Активность ферментов метаболизма активных форм кислорода и уровень световой эмиссии мицелия гриба Neonothopanus nambi при действии салициловой кислоты

Автор: Тюлькова Н.А., Бондарь В.С.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Биологические науки
Статья в выпуске: 2, 2017 года.
Бесплатный доступ
Проведена оценка активности ферментов метаболизма активных форм кислорода (АФК) - НАДФН-оксидазы, каталазы, супероксиддис-мутазы и пероксидазы и интенсивности све-товой эмиссии механически поврежденного мицелия светящегося гриба Neonothopanus nambi без и при добавках экзогенной салицило-вой кислоты (СК). Показано, что в условиях острого эксперимента, после повреждения образцов мицелия и последующей их инкубации в питательной среде, в течение короткого (6-7 часов) интервала времени наблюдается значительное (на 2 порядка и более) увеличе-ние уровня световой эмиссии гриба. Повыше-ние свечения регистрируется на фоне суще-ственного снижения в мицелии активности супероксиддисмутазы (СОД) и общей перокси-дазной активности. При этом уровень актив-ности НАДФН-оксидазы в грибе практически не меняется в течение всего эксперимента, а активность каталазы незначительно возрас-тает. Установлено, что добавка СК в среду инкубации заметным образом подавляет све-товую эмиссию мицелия N. nambi. При этом выявлено, что экзогенная СК активирует все изучаемые ферменты метаболизма АФК за исключением каталазы, активность которой несущественно снижается. Установлено, что эффекты СК являются дозозависимыми и на-растают с увеличением концентрации реа-гента. Совокупность полученных данных со-гласуется с высказанной нами ранее гипоте-зой, что грибное свечение является дополни-тельным механизмом антиоксидантной за-щиты от повреждающего действия АФК. На-блюдаемое в работе значительное увеличение световой эмиссии поврежденного мицелия при снижении активности СОД, особенно общей пероксидазной активности, может свиде-тельствовать о запуске в грибе этого меха-низма защиты от АФК, прежде всего от Н2О2 и иных пероксидных соединений.
Активные формы кисло-рода, светящиеся грибы, салициловая кисло-та, надфн-оксидаза, каталаза, супероксид-дисмутаза, пероксидаза
Короткий адрес: https://sciup.org/14084845
IDR: 14084845 | УДК: 577.334
Activity of ros metabolism enzymes and light emission level of Neonothopanus nambi myccelium under the action of salicylic acid
The metabolism enzymes activity of reactive ox-ygen species (ROS) - NADPH-oxidase, catalase, superoxide dismutase and peroxidase and the in-tensity of light emission in the mechanically dam-aged mycelium of the luminous fungus Neonothopanus nambi without and with addition of salicylic acid (SA) were studied. It was shown that in the conditions of acute experiment, after the me-chanical damage of mycelium samples and its sub-sequent incubation in a nutrient medium for a short time interval (6-7 hours), there was a significant (2 orders of magnitude or more) increase in the light emission of the fungus. A luminescence in-crease was registered against the background of a significant decrease of the superoxide dismutase (SOD) activity and total peroxidase activity in the mycelium. At the same time, the activity level of NADPH-oxidase in the fungus remained virtually unchanged throughout the experiment, but the ac-tivity level of catalase increased slightly. The addi-tion of SA in the incubation medium considerably inhibited the light emission of N. nambi mycelium. Wherein it was shown that the exogenous SA acti-vated all the studied ROS metabolism enzymes, except for catalase activity, which insignificantly decreased. The data obtained is consistent with our earlier hypothesis that the light emission of fungi is an additional mechanism of antioxidant protection against damaging effects of ROS. Significant in-crease in the light emission of the damaged myce-lium with a decrease in SOD activity was observed and especially, total peroxidase activity may indi-cate the start of this mechanism in the fungus for the protection against ROS, first of all H2O2 and other peroxide compounds.
