Активация экспрессии генов, обеспечивающих иммунный ответ винограда, засухой, высокой температурой и их комбинацией

Автор: Сундырева М.А., Луцкий Е.О., Баранов М.О.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Агрономия
Статья в выпуске: 3, 2025 года.
Бесплатный доступ
Цель исследования - изучение влияния засухи, высокотемпературного стресса и их комбинации на транскрипционные реакции винограда различного происхождения, определяющие иммунный ответ. Исследование проведено на сортах винограда различного происхождения в моделируемых стрессовых условиях. Методы исследования общепринятые. Происхождение сорта винограда оказывало влияние на устойчивость к разным стрессовым факторам. Сорт Каберне-Совиньон в равной степени повреждался засухой, высокой температурой и их комбинированным действием (увеличение EL на 22 %). Сорта Молдова и Курчанский повреждались засухой (увеличение EL на 21 и 19 % соответственно), сорт Морозко повреждался засухой на 31 % и температурой на 45 %. Экспрессия генов «иммунного ответа» PR2, PR3, STS согласована с преобладающей устойчивостью сорта винограда к засухе или высокой температуре. При комбинировании засухи и высокой температуры уровень экспрессии данных генов снижался относительно воздействия отдельных стрессов, возрастала экспрессия генов NCED и MYC2, которые обусловливают ответную реакцию на абиотический стресс. Каждый сорт демонстрировал индивидуальные особенности приспособления к засухе, высокотемпературному стрессу и их комбинации. Установлено, что в условиях высоких температур и засухи защитные реакции против патогенов активно функционируют, а комбинированное действие двух факторов снижает уровень «иммунной» защиты сортов винограда евро-американского происхождения (Молдова, Кутузовский). Для сорта евро-амурского происхождения Морозко было характерно повышение экспрессии генов PR5 и PR10, связанных с иммунным ответом, при комбинированном действии абиотических стрессоров, что может обеспечивать усиленные защитные реакции.
Виноград, засуха, высокотемпературный стресс, комбинированный стресс, экспрессия генов, иммунный ответ, pr гены
Короткий адрес: https://sciup.org/140309929
IDR: 140309929 | УДК: 634.8.03+581.1+581.54 | DOI: 10.36718/1819-4036-2025-3-41-51
Drought, high temperature and their combinations activate the expression of genes that provide the immune response of grapes
The aim of the study is to investigate the effect of drought, high-temperature stress and their combination on the transcriptional reactions of grapes of different origins that determine the immune response. The study was conducted on grape varieties of different origins under simulated stress conditions. The methods of the study are generally accepted. The origin of the grape variety affected resistance to different stress factors. The Cabernet Sauvignon variety was equally damaged by drought, high temperature and their combined effect (EL increase by 22 %). The Moldova and Kurchansky varieties were damaged by drought (EL increase by 21 and 19 %, respectively), the Morozko variety was damaged by drought by 31 % and temperature by 45 %. Expression of the PR2, PR3, STS "immune response" genes is consistent with the predominant resistance of the grape variety to drought or high temperature. When drought and high temperature were combined, the expression level of these genes decreased relative to the effects of individual stresses, while the expression of the NCED and MYC2 genes, which determine the response to abiotic stress, increased. Each variety demonstrated individual features of adaptation to drought, high-temperature stress, and their combination. It was found that under conditions of high temperatures and drought, defense reactions against pathogens actively function, and the combined effect of the two factors reduces the level of "immune" protection of Euro-American grape varieties (Moldova, Kutuzovsky). The Morozko variety of Euro-Amur origin was characterized by an increase in the expression of the PR5 and PR10 genes associated with the immune response under the combined effect of abiotic stressors, which can provide enhanced defense reactions.
