Адаптивный потенциал фотосинтеза и продукционного процесса у местных форм и сортообразцов гречихи (Fagopyrum esculentum Moench) разных периодов селекции
 разных периодов селекции")
Автор: Амелин А.В., Фесенко А.Н., Чекалин Е.И., Заикин В.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Местные и селекционные сорта - факторы адаптации
Статья в выпуске: 1 т.51, 2016 года.
Бесплатный доступ
Современная селекция сельскохозяйственных культур, направленная главным образом на достижение максимальной продуктивности, существенно ослабляет защитные системы растений и, как следствие, снижает их устойчивость к неблагоприятным факторам внешней среды. Поэтому в последнее время особое внимание начинает уделяться созданию сортов на основе эволюционных принципов. В этой связи у сортообразцов гречихи ( Fagopyrum esculentum Moench) разных периодов селекции мы изучили норму реакции на изменение условий произрастания, отражающую способность растений адаптировать процессы фотосинтеза и формирования урожая к изменению условий произрастания для последующего использования этих данных в селекции культуры на повышение урожайности. В работе были использованы 13 сортообразцов гречихи, условно разделенных на три группы в зависимости от характера и срока селекции: местные (к-406, к-1709, коллекция ВИР, г. Санкт-Петербург), полученные селекционерами в 1930-1970-х годах (Калининская, Богатырь и Шатиловская 5) и современные сорта (Чатыр-Тау, Батыр, Девятка, Дизайн, Деметра, Дождик, Дикуль и Башкирская красностебельная). Впервые показано, что в результате селекции культуры не удалось добиться существенного повышения у растений устойчивости продукционного и фотосинтетического процессов, а также гомеостаза плодообразования. Современные сорта этой культуры проявляют высокую активность фотосинтеза и формируют высокую продуктивность лишь в благоприятных погодных условиях, но в стрессовых не имеют существенного преимущества перед своими предшественниками. В засушливом 2010 году они по семенной продуктивности достоверно не отличались от местных и старых сортов, тогда как в сравнительно благоприятные по температурному и водному режиму 2011-2013 годы превышение по анализируемому показателю составило в среднем 67,5 %, что в значительной степени было обусловлено реакцией фотосинтетического аппарата растений на условия произрастания. В засуху в фазу налива семян интенсивность фотосинтеза в листьях была снижена в среднем на 32,1 %, сухая масса надземных органов - на 46,7 %, а масса семян - на 67,5 %, по сравнению с показателями в благоприятные по температурному режиму и увлажнению 2011-2013 годы. С усилением дефицита влаги ситуация еще более ухудшается. При влажности почвы 30 % от полной влагоемкости у растений гречихи интенсивность фотосинтеза листьев снижалась в среднем в 4,4 раза, а продуктивность - на 41,8 %, по сравнению с показателями в варианте с оптимальным увлажнением. При этом более значимое уменьшение обоих показателей отмечали прежде всего у современных сортов: если у местного сортообразца к-1709 интенсивность фотосинтеза уменьшилась на 66,1 %, то у сортов Дикуль и Дождик - в среднем на 78,8 %. Таким образом, полученных данные указывают на низкий адаптивной потенциал современных сортов гречихи. В этой связи предлагается изменить принципы селекции культуры, направив ее на повышение гомеостаза плодообразования посредством отбора автогамных форм и создания самоопыляющихся сортов гречихи. Гибридизация с F. homotropicum позволяет существенно повысить инбредную жизнеспособность самоопыляющихся линий культурной гречихи, которые могут быть успешно использованы в селекционных программах по созданию автогамных сортов. Также больше внимания следует уделить повышению адаптивных возможностей самого продукционного процесса у растений, который недостаточно устойчив, чтобы обеспечить формирование высокого и стабильного урожая культуры. Перспективным представляется повышение активности и эффективности фотосинтеза, усиление начального роста, где скрыты огромные и пока что не реализованные резервы. Показан широкий диапазон наследственной изменчивости растений гречихи по интенсивность фотосинтеза в листьях (от 4,65 до 17,8 мкмоль СО2×м-2×с-1), позволяющий проводить по этому показателю целенаправленный отбор форм как методом гибридизации, так и массовым отбором в пределах отдельной популяции.
