Адаптивный морфогенез Solanum tuberosum

Автор: Ишбирдин Айрат Римович, Кливаденко Елена Владимировна, Ишмуратова Майя Мунировна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экосистемы
Статья в выпуске: 5-2 т.13, 2011 года.
Бесплатный доступ
Рассмотрены результаты исследования морфогенетических реакций картофеля сорта «Ароза» на ухудшение условий роста, создаваемых различной плотностью посадок в вариантах с удобрениями и без удобрений. Показано, что сорт имеет высокий защитный потенциал - выявлено проявление защитно-стрессовой онтогенетической стратегии.
Стратегии жизни, морфогенез, адаптация
Короткий адрес: https://sciup.org/148200410
IDR: 148200410 | УДК: 58.087
Adaptive morphogenesis of Solanum tuberosum
The results of research of morphogenetic reactions of a potato of a grade «Aroza» on stress are considered. Stressful conditions were created by increase in density of landing. Variants with application of fertilizers and without fertilizers were tested. It is shown that the grade has high protective potential. Ontogenetic strategy is revealed protectivelystressful.
Текст научной статьи Адаптивный морфогенез Solanum tuberosum
Оценка защитного потенциала сельскохозяйственных растений и их сортов является актуальной задачей, решение которой должно ускорить внедрение новых сортов и оптимизировать сортообо-рот при соблюдении принципов адаптивного земледелия. Авторским коллективом накоплен большой опыт оценки адаптивных морфогенетических реакций дикорастущих видов травянистых растений, преимущественно редких и ресурсных видов [6], а также пшеницы мягкой [1]. Исследуются проявления онтогенетических тактик и стратегии вида, как составляющих общей стратегии жизни.
МАТЕРИАЛ И МЕТОДЫ
Объектом исследования был картофель сорта «Ароза» (Германия). Агротехнологический паспорт характеризует сорт как отзывчивый на повышенные дозы минеральных удобрений и как засухоустойчивый, хорошо зарекомендовавший себя в южных регионах возделывания. Исследования проводили в 2010 г. на территории СПКК им. Салавата Стерлитамакского р-на Республики Башкортостан, были задействованы материалы, полученные на опытных участках и производственных полях хозяйства.
Год проведения исследований был экстремально засушливым и жарким: за весь период вегетации экспериментальных растений (1.05-8.08) выпало два дождливых дня в середине и конце мая (17 и 28 числа). Дневная температура длительный период держалась выше 25°С, поднимаясь до 35°С, относительная влажность воздуха падала до 6-9% (данные метеостанции г. Стерлитамак).
Полевые эксперименты включали 7 вариантов опыта: с плотностью посадок 30х30, 50х50 и 70х70 см (варианты с удобрением и без удобрений), а также 30х70 см (производственные посадки). Удобрения (нитрофоска) вносили в лунки в дозе 20 г. Для биоморфологических исследований отбирали по 30 модельных растений генеративного состояния. Для исключения краевого эффекта не учитывались растения периферических зон делянок.
Учет проводили 5-8 августа. У каждого растения измеряли по 14 признаков: число побегов, высота наиболее развитого побега (учетная единица при оценке следующих параметров), число листьев, число усохших листьев, длина листа серединной формации, длина непарной доли листа, ширина непарной доли листа, число пар развитых долей листа, число пар неразвитых долей листа, число пазушных побегов, общее число клубней, число товарных клубней (более 70 г), общая масса клубней, масса товарных клубней. Кроме того, учитывали отношения числа усохших листьев к общему числу листьев (интенсивность опада), числа пазушных побегов к общему числу листьев (вегета-тивность), числа товарных клубней к общему числу клубней (доля крупных клубней).
Для каждой группы (варианта опыта) модельных растений рассчитан общий коэффициент детерминации (R2 m ), как усредненный по всей матрице квадрат коэффициентов корреляции всех пар учитываемых параметров и коэффициенты детерминации отдельных учитываемых параметров (R2 ch ).