Текст научной статьи Активация экспрессии генов, обеспечивающих иммунный ответ винограда, засухой, высокой температурой и их комбинацией
Введение. Виноград – многолетняя сельскохозяйственная культура, имеющая большое экономическое значение в мире. Зона виноградарства России характеризуется умеренно континентальным климатом с тенденцией увеличения частоты стрессовых явлений [1]. Ограничение развития большинства фитопатогенов происходит в условиях дефицита осадков и температуры, превышающей 32–35 °С [2–4]. Засуха усиливает развитие бактериального патогена X. fastidiosa [5], однако снижает развитие возбудителя милдью Plasmopara viticola [6]. Летом наблюдается комбинированное действие засухи и жары, и ответ растения на один стрессовый фактор может как усиливать защитные реакции против другого воздействия, так и ослаблять их [7] из-за перекрытия реакций на отдельные стрессоры [8]. Стрессовые реакции являются затратными процессами, поэтому происходит «приоритизация» адаптации к тому или иному воздействию [9, 10]. В условиях высокой температуры развитие патогенов подавлено внешними условиями, и потребность в иммунных реакциях растения снижается. Засуха оказывает как положительное, так и отрица-
the financial support of the RSF grant, project № 23
тельное влияние на развитие патогенов в связи с особенностями физиологического ответа растения на водный дефицит [5, 6]. Абиотические воздействия могут оказывать ингибирующее и стимулирующее воздействие на «иммунный статус» винограда и, как следствие, влиять на последующее развитие фитопатогенов.
Цель исследования – изучение влияния засухи, высокотемпературного стресса и их комбинации на транскрипционные реакции винограда различного происхождения, определяющие иммунный ответ.
Объекты, материалы и методы . Объектами исследования были растения винограда сортов Каберне-Совиньон ( V. vinifera L.), Молдова (Гузаль кара × Виллар блан), Кутузовский (Молдавский × Датье де Сен-Валье), Курчанский (Мускат кубанский × Саперави северный), Мо-розко (Мицар × Саперави северный).
Эксперимент проводили на саженцах винограда 4-месячного возраста в 4-литровых горшках со смесью коммерческого грунта, торфа и вермикулита в соотношении 2 : 1 : 1. Экспериментальные условия создавали в климатической камере. Контрольные условия – (23 ± 0,5) °C
днем и (20 ± 0,5) °C ночью, полив ежедневный. Засуха – (23 ± 0,5) °C днем и (20 ± 0,5) °C ночью, полив отсутствовал в течение 3 недель. Высокотемпературный стресс поддерживали в течение 2 сут: с 6:00 до 11:00 – 30 °С, с 11:00 до 16:00 – 40 °С, с 16:00 до 20:00 – 30 °С, с 20:00 до 06:00 – 25 °С. При комбинированном стрессе растения 2 сут выдерживали в условиях высокой температуры на фоне засухи.
Повреждение тканей растений определяли по выходу электролитов (EL) из листовых дисков массой 0,1 г кондуктометрическим методом [11]. Развитие окислительного стресса оценивали в экстракте растительной ткани 0,5 % ТХУК по содержанию ТБК-активных веществ (TBARS) спектрофотометрическим методом [12]. Экспрессию генов определяли методом количественной ПЦР. Тотальная РНК выделена с помощью модифицированного CTAB-метода [13]. Синтез кДНК и ПЦР в реальном времени прове-
ден с использованием коммерческих наборов MMLV RT kit (Евроген) и qPCRmix-HS SYBR (Евроген) согласно рекомендациям производителя. Относительную экспрессию генов рассчитывали методом 2–ΔΔCt [14], референсный ген – фактор элонгации α. Последовательности праймеров генов приведены в таблице (F – прямой праймер, R – обратный праймер).
Все измерения проведены в 3-5 повторах, в 1 повторе 3 растения. Данные представлены как среднее ± стандартная ошибка. Тест Дункана был проведен для оценки достоверности различий между вариантами эксперимента при уровне значимости ниже 0,05. На рисунках достоверные различия представлены в виде строчных латинских букв. Статистические расчеты и анализ главных компонент выполнены в программе Statistica. Работа выполнена на базе ЦКП «Приборно-аналитический» ФГНУ СКФНЦСВВ.