Культура, гречиха, селекция, сухая масса, интенсивность фотосинтеза и транспирации, адаптивность сорта
Короткий адрес: https://sciup.org/142133661
IDR: 142133661 | УДК: 633.12:631.52:631.524.85:581.132 | DOI: 10.15389/agrobiology.2016.1.79rus
Adaptiveness of productivity and photosynthesis in buckwheat (Fagopyrum esculentum Moench) landraces and varieties produced at different periods
Crop breeding if aimed mainly at the highest productivity results in a significant loss of defense system activity thus causing a decreased plant resistance to adverse environment factors. Reasonably, more attention is now being paid to evolution base in breeding. With this, we studied the norm of reaction to environment changes in buckwheat ( Fagopyrum esculentum Moench) cultivars and landraces as reflecting plant adaptation potential for photosynthesis and yield production to be further involved in breeding. A total of 11 buckwheat cultivars of which Kalininskaya, Bogatyr’ and Shatilovskaya 5 have been derived in 1930-1970, and Chatyr-Tay, Batyr, Devyatka, Dizain, Demetra, Dikul’ and Bashkirskaya krasnostebel’naya are the modern cultivars, together with landraces k-406 and k-1709 (VIR collection, St. Petersburg) were investigated. For the first time it was shown that in the course of buckwheat breeding no improvements in photosynthesis and production sustainability, as well as in homeostasis of grain formation have been achieved. Modern buckwheat cultivars possess high photosynthesis and productivity under favorable weather conditions, whereas at stresses do not have any significant advantage over their predecessors. In dry 2010 the seed production in modern cultivars was not reliably different from that in landraces and old cultivars, while in 2011-2013 at more favorable water supply and temperature it was on average 67.5 % higher, mostly due to the response of photosynthetic system to growing conditions. When drought occurred during the seed filling phase the photosynthetic activity in leaves decreased on average by 32.1 %, dry mass of the aboveground parts and seeds was lower by 46.7 % and 67.5 %, respectively, compared to those under favorable conditions in 2011-2013. With increasing water deficit the situation becomes worse. At soil moisture of 30 % of full capacity the activity of photosynthesis in buckwheat plant leaves was on average 4.4 times less, and seed production was 41.8 % less compared to the optimal moistening. At that, the losses were significantly higher in modern cultivars, e.g. in the k-1709 plants a 66.1 % decrease was found compared to 78.8 % in Dikul’ and Dozhdik plants. Thus the obtained data suggests a low adaptive potential of modern buckwheat varieties. So we propose to improve the seed formation homeostasis in buckwheat plants. In this regard, the selection of autogamous form and the creation of self-pollinating varieties can be used as more effective approaches. The hybridization with F. homotropicum can significantly improve the viability of self-pollinated inbred lines of buckwheat, which can be successfully used in breeding programs to create autogamous varieties. Moreover, the adaptiveness of the of yield formation processes which are not sustainable enough to guarantee the high and sustainable crop production must be improved. An increased activity and effectiveness of photosynthesis and initial growth seem to be of interest. It is shown that the leaf photosynthetic rate in buckwheat varies plants varies widely from 4.65 to 17.8 μmol CO2×m-2×s-1, which allows to select forms both by hybridizing and using selection within a population
Текст научной статьи Адаптивный потенциал фотосинтеза и продукционного процесса у местных форм и сортообразцов гречихи (Fagopyrum esculentum Moench) разных периодов селекции
За прошедшие 50 лет урожайность озимой пшеницы, ячменя, гре-
Эксперименты выполнены на научном оборудовании и приборах Центра коллективного пользования Орловского государственного аграрного университета «Генетические ресурсы растений и их использование».
чихи, сои, гороха, кормовых бобов и других сельскохозяйственных культур возросла в 2 раза и более (1). В этом определяющую роль, несомненно, сыграла селекция: во многих странах мира вклад сорта в обеспечение урожайности культур стал составлять свыше 50 % (2-5). При этом ощутимо ухудшилась устойчивость к биотическим и абиотическим факторам среды (6-9). По мнению ученых, последнее обстоятельство обусловлено тем, что земледелие в целом, и селекция в частности, направленные главным образом на достижение максимальной продуктивности, существенно ослабляют защитные системы растений и, как следствие, снижают их устойчивость к неблагоприятным условиям (10-13). Все острее проявляются различия в направлении действия естественного и искусственного отбора: если в стратегии эволюции определяющее свойство вида — способность выживать (адаптивность), то в селекции культурных растений к основным признакам длительное время относили технологичность сорта и высокую урожайность (экономическая эффективность производства).
Поэтому в последнее время все более актуальной становится смена приоритетов в селекции сельскохозяйственных культур, в частности ставится задача создавать адаптивные сорта на основе эволюционных принципов (10, 12, 14). Необходимость выведения для сельскохозяйственного производства адаптивных сортов определяется как контрастностью природно-климатических условий России, так и глобальными изменениями климата и возрастающей непредсказуемостью погоды (15-17).
Эта задача актуальна для гречихи, урожайность которой в России по-прежнему остается низкой (в среднем 0,75 т/га) ― прежде всего из-за недостаточной устойчивости современных сортов к экстремальным погодным факторам (18). Согласно данным Г.Е. Мартыненко с соавт. (19), у сортов гречихи величина средней урожайности отрицательно коррелирует с их экологической пластичностью ( r = - 0,737). Поэтому дальнейшее повышение урожайности культуры планируется обеспечить за счет реконструкции ее адаптивного генома (12), что требует проведения комплексных научных исследований.