Как показатель жизненности в каждой из выборок рассчитан индекс виталитета растений [5]. Ин- декс рассчитывается с использованием выравнива- ния методом взвешивания средних:
L x / X •
IVC= Z1 _______ , где
N
X - среднее значение i-
того признака в ценопопуляции, X - среднее значение i-того признака для всех ценопопуляций, N -число признаков. Учитывая феномен "признакос-пецифичности" [3] для признаков, показавших тенденцию к увеличению с повышением неблагоприятности условий роста рассчитывалось обратное отношение: X / X .
Проводили регрессионный анализ зависимости коэффициента детерминации (R2m) признаков в каждой выборке (показатель морфологической целостности растений) от виталитета особей (IVC), а также корреляционный анализ связи средних зна- чений параметров и их вариабельности (коэффициент вариации, CV) с показателями виталитета особей.
Усиление морфологической интеграции растений расценивали как проявление защитной онтогенетической стратегии, а ее ослабление и дезинтеграцию морфологической структуры - стрессовой онтогенетической стратегии. По характеру онтогенетической стратегии оценивали адаптивность морфогенетических реакций в условиях стрессирующего воздействия. Наличие защитной стратегии или защитной составляющей в комбинированной стратегии оценивали как проявления устойчивости к стрессу на морфологическом уровне [4].
По характеру зависимости изменчивости параметра от виталитета особей выявляли онтогенетические тактики в формировании признаков морфологической структуры растения. Снижение вариабельности признака с уменьшением виталитета особей (ухудшением условий роста) оценивали как конвергентную онтогенетическую тактику, а увеличение – дивергентную онтогенетическую тактику [2].
Структуру изменчивости морфологической структуры оценивали по соотношению общей (CV) и согласованной (R2 ch ), изменчивости признака [7]. Индикаторная роль признака выявлялась по его положению в координационном поле структуры изменчивости.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
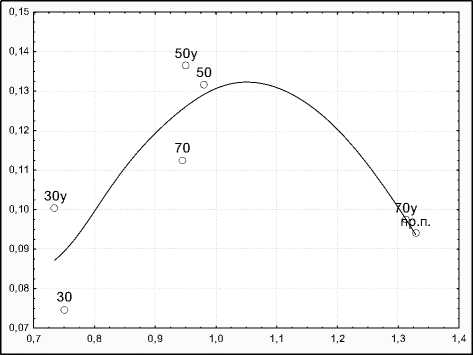
Максимальные значения виталитета растений отмечены в варианте 70х70 с удобрениями и в производственных посадках, причем как значения ви-талитета, так и морфологической целостности растений в этих вариантах были близки. Минимальный виталитет имели растения в варианте 30х30 без удобрения. Максимальной морфологической целостности растения достигли в вариантах 50х50.
Тренд онтогенетической стратегии представлен на рис. 1. Онтогенетическая стратегия сорта – защитно-стрессовая. Защитная составляющая проявилась в условиях нарастания стрессирующего воздействия, вызываемого конкуренцией растений за влагу и свет. В условиях крайне неблагоприятных (30х30 и, особенно, 30х30 без удобрений) наблюдали дезинтеграцию морфологической структуры, которая выразилась в дальнейшем уменьшении уровня средней детерминированности признаков морфологической структуры почти в два раза.
В формировании отдельных признаков морфологической структуры выявлены конвергентные, дивергентные и неопределенные тактики. Дивергентную тактику проявили все признаки структуры урожая: общее число клубней, число товарных клубней, общая масса клубней, масса товарных клубней, доля крупных клубней. Причем все эти признаки значимо (на уровне p <,05) коррелируют с жизненностью растений. Например, для массы то-
варных клубней это такие значения – -0,89 (вариабельность признака) и 0,99 (средние значения) .
Рис. 1. Зависимость морфологической интеграции (R2 m ) от виталитета (IVC) растений Solanum tuberosum .
Прим.: 30, 50, 70 – варианты с нормой посадки 30х30, 50х50 и 70х70 см, соответственно; у – варианты с удобрениями; пр.п. – производственные посадки.
Дивергентную тактику проявил и такой признак, как число пар неразвитых долей листа (корреляция с жизненностью растений вариабельности признака и его среднего значения, соответственно, r=-0,49 и r=0,63; статистически не значима). Все перечисленные признаки с ухудшением условий роста и при падении уровня морфологической целостности растений увеличивают свою вариабельность и не участвуют в адаптивном морфогенезе.
