*** |
*** |
* |
*** |
- |
** |
* |
- |
*** |
|
75 и 25 |
*** |
** |
*** |
- |
- |
- |
- |
- |
- |
- |
*** |
|
50 и 25 |
** |
*** |
- |
*** |
*** |
*** |
- |
** |
** |
- |
- |
|
Изменчивость признака. CV |
|||||||||||
|
75 |
28.9 |
10.6 |
10.5 |
14.0 |
10.8 |
15.7 |
12.6 |
15.6 |
15.9 |
12.1 |
40.0 |
|
50 |
29.1 |
12.1 |
7.8 |
8.1 |
12.8 |
18.5 |
23.6 |
20.0 |
18.7 |
12.2 |
47.6 |
|
25 |
27.9 |
9.8 |
8.7 |
11.3 |
9.7 |
14.4 |
17.5 |
12.9 |
13.3 |
12.1 |
39.6 |
|
Достоверность различия пар выборок по изменчивости признака |
|||||||||||
|
75 и 50 |
- |
- |
- |
*** |
- |
- |
*** |
- |
- |
- |
- |
|
75 и 25 |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
50 и 25 |
- |
- |
- |
* |
- |
- |
- |
** |
* |
- |
- |
|
Примечание. Различия статистически значимы для Р < 0.05 (*), Р < 0.01 (**) и Р < 0.001 (***). |
|||||||||||
Адаптивность различий размерных и счетных признаков растения на экологическом градиенте может быть оценена только при параллельном исследовании различий в изменчивости этих признаков. Так, например, при достоверном различии числа побегов и высоты растений между всеми вариантами опыта эти признаки не различаются значимо по изменчивости признака. Вероятно, что изменения этих признаков являются проявлениями, направленными на полноту использования ресурсов и не связанны с адаптивными механизмами морфогенеза, поскольку стабилизация любого признака является отражением результата, свершившейся (и на генетическом уровне контролируемой) адаптации с поддержанием уровня энтропии системы.
Рассмотрим для примера такой признак морфологической структуры листа, как число пар долей листа. При увеличении плотности посадки среднее число пар долей листа достоверно уменьшается с 4.3 до 4.0 шт. со статистически значимым уменьшением изменчивости признака. В этом проявляется адаптационное значение признака при умеренном загущении посадок. Дальнейшее загущение с увеличением конкуренции за свет и минеральное питание приводит к изменению роли признака морфологической структуры листа в поисках путей морфогенетической адаптации: происходит увеличение среднего значения признака (до 4.5 шт.) со статистически значимым увеличением изменчивости (дестабилизацией) признака (CV достоверно возрастает от 8.1 до 11.3%). Перераспределение усилий идет на увеличение размеров доли листа (средние размеры длины и ширины доли листа увеличиваются с 6.7 и 3.9 см до 7.4 и 4.2 см соответственно) и их стабилизацию (минимальные значения CV признаков в вариантах опыта – 12.9 и 13.3% соответственно). Таким образом, при возрастании конкуренции за свет и минеральное питание основная роль в адаптации переходит от признака число долей листа к размерам долей листа.
Для оценки характера изменения вариабельности признака по направлению усиления стресса (онтогенетические тактики) рассчитывали индекс виталитета растений (IVC) в вариантах опыта. Наибольшее значение виталитета растений - 1.06 - отмечено в варианте 75x75 см, в вариантах 75x50 см и 75x25 см, соответственно, 0.94 и 1.00. Таким образом, наименьший виталитет растений выявлен в варианте со средней нормой посадки. Более высокий виталитет растений в загущенной посадке складывается из-за более высоких, чем в варианте 75x50 см, средних значений параметров практически всех морфологических признаков кроме числа побегов.
Онтогенетические тактики формирования признаков морфологической структуры устанавливали по характеру изменчивости признака на градиенте благоприятствования условиям роста, устанавливаемом по значениям IVC. Как явно выраженные онтогенетические тактики рассматривали статистически значимые изменения уровня изменчивости признака в вариантах опыта. Таковыми проявили себя следующие признаки: число пар боковых долей листа, длина и ширина боковой доли листа. В изменениях числа пар боковых долей листа проявляется конвергентная онтогенетическая тактика (рис. 2). Это выражается в стабилизации признака с уменьшением его средних значений в направлении усиления стресса, приводящего к снижению продукционных процессов.
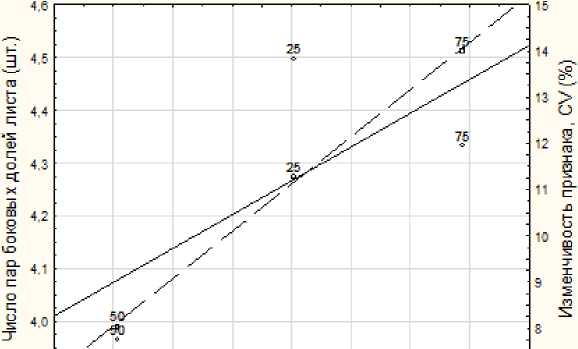

Рис. 2. Тренды конвергентной онтогенетической тактики признака число пар боковых долей листа картофеля сорта ‘Башкирский’.
Левая ось и сплошная линия – средние значения признака, правая ось и пунктирная линия – изменчивость признака
[Trends in the convergent ontogenetic tactics of the trait “number of pairs of lateral leaf lobes” of the ‘Bashkir’ potato variety.