В связи с проблемой сохранения и повышения адаптивного потенциала культуры нами впервые в вегетационных и полевых опытах при экстремальных изменениях метеоусловий исследована норма реакции продукционного процесса и фотосинтеза у сортообразцов гречихи разных периодов селекции.
Цель представляемой работы — выявить адаптивный потенциал фотосинтеза и продукционного процесса у растений гречихи и возможности его использования в селекции.
Методика . Объектами исследований, проведенных в 2010-2013 годах (Орловская обл.), были 13 сортообразцов гречихи ( Fagopyrum esculentum Moench), условно разделенных на три группы в зависимости от характера и периода селекции: местные (к-406, к-1709, коллекция ВИР, г. Санкт-Петербург), полученные селекционерами в 1930-1970-х годах (Калининская, Богатырь и Шатиловская 5) и современные сорта (Чатыр-Тау, Батыр, Девятка, Дизайн, Деметра, Дождик, Дикуль и Башкирская красностебельная).
Растения выращивали в селекционном севообороте (Всероссийский НИИ зернобобовых и крупяных культур). Преобладающий тип почв опытного участка — серые лесные среднесуглинистые. Посев проводили селекционной сеялкой СКС-6-10 (Россия) рядовым способом с нормой высева семян 3 млн шт/га. Площадь делянки составляла 10 м2, размещение рендомизированное, повторность 4-кратная. Уход за посевами и уборку выполняли в соответствии с методическими рекомендациями для ре-80
гиона. Вегетационный опыт по изучению засухоустойчивости сортов проводили в контролируемых тепличных условиях с использованием специальных вегетационных сосудов объемом 10 дм3. В опыте влажность почвы составляла 30 и 45 % от полной влагоемкости (ПВ), контролем служили растения, произраставшие при влажности почвы 70 % ПВ (повторность по каждому сорту 6-кратная).
Количество сухого вещества, накопленное листьями, стеблем, боковыми ветвями, соцветиями и семенами растений в разные фазы роста, учитывали при отборе проб (по 10 растений сорта с делянки, повторность 3-кратная) с последующим высушиванием образцов при 105 ° С в сушильном шкафу СМ 50/250 - 1000 ШС («СМ Климат», Россия). Уборочный индекс рассчитывали как отношение массы семян к общей сухой массе растения, выраженное в процентах.
Интенсивность фотосинтеза (ИФ) и транспирации (ИТ) определяли на интактных растениях в режиме реального времени с помощью портативного газоанализатора марки Li-COR-6400 («Li-COR Bioscience», США) согласно оригинальной методике в прилагаемой инструкции.
Статистическую обработку экспериментальных данных выполняли в программе Microsoft Excel.
Результаты . Погодные условия в период исследований оказались контрастными. В 2010 году во время вегетации отмечались высокие дневные температуры и выраженный дефицит осадков, тогда как в 2011 и 2013 годах метеорологическая обстановка была более благоприятной для гречихи как тепло- и влаголюбивой культуры.
1. Накопление сухой массы растений и эффективность ее использования на формирование семян у 13 изученных сортообразцов гречихи ( Fagopyrum esculentum Moench) по годам исследований (селекционный севооборот, Орловская обл.)
|
Показатель |
2010 |
2011 |
2012 |
2013 |
Среднее за 4 года |
|
Сухая масса надземных органов в |
|||||
|
период уборки, г/растение: |
|||||
|
среднее по сортам |
3,22 |
5,18 |
6,89 |
6,04 |
5,33 |
|
пределы варьирования |
2,51-4,18 |
3,91-6,21 |
6,47-6,88 |
4,65-7,10 |
4,57-5,90 |
|
Уборочный индекс, % |
|||||
|
среднее по сортам |
16,2 |
27,1 |
25,4 |
26,5 |
23,8 |
|
пределы варьирования |
10,4-24,7 |
18,7-36,0 |
20,9-32,4 |
14,1-46,5 |
19,1-30,7 |
П р и м еч а ни е. Описание сортообразцов (местные формы, современные и полученные в 1930-1970-е годы сорта) см. в разделе «Методика».
Проведенные исследования показали, что при умеренном увлажнении и оптимальной температуре воздуха (2011-2013 годы) растения гречихи способны накапливать в среднем 6,04 г, а в благоприятных метеоусловиях — до 7,10 г сухого вещества (табл. 1). Но в случае экстремальных условий 2010 года (высокая температура и дефицит влаги) общая продуктивность растений снижается более чем в 1,9 раза. При этом особенно резко падала эффективность использования ассимилятов на формирование семян. Так, величина уборочного индекса в 2010 году составляла всего 16,2 %, что было в 1,6 раза меньше, чем в более благополучные по увлажнению и температурному режиму 2011-2013 годы (см. табл. 1).