Текст научной статьи Активность ферментов метаболизма активных форм кислорода и уровень световой эмиссии мицелия гриба Neonothopanus nambi при действии салициловой кислоты
Введение. Исследования светящихся высших грибов Neonothopanus nambi и Armillaria borealis позволили нам развить идею об участии активных форм кислорода (АФК) в механизме грибной люминесценции [1–7]. В этих работах было показано, что световая эмиссия грибного мицелия значительно возрастает при инкубации в деионизованной воде, механическом повреждении, радиационном облучении. Было установлено, что воздействие стрессовых факторов на гриб сопровождается не только увеличением интенсивности его люминесценции, но и активацией образования АФК и повышением общей пероксидазной и каталазной активностей [5, 7– 9]. Совокупность полученных данных позволила высказать гипотезу, что свечение грибов явля- ется дополнительным механизмом антиоксидантной защиты, нейтрализующим повреждающее действие АФК [5, 7–9].
Известно, что активация образования АФК является универсальной неспецифической ре-а к ц ией клетки на воздействие стрессовых факторов разной природы (физические, химические, биологические): температура, влажность, экотоксиканты, фитопатогены [10]. При этом известно, что незначительное повышение уровня АФК (как сигнальных молекул) индуцирует в клетке ряд молекулярных, биохимических и физиологических реакций, способствующих формированию адаптивных механизмов и повышению устойчивости организма [11–15]. В то же время слишком высокий уровень АФК может инициировать разветвленные цепи окислительных реакций, приводящих к повреждениям макромолекул (белки, ДНК), клеточных структур и в конечном итоге – к гибели клетки [15, 16]. Ключевая роль в регуляции стационарного уровня АФК в клетке принадлежит сбалансированному функционированию систем про- и антиоксидантной защиты. При этом антиоксидантная система включает ферменты и низкомолекулярные компоненты, которые нейтрализуют избыток АФК и обеспечивают защиту биологических структур клетки, замедляя (или предотвращая) окисление внутриклеточных органических соединений и участвуя в детоксикации вторичных метаболитов [15, 17].
Индукторами образования АФК в клетке являются многие соединения, которые активируют ее сигнальную систему и защитные реакции. В частности, салициловая кислота (СК) в настоящее время рассматривается как эндогенный, полифункциональный биорегулятор фенольной природы, принимающий участие в клеточном сигналинге, ростовых процессах и формировании адаптивных реакций растений [18, 19]. Известно, что СК сочетает в себе свойства сигнального посредника и антистрессового фитогормона [20–22]. Как полагают, важной функцией эндогенной СК является модификация эффектов АФК, что связано с ее разнонаправленным влиянием на ключевые про- и антиоксидантные ферменты.
Участие СК в реализации ответных реакций растений на действие абиотических стрессоров изучено в значительно меньшей степени, чем ответные реакции на инфицирование патогенами [23–25]. Тем не менее действие СК на устойчивость растений обычно связывают с инактивацией каталазы и соответствующим увеличением количества пероксида водорода [26]. В то же время показано, что формирование АФК-опосредованного сигнала у растений под действием СК сопровождается активацией ферментов, участвующих в метаболизме активных радикалов кислорода: НАДФН-оксидазы, суперок-сиддисмутазы и пероксидазы [16, 18, 27, 28].
Цель работы . Оценить уровень световой эмиссии мицелия гриба Neonothopanus nambi и активность ферментов, участвующих в метаболизме АФК (НАДФН-оксидаза, каталаза, супер-оксиддисмутаза, пероксидаза), без и при воздействии экзогенной салициловой кислоты в условиях стресса.
Материалы и методы. Исследования выполнены с образцами мицелия гриба N. nambi , выращенными на жидкой питательной карто-фельно-сахарозной среде по технологии, разработанной нами ранее [1]. Для исследований из полученного пленочного мицелия высекали диски диаметром 12 мм [8]. Диски помещали в свежую питательную картофельно-сахарозную среду, в которую добавляли СК до финальной концентрации в диапазоне 0,1–5,0 мМ. Исходный раствор СК был предварительно нейтрализован до нейтральных значений рН. Контрольными являлись диски мицелия, которые инкубировали в питательной среде без добавления СК. Через равные промежутки времени (через каждый час) измеряли уровень световой эмиссии образцов мицелия с помощью люминометра Glomax 20/20 (Promega, USA), калиброванного по радиоактивному стандарту Гастингса-Вебера (одна люминесцентная единица (LU) составляет 2.7 · 103 квантов в 1 секунду). Активность ферментов определяли в экстрактах из биомассы мицелия. Активность СОД определяли методом [29]. Для определения активности каталазы использовали метод [30]. Активность НАДФН-оксидазы определяли по методу [31]. Активность пероксидазы оценивали по интенсивности окисления о-фенилендиамина в присутствии
Н 2 О 2 [32]. Спектральные исследования образцов при определении активности ферментов проводили с помощью спектрофотометра UV-1800 (Shimadzu, Japan).