Нуклеотидные последовательности праймеров Nucleotide sequences of primers
|
Ген |
F, 5’-3’ |
R, 5’-3’ |
|
1 |
2 |
3 |
|
Фактор элонгации α (EFα) |
GAA CTG GGT GCT TGA TAG GC |
ACC AAA ATA TCC GGA GTA AAA GA |
|
Стильбенсинтаза (STS) |
ATC GAA GAT CAC CCA CCT TG |
CTT AGC GGT TCG AAG GAC AG |
|
Халконсинтаза (СНS) |
GTT CTG GTC GTC TGC TCT G |
CCA GTC GCT GAT GCC TATC |
|
Фенилаланин-аммиаклиаза (PAL) |
TGA ACA ATG GCG AAA GTG AGA A |
TCT CTT GCG CTC TCA ACC TCT T |
|
Алленоксид циклаза (AOC) |
CTA CAC GGG AGA CCT GGA GAAG |
CTC CCT TCT TCC CTG GAA CAT T |
|
Алленоксид синтаза (AOS) |
CGG CGG AGA ATA AAC AGT GC |
GAC TTG AAA AGC ACA GCG GAT |
|
Синтетаза жасмоноил-L-аминокислот (JAR1) |
TGG GAT CAT GAC TGG CTC |
TGT CCT CCT CAG CAC TCC CCC |
|
Транскрипционный фактор MYC2 (MYC2) |
ACT GGG GTC TGG TCT TGT TAG C |
TGA GGC TGC TCC AAT GTC G |
|
Транскрипционный фактор NPR1 (NPR1) |
GAC CAC AAC CGA GCT TCT TGA TCT |
ATA ATC TTG GGC TCT TTC CGC ATT |
|
Рецептор АБК (PYL4) |
CCG TCG TCC AGC AAA TCG |
ACA TCT CCA TCG CCA ACA AC |
|
АБК-зависимая протеинкиназа (SnRK) |
ACC CAT CCC AAA GTG TAG AAG AG |
TCA TCA GCG TCA ATG TCA TCA AG |
|
9-цис-эпоксикаротиноид диоксигеназа (NCED) |
CCT CTG TCT CAC CGC AAT GG |
AGA GAG TGA TGG ACG GGC TG |
|
АБК альдегидоксидаза (AAO3) |
ATT GCT TCA GGG TTC ATC CG |
AGC TCG ATA CGT TTC AAA CA |
|
Белок теплового шока (HSP17) |
AGA AGA AGA GCC AGA AGA GAA G |
ACA CAC GAA GCG ACC AAG |
|
Осмотин (OSM) |
ACT GCA ACT TCG ATG CGT CA |
TGC GAA TTC GGC TAA GGT GT |
Окончание табл.
|
1 |
2 |
3 |
|
Патогенез-связанный белок 1 (PR1) |
CCC AGA ACT CTC CAC AGG A |
GCA GCT ACA GTG TCG TTC CA |
|
β-1,3-глюканаза (PR2) |
TGC TGT TTA CTC GGC ACT TG |
CTG GGG ATT TCC TGT TCT CA |
|
Хитиназа (PR3) |
TAT CCA TGT GTC TCC GGT CA |
TGA ATC CAA TGC TGT TTC CA |
|
Тауматин-подобный белок (PR5) |
TTC ACC CCC AGC TAT GCA G |
GCA TCG AAG TTG CAG TTG GT |
|
Рибонуклеаза-подобный белок (PR10) |
CGT TAA GGG CGG CAA AGA G |
GCA TCA GGG TGT GCC AAG A |
Electrolyte output (A) and MDA content (B) in grape leaves (Latin letters indicate significant statistical differences between the variants of stressors for a particular variety at p = 0.05)
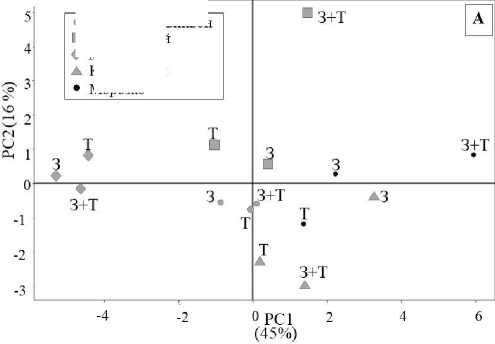
Согласно анализу главных компонент, разделение сортов винограда в условиях засухи, температурного стресса и при их комбинировании по экспрессии генов, участвующих в иммунных реакциях, было представлено главной компонентой 1 (PC1), и доля объясненной дисперсии составила 45 %. Наибольший вклад в PC1 вносят изменения экспрессии генов MYC2, NCED, PR1, SnRK, PYL4, STS, JAR1. Для сортов Кутузовский и Курчанский было выражено значительное смещение по главной компоненте 2 (PC2), где наибольший вклад вносили изменения различия в экспрессии генов CHS (46 %),
PR10 (37 %), PR3 (37 %), PR5 (29 %). Различия по главным компонентам у сортов Каберне-Совиньон и Молдова минимальны, а у сортов Морозко и Кутузовский наблюдались существенные изменения под воздействием стрессоров (рис. 2).