В результате семенная продуктивность растений в засушливых условиях 2010 года оказалась очень низкой — в среднем 0,52 г/растение, или 33 % от показателя за 2011-2013 годы (рис. 1). При этом масса семян уменьшалась более значительно, чем вегетативных органов. В условиях выраженного дефицита влаги и высокой температуры воздуха у растений гречихи семенная продуктивность была в 3,0 раза, а сухая масса вегета- тивных органов в 1,9 раза меньше, чем в более благоприятные по метеоусловиям годы. Варьирование семенной продуктивности у сортов гречихи находилось в пределах 0,33-0,87 г в 2010 году; 0,73-2,43 г — в 2011 году; 1,32-2,20 г — в 2012году и 0,77-2,58 г — в 2013 году.
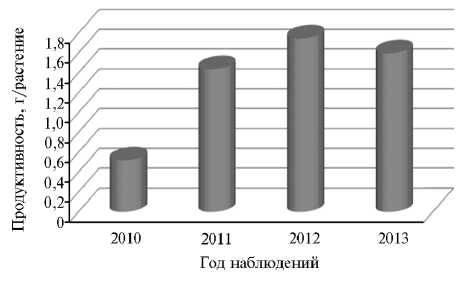
Рис. 1. Семенная продуктивность у растений гречихи ( Fagopyrum escu-lentum Moench) по годам исследований (среднее по 13 сортообразцам разных периодов селекции; селекционный севооборот, Орловская обл.). Описание сортообразцов (местные формы, современные и полученные в 1930-1970-е годы сорта) см. в разделе «Методика».
Наблюдаемые различия между надземными органами по накоплению сухого вещества в разные годы исследований могут быть обусловлены энтомофильным типом опыления растений и такой характерной для культуры особенностью, как способность поддерживать высокую интенсивность ве- гетативного роста на протяжении почти всей вегетации, в том числе в пе- риод репродуктивного развития, что негативно сказывается прежде всего на плодообразовании (12, 18).
По нашим данным, у современных сортов гречихи диапазон фенотипической изменчивости по числу плодов с растения был почти в 2 раза шире, чем по массе 1000 семян: по годам наблюдений первый показатель варьировал от 23,6 до 52,4 г, второй — от 27,2 до 28,5 г, что находилось в пределах ошибки опыта. Вследствие столь высокой зависимости плодо-образования от погодных условий у сортообразцов гречихи число семян, сформировавшихся на растении в засушливом 2010 году, оказалось в среднем на 51,5 % меньше, чем в 2011 и 2013 годах, тогда как их крупность почти не изменилась. Масса 1000 семян у 13 изученных сорто-образцов гречихи в 2010 году составляла в среднем 28,5±1,2 г, в 2011 году — 27,2±1,4 г, в 2013 году — 28,4±1,3 г. Иными словами, семенную продуктивность в значительной степени лимитировал процесс плодооб-разования. Одной из причин может быть низкая эффективность формирования фертильной пыльцы в экстремальных погодных условиях — характерная особенность гречихи как перекрестноопыляемой энтомофильной культуры (20).
Следует отметить, что стабильность плодообразования в процессе селекции гречихи практически не повышается (21), а ее широкое распространение (от субтропиков Южного Китая до северной границы земледелия) преимущественно обусловлено полиморфизмом популяций по продолжительности вегетации при слабом развитии физиологических адаптаций (22). В результате плодовитость вида обеспечивается длительным массовым цветением при очень низкой эффективности семяобразования: лишь около 10 % цветков завязывают семена (18, 23).
В связи с этим активно ведутся работы по повышению продуктивности семяобразования посредством отбора по указанному признаку в сортовых популяциях (24, 25). Считается, что такой отбор может быть наиболее результативен в гибридных популяциях с участием сортов гречихи из горных районов Индии и Юго-Восточной Азии, для которых характерны резкие колебания погодных условий (26). Еще одно селекционное направление ― выведение самоопыляющихся форм гречихи (27, 28). Предполагается, что получение автогамных сортов в первую очередь существенно снизит зависимость гречихи от опыления пчелами. Интерес к созданию автогамной гречихи резко возрос после обнаружения в горах на юге Китая дикого самоопылителя F. homotropicum Ohnishi, близкородственного гречихе обыкновенной F. esculentum Moench (29). В ряде лабораторий были проведены их успешные скрещивания и получены фертильные межвидовые гибриды, генетика которых сейчас детально изучается (30-33). Особый интерес к этому направлению исследований вызывает обнаруженная нами у дикого вида способность к адаптивному регулированию времени формирования семени, отсутствующая у культурной гречихи (34). Создать на основе этого материала самоопыляющийся сорт гречихи обыкновенной пока не удалось, что связано в основном с трудностями преодоления свойственной гречихе инбредной депрессии и слабой адаптированно-сти межвидовых гибридов к условиям умеренного климата (35). Тем не менее, это направление селекции весьма перспективно.