Результаты и их обсуждение. Как показали эксперименты (рис. 1), у контрольных образцов мицелия, помещенных в питательную среду, наблюдается значительное (на 2 порядка и более) увеличение уровня световой эмиссии – от 6,6 · 106 LU до 1,1 · 109 LU. Свечение мицелия достигает максимальных значений через 3 часа инкубации, после чего наблюдается его медленное снижение. Эти данные согласуются с результатами наших предыдущих исследований, в которых было установлено, что механическое повреждение мицелия N. nambi сопровождается увеличением его люминесценции [1, 6, 8]. В то же время из представленных данных следует, что наличие СК в инкубационной среде заметным образом подавляет свечение поврежденного мицелия. Этот эффект является дозозависимым – повышение концентрации СК приводит к большему снижению уровня световой эмиссии (рис. 1). Видно, что ингибирование люминесценции наблюдается практически сразу после помещения образцов мицелия в питательную среду, содержащую СК. При этом было установлено, что зависимость снижения свечения мицелия N. nambi от концентрации СК имеет экспоненциальный вид (рис. 2).
Исследования показали (рис. 3), что у контрольных образцов мицелия, вне зависимости от интенсивности их свечения при инкубации, уровень активности НАДФН-оксидазы практически не меняется в ходе всего эксперимента. Однако было показано, что в присутствии экзогенной СК активность фермента увеличивается и этот эффект возрастает с увеличением концентрации кислоты (рис. 3). Видно, что повышение активности фермента становится заметным уже при 0,1 мМ концентрации СК в среде инкубации. Наблюдаемая активация НАДФН-оксидазы в мицелии N. nambi может свидетельствовать, что в присутствии СК в поврежденном грибе происходит более интенсивная генерация супероксид анион радикала (О 2 -).
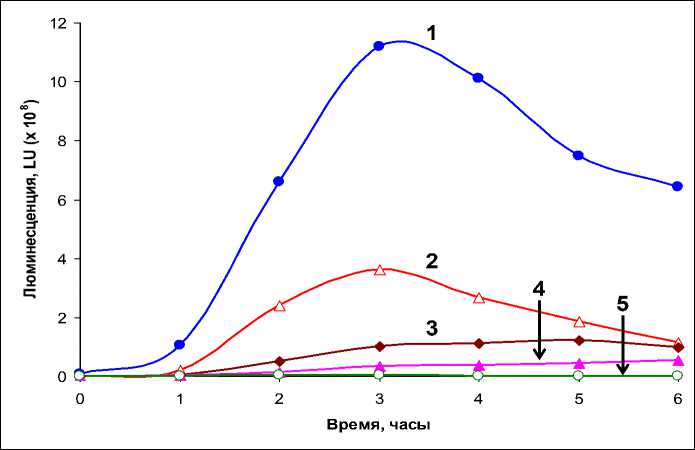
Рис. 1. Эффект салициловой кислоты (СК) на люминесценцию образцов мицелия N. nambi в зависимости от времени инкубации: 1 – контроль (без добавления СК); 2–5 – концентрация СК в инкубационной среде 0,1; 0,5; 1; 2; 5 и 5 мМ соответственно