Гены NCED, SnRK, PYL4 обеспечивают синтез и рецепцию абсцизовой кислоты (АБК), JAR1, MYC2 – синтез и сигналинг жасмоновой кислоты, PR1, 2, 3, 5, 10 – патогенез-связанные белки, обеспечивающие подавление роста и развития патогенов, гены STS, CHS и PAL – гены фенольного обмена растений.
• Каберне-Совиньон
□ Кутузовский
♦ Молдова Курчанский Морозко
Рис. 2. Анализ главных компонент сортов винограда после воздействия засухи, высокой температуры и их сочетания (А): РС1 – главная компонента 1; PC2 – главная компонента 2; З – засуха; Т – температурный стресс; З + Т – комбинированное действие засухи и температурного стресса (коэффициент объясненной дисперсии PC1 = 45 %, коэффициент объясненной дисперсии PC2 = 16 %; вклад относительной экспрессии 19 генов в главную компоненту 1 (Б))
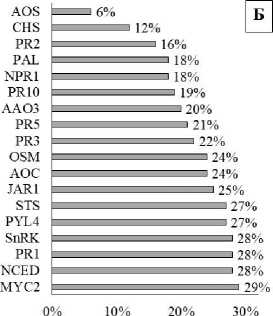
Analysis of the main components of grape varieties after exposure to drought, high temperature and their combination (A): PC1 – main component 1; PC2 – main component 2; H – drought; T – temperature stress; H + T – combined effect of drought and temperature stress (coefficient of explained dispersion PC1 = 45 %, coefficient of explained PC2 variance = 16 %; contribution of the relative expression of 19 genes to the main component 1 (Б))
Экспрессия генов «иммунитета» значительно отличалась между сортами винограда как по относительному уровню, так и по наличию или отсутствию разницы при воздействии разных стрессоров. Экспрессия большинства генов существенно возрастала при воздействии стрессоров, за исключением экспрессии генов PYL4 и PR10, когда изменения были либо минимальными, либо наблюдалось снижение их экспрессии (рис. 3).
Типы стрессоров инициировали различия в уровне экспрессии генов у винограда. У сорта Каберне-Совиньон наблюдалась разница в экспрессии 3 генов при разных стрессовых усло-
виях, у сортов Молдова и Кутузовский – 8 генов, у Курчанского стрессовые воздействия по-разному изменяли уровень экспрессии 5 генов, а у сорта Морозко – 13 генов. В условиях комбинированного стресса проявлялись различия в экспрессии гена NCED, а у сортов межвидового происхождения и MYC2. Данные различия были согласованы со значительным увеличением содержания TBARS, характеризующим развитие вторичного окислительного стресса. АБК – ведущий регулятор ответных реакций на засуху и тепловой стресс, и ключевым ферментом, определяющим биосинтез АБК, является 9-цис-эпокси-каротиноиддиоксигеназа (NCED) [21]. При зара-
Результаты и их обсуждение. Восприимчивость сортов винограда к воздействию стрессоров определяли по уровню повреждения тканей на основе выхода электролитов (EL) и степени перекисного окисления липидов на основе содержания ТБК-активных соединений (TBARS). Для сорта Каберне-Совиньон был характерен выраженный рост EL при всех типах воздействия, при этом разница между ними отсутствовала. Сорта Молдова и Курчанский повреждались засухой, сорт Морозко больше повреждался высокой температурой. Такие особенности могут объясняться наследуемыми от видов рода Vitis североамериканского происхождения высокой засухо- и жаростойкостью для Молдовы и низкой жаростойкостью V. amurensis для сорта Морозко [15, 16]. EL в листьях сортов Молдова, Курчанский и Морозко был ниже при комбини-
рованном действии засухи и высокой температуры, чем при действии наиболее повреждающего стрессора (рис. 1, А), что может объясняться закаливанием растений засухой перед температурным воздействием [17].