Повышая эффективность семяобразования у гречихи, по-види-мому, больше внимания следует уделить устойчивости самого продукционного процесса к неблагоприятным погодным факторам, так как у современных сортов она имеет выраженную тенденцию к снижению. В результате варьирование сухой массы надземных органов растений (в особенности семян) усиливается. В годы исследований диапазон генотипической изменчивости по массе семян с одного растения составлял у местных популяций ― 0,41-1,37 г; у старых сортов ― 0,61-1,76 г; у современных сортов ― 0,53-1,85 г. В засушливом 2010 году современные сорта по семенной продуктивности фактически не отличались от местных и старых сортов, тогда как в сравнительно благоприятные по температурному и водному режиму 2011-2013 годы превышение по анализируемому показателю у них составило в среднем 67,5 % (рис. 2).
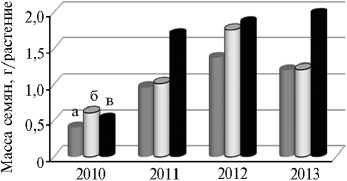
Год наблюдений
Рис. 2. Семенная продуктивность у сортооб-разцов гречихи ( Fagopyrum esculentum Moench) разной селекции по годам исследований : а ― местные сорта (к-406, к-1709, коллекция ВИР, г. Санкт-Петербург); б ― сорта Калининская, Богатырь и Шатиловская 5 (селекция 1930-1970-х годов); в ― современные сорта Чатыр-Тау, Батыр, Девятка, Дизайн, Деметра, Дождик, Дикуль и Башкирская красностебельная (селекционный севооборот, Орловская обл.).
Иными словами, современные сорта гречихи, как и многих других сельскохозяйственных культур, имеют выраженное преимущество перед своими предшественниками в основном в благоприятных условиях произрастания, но не в экстремальных. К возможным причинам этого можно также отнести слабое развитие корневой системы растений. По результатам исследований A. Lakhanov с соавт. (36), доля корней в общей биомассе растений у селекционных сортов гречихи формируется существенно меньше по сравнению с предковыми формами ( F . homotropicum и F . escu-lentum ssp. ancestrale ).
Низкая устойчивость продукционного процесса у современных сортов гречихи может быть обусловлена и высокой зависимостью фотосинтеза, происходящего в листьях растений, от внешних условий, что при их ухудшении приводит к резкому уменьшению урожайности (37). Известно, что продуктивность фотосинтеза сельскохозяйственных культур существенно зависит от таких факторов внешней среды, как температура, освещенность, влагообеспеченность, а также адаптивных особенностей вида и сорта (38-43). Об этом наглядно свидетельствуют и результаты проведенных нами исследований. Так, в 2010 засушливом году, когда на протяжении почти всего времени развития растений стояла сухая и жаркая погода, интенсивность фотосинтеза в листьях в фазу налива семян была снижена в среднем на 32,1 %, сухая масса надземных органов — на 46,7 %, а масса семян — на 67,5 % по сравнению с 2011-2013 годами (рис. 3). Пределы генотипического варьирования ИФ составляли в 2010 году — 4,65-10,80; в 2011 — 9,81-14,38; в 2012 — 14,74-17,8 и в 2013 — 7,92-12,9 мкмоль СО2 • м-2 • с-1.
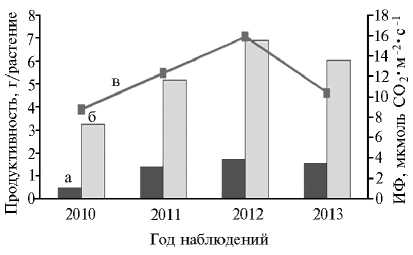
Рис. 3. Продуктивность и интенсивность фотосинтеза (ИФ) в листьях растений гречихи ( Fagopyrum esculentum Moench) у 13 изученных сортообразцов по годам исследований: а, б ― соответственно семенная продуктивность и общая надземная биомасса; в ― кривая ИФ (фаза цветения + 10 сут; селекционный севооборот, Орловская обл.).
При обострении засухи ситуация еще более усугубляется. В модельном вегетационном опыте было показано, что при влажности почвы 30 % от полной влагоемкости у растений гречихи интенсивность фотосинтеза листьев снижалась в среднем в 4,4 раза, а продуктивность — на 41,8 %, по сравнению с показателями в варианте с оптимальным увлажнением. При этом более значимое уменьшение обоих этих показателей отмечали прежде всего у современных сортов: если у местного сортообразца к-1709 интенсивность фотосинтеза уменьшилась на 66,1 %, то у сортов Дикуль и Дождик — в среднем на 78,8 % (табл. 2).