Конвергентную тактику проявил признак «длина листа», изменчивость которого положительно и статистически значимо коррелирует с жизненностью растений (r=0,92). Стабилизацию признака при морфологической дезинтеграции растений в целом можно считать адаптивной реакцией на стресс.
В структуре морфологической изменчивости растений (рис. 2) можно выделить две группы индикаторных признаков: генетические индикаторы (относительно низкие общая и согласованная изменчивость) и системные индикаторы (относительно высокие общая и согласованная изменчивость).
К системным индикаторам можно отнести все признаки структуры урожая, кроме числа клубней (генетический индикатор). Наиболее значимыми для оценки состояния системы являются число и масса товарных клубней.
Испытуемый сорт картофеля в условиях крайне неблагоприятного года и искусственно создаваемого стресса проявил свой адаптационный потенциал на уровне морфогенетических реакций.
Таким образом, разработанная для дикорастущих травянистых растений методика оценки морфогенетических реакций растений на стресс может с успехом применятся для оценки адаптивного потенциала сортов культурных растений.
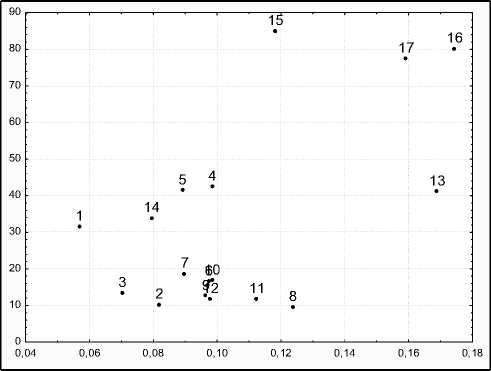
Рис. 2 . Структура изменчивости морфологических признаков Solanum tuberosum.
Прим. По оси абсцисс – согласованная изменчивость (R2 ch ), по оси ординат – общая изменчивость (CV). 1 – число побегов , 2 – высота побега, 3 – число листьев, 4 – число усохших листьев, 5 – интенсивность опада, 6 – число пазушных побегов, 7 – вегетативность, 8 – длина листа, 9 – число пар развитых долей , 10 – число пар неразвитых долей, 11 – длина непарной доли листа, 12 – ширина непарной доли листа, 13 – общая масса клубней, 14 – общее число клубней, 15 – доля крупных клубней, 16 – масса товарных клубней, 17 – число товарных клубней
Работа выполнялась при поддержке ФЦП «Исследования и разработки по приоритетным направлениям развития научно-технологического комплекса России на 2007-2012 годы», № 16.512.11.2014.
Список литературы Адаптивный морфогенез Solanum tuberosum
- Байзигитова Л.Н., Ишбирдин А.Р., Юмагужин Ф.Г. О некоторых проявлениях стратегии жизни мягкой пшеницы (Triticum aestivum) сорта Саратовская 55//Вестник Оренбургского гос. ун-та. 2009. № 6 (100). С. 54-56.
- Злобин Ю.А. Принципы и методы ценотических популяций растений. Казань: Изд. КазГУ, 1989. 146 с.
- Злобин Ю.А., Скляр В.Г., Мельник Т.И. Концепция континуума и градиентный анализ на уровне особей и популяций растений//Журнал общей биологии. 1996. Т. 57. № 6. С. 684-695.
- Ишбирдин А.Р., Ишмуратова М.М. Адаптивный морфогенез и эколого-ценотические стратегии выживания травянистых растений//Методы популяционной биологии: Материалы VII Всерос. популяц. семинара. Ч. 2. Сыктывкар, 2004а. С. 113-120.
- Ишбирдин А.Р., Ишмуратова М.М. К оценке виталитета ценопопуляций Rhodiola iremelica Boriss. по размерному спектру//Ученые записки НТГСПА: Материалы VI Всерос. популяц. семинара. Н. Тагил, 2004б. С. 80-85.
- Ишбирдин А.Р., Ишмуратова М.М. Некоторые направления и итоги исследований редких видов флоры Республики Башкортостан//Вестник Удмуртского ун-та. Серия 6: Биология. Науки о Земле. Вып. 1. 2009. С. 59-72.
- Ростова Н.С. Корреляции: структура и изменчивость. СПб., 2002. 308 с.