Left axis and solid line are the average values of the trait; right axis and dotted line are the variability of the trait]
Дивергентную онтогенетическую тактику проявили параметры (длина, ширина) наиболее крупной центральной доли пластинки листа. На рис. 3 представлен характер изменения длины доли листа и изменчивости признака.
7,5
7,3
7,2
7,1
7,0
6,9
6,8
7,4
мс ,атсил илод анил
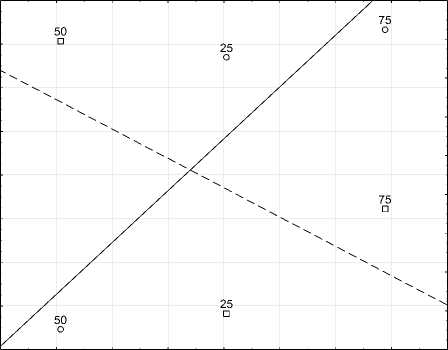
,аканзирп ьтсовичнемз
0,92 0,94 0,96 0,98 1,00 1,02
IVC
1,04 1,06 1,08
Рис. 3 Тренды конвергентной онтогенетической тактики признака длина доли листовой пластинки картофеля сорта ‘Башкирский’.
Левая ось и сплошная линия – средние значения признака, правая ось и пунктирная линия – изменчивость признака
6,7
[Trends in divergent ontogenetic tactics of the trait “length of the leaf blade lobe” of the potato variety ‘Bashkir’.
Left axis and solid line are the average values of the trait; right axis and dotted line are the variability of the trait]
Заключение
Анализ закономерностей изменчивости признаков морфологической структуры картофеля сорта ‘Башкирский’ на определяемом плотностью посадки экологическом градиенте показал их различную роль в адаптационном морфогенезе растений. Наиболее выраженные адаптивные морфогенетические реакции показали признаки, определяющие площадь фотосинтезирующей поверхности: число пар боковых долей листовой пластинки, длина наиболее развитой боковой доли листовой пластинки и ширина боковой доли листовой пластинки. Для этих признаков установлены значимые различия по вариантам опыта как их средних значений, так и их изменчивости.
Список литературы Адаптивные реакции надземной вегетативной сферы картофеля (Solanum tuberosum L.) сорта 'Башкирский' на густоту посадки
- Васильев А.А. Результаты многофакторных исследований по картофелю в условиях лесостепной зоны Южного Урала // Достижения науки и техники АПК. 2012. № 12. С. 32-35.
- Васильев А.А., Бобоев Д.А. Результаты многофакторных исследований по агротехнике картофеля // Селекция, семеноводство и технология плодово-ягодных культур и картофеля: сб. науч. тр. Челябинск: Челябинский дом печати, 2012. Т. 14. С. 246--267.
- Владимиров В.П., Гайнутдинов М.Т., Аппаков В.И. Агротехнические приемы повышения продуктивности картофеля на серой лесной почве лесостепи Среднего Поволжья // Вестник Казанского государственного аграрного университета. 2012. Т. 7, № 1. С. 119-124.
- Дыйканова М.Е. Декапитация в технологии возделывания раннего картофеля // Картофелеводство: материалы науч.-практ. конф. «Современные технологии производства, хранения и переработки картофеля». М., 2017. С. 161-164.
- Злобин Ю.А. Принципы и методы изучения ценотических популяций растений: учеб.-метод. пособие. Казань: Изд-во Казан. ун-та, 1989. 145 с.
- Иванова Н.В. Эффективность агротехнических приемов повышения урожайности и качества раннего картофеля в лесостепи Новосибирского Приобья // Вестник НГАУ (Новосибирский государственный аграрный университет). 2011. № 1. С. 36-41.
- Ишбирдин А.Р., Ишмуратова М.М. Адаптивный морфогенез и эколого-ценотические стратегии выживания травянистых растений // Методы популяционной биологии: сб. материалов VII Всерос. популяционного семинара. Сыктывкар, 2004. С. 113-120.
- Ишбирдин А.Р., Кливаденко Е.В., Ишмуратова М.М. Адаптивный морфогенез Solanum tuberosum // Известия Самарского научного центра РАН. 2011. Т. 13, № 5-2. С. 76-78.
- Каримова Л.Г., Аташ Пур Д. Популяционно-экологические реакции картофеля на условия выращивания // Актуальные вопросы современной науки: теория, технология, методология и практика: материалы XV междунар. науч.-практ. конф. Уфа, 2024. С. 20-24.
- Мамаев С.А. Закономерности внутривидовой изменчивости семейства Pinaceae на Урале: автореф. дис.... докт. биол. наук. Свердловск, 1970. 54 с.
- Методика изучения популяций редких и ресурсных видов растений на охраняемых природных территориях Республики Башкортостан / под ред. М.М. Ишмуратовой. Уфа: Башкирская энциклопедия, 2020. 275 с.
- Митрофанов Ю.И. и др. Влияние обработок картофеля от фитофторы на урожайность и экономическую эффективность его возделывания // Инновационные агро- и биотехнологии в адаптивно-ландшафтном земледелии на мелиоративных землях: материалы междунар. науч.-практ. конф. Тверь, 2016. С. 32-36.
- Ростова Н.С. Корреляции: структура и изменчивость. СПб.: Изд-во СПб. ун-та, 2002. 308 с.
- Табаков А.Г., Самаркина М.А., Шашкаров Л.Г. Урожайность картофеля в зависимости от агротехнических приемов возделывания // Вестник Казанского государственного аграрного университета. 2013. Т. 8, № 4. С. 143-145.