2. Интенсивность фотосинтеза (ИФ) и транспирации (ИТ) листьев у сорто-образцов гречихи ( Fagopyrum esculentum Moench) разных периодов селекции в зависимости от влажности почвы (вегетационный опыт, 2013 год, фаза цветения + 30 сут)
|
Сортообразец |
ИФ, мкмоль СО2 •м 2• с 1 |
ИТ, ммоль H 2 O •м 2^с 1 |
Температура листа, ° С |
|||
|
контроль |
опыт |
контроль |
опыт |
контроль |
опыт |
|
|
к-1709 (местная форма) Богатырь (сорт из |
6,22 |
2,11 |
2,80 |
0,96 |
20,2 |
21,3 |
|
группы полученных в 1930-1970-е годы) |
6,51 |
1,03 |
2,64 |
0,55 |
24,2 |
25,1 |
|
Дикуль (современный сорт) |
6,70 |
1,73 |
3,26 |
0,66 |
25,0 |
25,5 |
|
Дождик (современный сорт) |
8,23 |
1,37 |
2,81 |
0,65 |
22,2 |
23,7 |
|
Среднее |
6,92 |
1,56 |
2,88 |
0,71 |
22,9 |
23,9 |
|
НСР 05 |
1,69 |
1,47 |
1,12 |
0,52 |
0,23 |
0,22 |
|
Прим еч ани е. Контроль — 70 % от полной влагоемкости почвы |
(ПВ), опыт |
— 30 % от ПВ. |
||||
По нашему мнению, эти различия между генотипами могли быть обусловлены повышенной способностью растений местного сорта к транспирации, обеспечивающей в условиях недостатка влаги поддержание необходимого температурного режима листа и более активное поглощение корневой системой молекул воды из почвы. У всех изученных образцов гречихи в годы исследований отмечали достаточно высокую положительную корреляцию между интенсивностью фотосинтеза и транспирации ( r = 0,68; P0< 0,05).
В выполненных нами опытах активность газообмена в листьях рас- тений гречихи значительно варьировала и внутри сортовых популяций. Так, анализ 159 растений сорта Дикуль показал, что интенсивность фотосинтеза в листьях изменялась от 0,2 до 14,8 мкмоль СО2•м-2•с-1. При этом у 15 % обследованных растений отмечали интенсивность фотосинтеза от 12,0 до 14,5 мкмоль СО2•м-2•с-1, у 26 % — от 9,0 до 12,0, у 41 % — от 3,0 до 9,0 и у 18 % — от 0,2 до 3,0 мкмоль СО2•м-2•с-1. Полученные данные позволяют сделать вывод, что целенаправленную селекцию гречихи по интенсивности фотосинтеза можно проводить не только методом гибридизации, но и массовым отбором в популяции отдельного сорта, что позволит выровнять растения по способности обеспечивать себя ассимилятами и тем самым повысить урожайность популяции в целом.
Таким образом, анализ проблемы адаптивности сельскохозяйственных культур свидетельствует, что в результате селекции, направленной в основном на обеспечение максимальной продуктивности, устойчивость к неблагоприятным факторам внешней среды у современных сортов существенно снижена (10-13). Решением проблемы может быть создание самоопыляющихся сортов на основе отбора автогамных форм. Сложности, связанные с возникающей инбредной депрессии, которая сопровождает любые попытки получить автогамный материал на основе перекрестноопы-лителей, почти неизбежны, но, как показали наши эксперименты, преодолимы. Так, использовав гибридизацию с F. homotropicum , мы наблюдали существенное повышение жизнеспособности инбредных самоопыляющихся линий культурной гречихи (34, 35). Еще один подход предполагает повышение экологической пластичности и устойчивости самого продукционного процесса для обеспечения высокого и стабильного урожая. Здесь резервом остается селекция на повышении активности и эффективности фотосинтеза (44, 45).
Итак, впервые нами показано, что в результате селекции гречихи не произошло существенного повышения устойчивости продукционного процесса и фотосинтеза, а также стабильности и адаптивности плодообра-зования: в опытах интенсивность фотосинтеза и накопления сухого вещества растениями, а также эффективность использования ассимилятов на формирование семян в засушливых условиях были в среднем на 38,4 % ниже по сравнению с аналогичными показателями в благоприятные по влагообеспеченности и температурному режиму годы. Современные сорта проявили высокую активность фотосинтеза и формировали высокую продуктивность лишь в благоприятных условиях, но при стрессе не имели преимуществ по сравнению с сортами более ранних периодов селекции и местными формами. Выявлено, что у растений гречихи интенсивность фотосинтеза в листьях — признак с широким диапазоном генотипической изменчивости (от 4,65 до 17,80 мкмоль СО2•м - 2•с - 1), что позволяет проводить целенаправленную селекцию форм по этому показателю как с использованием гибридизации, так и при массовом отборе в пределах индивидуальной сортовой популяции.
Список литературы Адаптивный потенциал фотосинтеза и продукционного процесса у местных форм и сортообразцов гречихи (Fagopyrum esculentum Moench) разных периодов селекции
- Жученко А.А. Ресурсный потенциал производства зерна в России. М., 2004.
- Jain H.K. Eighty years of post Mendelian breeding for crop yield: nature of selection pressures and future potential. Indian J. Genet. Plant Breed., 1986, 1: 30-53.