Засуха не влияла на содержание TBARS у сортов Каберне-Совиньон и Морозко, провоцировала рост окислительных процессов у Молдовы и Курчанского. При высокотемпературном воздействии TBARS было ниже либо равно таковому у винограда на фоне засухи, а при комбинированном стрессе наблюдалось выраженное увеличение содержания TBARS, что свидетельствует о значительной генерации АФК (рис. 1, Б), которая может быть обусловлена ингибированием антиоксидантных ферментов избытком Н 2 О 2 [18, 19] и превышением способности растения к адаптации [20].
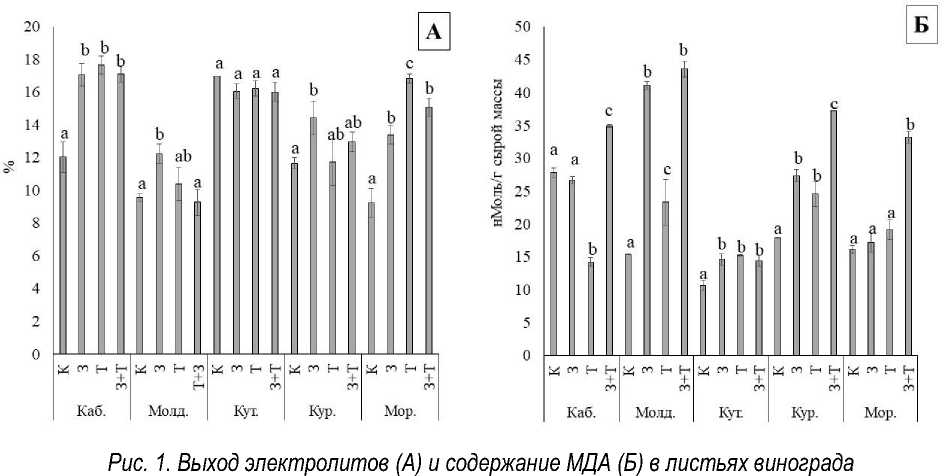
(латинскими буквами обозначены значимые статистические различия между вариантами стрессоров по отдельному сорту при p = 0,05)
жении винограда оидиумом наблюдается выраженное увеличение экспрессии генов, связанных в том числе с синтезом и сигналингом АБК [22], а экзогенная обработка АБК повышала устойчивость винограда к милдью, что обусловлено взаимодействием регуляторных путей жасмоната и АБК посредством PYL4 [23]. В то же время в зависимости от комбинации внешних условий АБК может подавлять экспрессию генов, связанных с синтезом и сигналингом салициловой, жасмоновой кислот и этилена – ключевых регуляторов «иммунной реакции» растений [22, 24], и содержание АБК значимо выше у чувствительного к милдью сорта винограда в сравнении с устойчивым сортом [25]. Фактор транскрипции MYC2 является основным регулятором сигнальной ветви жасмоновой кислоты, участвующей в ответных реакциях на множественные стрессовые факторы, главным образом биотические. MYC2 индуцирует экспрессию генов, обеспечивающих синтез и трансформацию жасмоновой кислоты, а промежуточное соединение в цепи образования жасмоновой кислоты – 12-оксофи-тодиеновая кислота – усиливает закрывание устьиц и повышает засухоустойчивость [26]. Ком-
Вестник КрасГАУ. 2025. № 3 (216) бинированное действие засухи и жары у всех изучаемых сортов винограда оказывает существенную нагрузку на растения, что проявляется в защитной реакции, регулируемой АБК. Для сортов межвидового происхождения было характерно усиление защитных реакций посредством жасмонат-зависимых процессов.
Под воздействием засухи наблюдали значимо больший уровень экспрессии генов PR1, PR2, PR3, PAL, STS, CHS у Морозко, что было согласовано с меньшей восприимчивостью сорта к дефициту воды. Менее восприимчивый к температурным воздействиям сорт Молдова демонстрировал более выраженное увеличение экспрессии генов PR2, PR3, PR5, STS, CHS в условиях высокой температуры. У сорта Кутузовский, который практически не повреждался засухой и высокой температурой в сравнении с контролем, одинаково высоким был уровень экспрессии PR2, PR3 и STS. Для сорта евро-амурского происхождения Морозко характерно повышение экспрессии генов PR5 и PR10, связанных с иммунным ответом, при комбинированном действии абиотических стрессоров.