- Richards R.A. Selectable traits to increase crop photosynthesis and yield of grain crops. J. Exp. Bot., 2000, 51: 447-458 ( ) DOI: 10.1093/jexbot/51.suppl_1.447
- Развитие инновационной деятельности в растениеводстве/Под ред. В.И. Нечаева. М., 2010.
- Сандухадзе Б.И., Кочетыгов Г.В., Рыбакова М.И., Бугрова В.В. Сортимент озимой мягкой пшеницы для Центрального региона России с повышенным потенциалом продуктивности и качества. Вестник ОрелГАУ, 2012, 3: 4-8.
- Молчан И.М., Ильина Л.Г., Кубарев П.И. Спорные вопросы в селекции растений. Селекция и семеноводство, 1996, 1-2: 36-51.
- Fernandez-Martinez J.M., Dominguez J., Perez-Vich B., Velasco L. Update on breeding for resistance to sunflower broomrape. Helia, 2008, 31: 73-84 ( ) DOI: 10.2298/HEL0848073F
- Fernandez-Escobar J., Rodriguez-Ojeda M.I., Flonso L.C. Distribution and dissemination of sunflower broomrape (Orobanche cumana Wallr.) race F in Southern Spain. In: Proc. 17th Int. Sunfl. Conf. Cordoba. Spain, 2008: 231-236.
- Амелин А.В. Морфофизиологические достоинства и недостатки современных сортов. Дальнейшие пути их совершенствования у зернобобовых и крупяных культур. Вестник ОрелГАУ, 2012, 3: 10-15.
- Шевелуха В.С. Эволюция агроэкотехнологий и стратегия адаптивной селекции растений. Вестник РАСХН, 1993, 4: 16-21.
- Амелин А.В. Биологический потенциал гороха и его реализация на разных этапах развития культуры. Селекция и семеноводство, 1999, 2: 15-21.
- Фесенко Н.В., Фесенко Н.Н., Романова О.И. и др. Теоретические основы селекции. Т. 5. Генофонд и селекция крупяных культур. Гречиха/Под ред. В.А. Драгавцева. СПб, 2006.
- Стрельников Е.А., Антонова Т.С. Новые экотипы Orobanche cumana Wallr. как следствие интенсификации возделывания подсолнечника. Мат. школы молодых ученых «Экологическая генетика культурных растений» (Краснодар, 24-26 сентября 2014 года). Краснодар, 2014: 122-133.
- Жученко А.А. Экологические и эволюционные подходы к адаптивной селекции растений. Мат. Всерос. науч.-произв. конф. «Интродукция нетрадиционных и редких сельскохозяйственных растений» (24-28 июня 1998 года). Пенза, 1998, т. 1: 7-25.
- Гончаренко А.А. Об экологической пластичности и стабильности урожайности сортов зерновых культур. Мат. Всерос. науч.-практ. конф. «Пути повышения устойчивости сельскохозяйственного производства в современных условиях» (13-15 июля 2005 года). Орел, 2005: 46-56.
- Грабовец А.И., Фоменко М.А. Создание и внедрение сортов пшеницы и тритикале с широкой экологической адаптацией. Зернобобовые и крупяные культуры, 2013, 6(2): 41-47.
- Парахин Н.В., Амелин А.В. Агробиологические индикаторы устойчивого развития растениеводства в Орловской области. Вестник ОрелГАУ, 2014, 5(50): 92-102.
- Daai Z., Jinfeng G., Yiping Q., Cui L., Qinqin L., Pengke W., Xiaoli G., Baili F., Pu Y., Yan C. Preliminary study on fecundity of common buckwheat under controlled conditions. Proc. 12th Int. Symp. on buckwheat (August 21-25, 2013). Slovenia, Laško, 2013: 172-174.
- Мартыненко Г.Е., Шипулин О.А., Фесенко А.Н., Бирюкова О.В. Сравнительная оценка урожайности и адаптивности современных сортов гречихи. В сб.: Новые сорта сельскохозяйственных культур -составная часть инновационных технологий в растениеводстве (научные материалы Шатиловских чтений, посвященных 115-летию Шатиловской СХОС, 12-13 июля 2011 года). Орел, 2011: 165-173.
- Zeller F.J. Buchweizen (Fagopyrum esculentum Moench): Nutzung, Genetik, Zuchtung. Bodenkultur, 2001, 3(52): 259-276.
- Фесенко А.Н., Бирюкова О.В., Шипулин О.А., Фесенко И.Н. Закономерности семяобразования у современных сортов гречихи различного морфотипа. Земледелие, 2014, 4: 43-45.
- Demidenko N., Logacheva M., Penin A. Comparison of abiotic stress response systems between Arabidopsis thaliana and Fagopyrum esculentum. Proc. 12th Int. Symp. on buckwheat (August 21-25, 2013). Slovenia, Laško, 2013: 72-73
- Kadirova L., Sitnykov A. Reproductive biology of buckwheat. Proc. 11th Int. Symp. on buckwheat (July 19-23, 2010). Orel, 2010: 331-339.