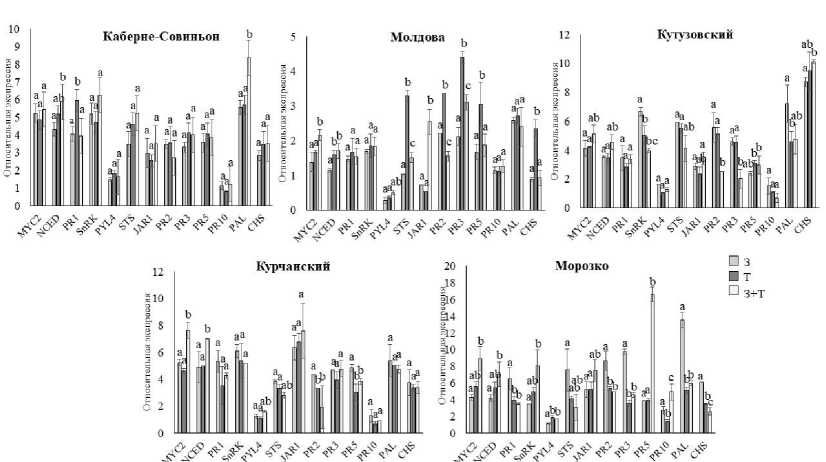
Рис. 3. Относительная экспрессия генов, участвующих в ответных реакциях на биотический стресс, в условиях засухи, высокотемпературного стресса и их комбинации: З – засуха;
Т – температурный стресс; З + Т – комбинированное действие засухи и температурного стресса (латинскими буквами обозначены значимые статистические различия экспрессии генов винограда между воздействием разных типов стрессоров при p = 0,05)
Relative expression of genes involved in responses to biotic stress in conditions of drought, high-temperature stress and their combinations: З – drought; T – temperature stress; З + T – combined effect of drought and temperature stress (Latin letters indicate significant statistical differences in gene expression of grapes between the effects of different types of stressors at P = 0.05)
Стильбенсинтазы (STS) осуществляют биосинтез стильбенов – фенольных соединений, обладающих множественными биологическими функциями. Наиболее изученным является влияние стильбенов на устойчивость винограда к заражению фитопатогенами [27], однако велика их роль в устойчивости к абиотическим стрессам, обусловленная участием в контроле АФК [28]. Было показано, что у трансгенных растений табака с высокой устойчивостью к патогенам и высокотемпературному стрессу существенно возрастала экспрессия PR3, обусловленная более интенсивной генерацией АФК [29]. В условиях биотического стресса PR2 и PR3 экспрессируются в ответ на сигналы жасмоновой кислоты [30], которая связана в том числе с ответными реакциями на засуху. Было показано, что более высокое содержание стильбеновых фитоалексинов и повышенная экспрессия PR2 и хитиназ были характерны для сорта с высокой устойчивостью к засухе и Botrytis cinerea [31, 32].
Заключение. Происхождение сорта винограда оказывало влияние на устойчивость к разным стрессовых факторам. Сорт Каберне-Совиньон в равной степени повреждался засухой, высокой температурой и их комбинирован-
ным действием. Сорта Молдова и Курчанский повреждались засухой, сорт Морозко – засухой на 31 % и температурой на 45 %. Экспрессия генов «иммунного ответа» PR2, PR3, STS согласована с преобладающей устойчивостью сорта винограда к засухе или высокой температуре. При комбинировании засухи и высокой температуры уровень экспрессии данных генов снижался относительно воздействия отдельных стрессов, и возрастала экспрессия генов NCED и MYC2, которые обусловливают ответную реакцию на абиотический стресс. Каждый сорт демонстрировал индивидуальные особенности приспособления к засухе, высокотемпературному стрессу и их комбинации. В условиях высоких температур и засухи защитные реакции против патогенов активно функционируют, а комбинированное действие двух факторов снижает уровень «иммунной» защиты сортов винограда евро-американского происхождения (Молдова, Кутузовский). Для сорта евро-амурского происхождения Морозко характерно повышение экспрессии генов PR5 и PR10, связанных с иммунным ответом, при комбинированном действии абиотических стрессоров, что может обеспечивать усиленные защитные реакции.