- Inoue N., Hagiwara M. Relationship between the harvest index and duration of each developmental stage of shoot apex in common buckwheat. Fagopyrum, 2000, 17: 51-56.
- Inoue N., Kumagai H., Hagiwara M. Improvement of fertilization rate by mass selection in common buckwheat. Fagopyrum, 2002, 19: 49-53.
- Naseem M., Dutta M., Shah S., Kumar P. Assessment of agro-morphological, physiological and genetic diversity among buckwheat cultivars. Proc. 11th Int. Symp. on buckwheat (July 19-23, 2010). Orel, 2010: 94-101.
- Campbell C.G. Present state and future prospects for buckwheat. Proc. 9th Int. Symp. on buckwheat (August 18-22, 2004). Czech Republic, Prague, 2004: 26-29.
- Woo S.H., Suzuki T., Mukasa Y., Morishita T., Yun Y.H., Park C.H. Present status, future breeding strategy and prospects for buckwheat. Proc. 12th Int. Symp. on buckwheat (August 21-25, 2013). Slovenia, Laško, 2013: 25-26.
- Ohnishi O. Distribution and classification of wild buckwheat species. 1. Cymosum group. Fagopyrum, 2010, 27: 1-8.
- Campbell C.G. Buckwheat crop improvement. Fagopyrum, 2003, 20: 1-6.
- Fesenko N.N., Fesenko I.N., Ohnishi O. Homostyly of two morphologically different lineages of Fagopyrum homotropicum Ohnishi is determined by locus S4, which is an S-locus related gene in the linkage group. Fagopyrum, 2006, 23: 11-15.
- Fesenko I.N., Fesenko A.N. Genetic basis of interspecific diversity of floral display size between cultivated outcrosser Fagopyrum esculentum Moench and wild selfer F. homotropicum Ohnishi. Fagopyrum, 2011, 28: 17-21.
- Woo S.H., Kim S.H., Tsai K.S., Chung K.Y., Jong S.K., Adachi Taiji, Choi J.S. Pollen-tube behavior and embryo development in interspecific crosses among the genus Fagopyrum. J. Plant Biol., 2008, 52: 302-310.
- Fesenko A.N., Fesenko I.N. Elements of genetic testing differences between Fagopyrum esculentum and F. homotropicum and some results of interspecific hybridization in selection of common buckwheat. Vestnik OrelGAU, 2013, 2(41): 2-5.
- Chrungoo N.K., Kreft I., Sangma S.C., Devadasan N., Dohtdong L., Chetri U. Genetic diversity in himalayan buckwheats: a perspective for use in crop improvement programmes. Proc. 12th Int. Symp. on buckwheat (August 21-25, 2013). Slovenia, Laško, 2013: 198-211.
- Lakhanov A., Napolova G., Napolov V., Kolomeychenko V. System of donor-acceptor relations between organs of buckwheat plants. Proc. 11th Int. Symp. on buckwheat (July 19-23, 2010). Orel, 2010: 241-245.
- Kasajima S., Itoh H. Effect of shading during different growth phases on yield parameters of common buckwheat cv. Kitawasesoba in the northern region of Japan. Fagopyrum, 2011, 28: 43-46.
- Thompson L. Weather variability, climatic change and grain production. Crop. Sci., 1975, 41: 535-541 ( ) DOI: 10.1126/science.188.4188.535
- Тарчевский И.А. Механизм влияния засухи на фотосинтетическое усвоение СО2. В сб.: Физиология фотосинтеза. М., 1982: 118-129.
- Kraft S.E., Dharmadhikari P. Variation in the relationship between corn yield and climate in a sample of counties in Illinois 1951-1980. Transactions of the Illinois State Academy of Science, 1984, 3-4: 219-228.
- Миракилов Х.М., Абдуллаев Х.А., Каримов Х.Х. Изучение интенсивности фотосинтеза у некоторых видов растений в связи с их эволюцией и селекцией новых сортов. Известия Академии наук Республики Таджикистан. Отделение биологических и медицинских наук, 2009, 1(166): 49-61.
- Monteith J.L., Moss C.J. Climate and efficiency of crop production in Britain. Philosophical Transactions of the Royal Society B: Biological Sciences, 1977, 281: 277-294 ( ) DOI: 10.1098/rstb.1977.0140
- Slattery R.A., Ort D.R. Photosynthetic energy conversion efficiency: setting a baseline for gauging future improvements in important food and biofuel crops. Plant Physiol., 2015, 168: 383-392 ( ) DOI: 10.1104/pp.15.00066
- Ничипорович А.А. Энергетическая эффективность фотосинтеза и продуктивность растений. Пущино, 1979.
- Zhu X.-G., Long S.P., Ort D.R. What is the maximum efficiency with which photosynthesis can convert solar energy into biomass? Curr. Opin. Biotechnol., 2008, 19: 1-7 ( ) DOI: 10.1016/j.copbio.2008.02.004