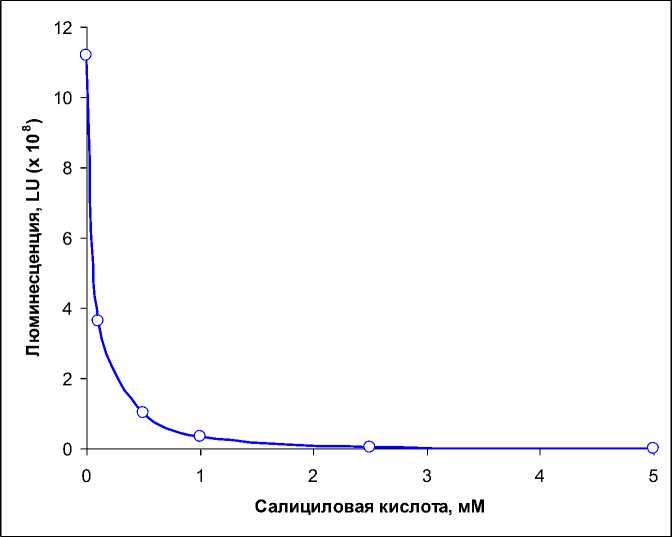
Рис. 2. Максимум интенсивности световой эмиссии мицелия N. nambi в зависимости от концентрации салициловой кислоты
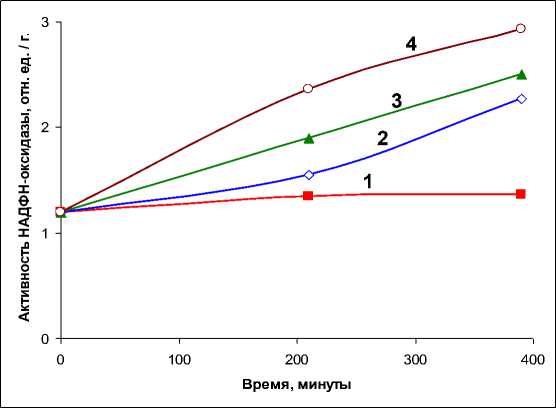
Рис. 3. Активность НАДФН-оксидазы в образцах мицелия N. nambi в зависимости от времени инкубации без (1) и при добавках (2–4) СК в инкубационную среду до концентрации 0,1, 0,5 и 1 мМ соответственно. Активность фермента представлена в относительных единицах на 1 грамм сырой биомассы мицелия
Активность СОД (рис. 4) в контрольных образцах мицелия снижается через 3 часа инкубации в 1,5 раза по сравнению с исходным уровнем и остается практически неизменной в течение последующих 3–4 часов. Из представленных данных видно, что наличие в инкубационной среде экзогенной СК в концентрации 0,1 мМ незначительно повышает уровень активности СОД. Однако при увеличении концентрации СК на порядок (1 мМ) активность СОД существенно возрастает через 3 часа инкубации мицелия (рис. 4). После этого активность СОД снижается до значений, соответствующих величине активности фермента в контрольном мицелии. Исходя из этого, можно предполагать, что в условиях острого эксперимента (травма) в короткие периоды времени (1–3 часа инкубации) под действием экзогенной СК происходит активация СОД. В свою очередь, это может свидетельствовать об активации образования пероксида водорода в грибе в этот период.
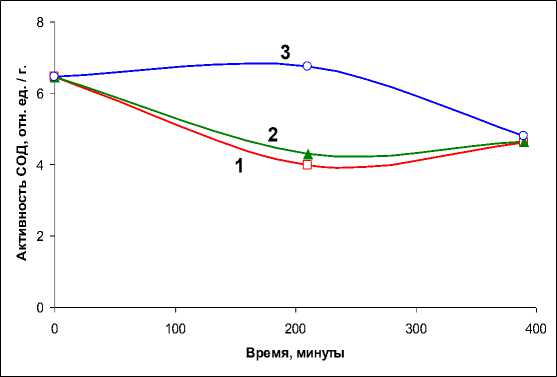
Рис. 4. Активность СОД в образцах мицелия N. nambi в зависимости от времени инкубации без (1) и при добавках (2, 3) СК в инкубационную среду до концентрации 0,1 и 1мМ соответственно. Активность фермента представлена в относительных единицах на 1 грамм сырой биомассы мицелия
В экспериментах были выявлены аналогичные изменения в общей пероксидазной активности (рис. 5). Из полученных данных следует, что в контрольных образцах мицелия уровень пероксидазной активности снижается практически в 2 раза через 3 часа инкубации, и эта тенденция сохраняется в последующие 3–4 инкубации. Однако в присутствии экзогенной СК уровень пероксидазной активности возрастает в 1,5–2 раза (рис. 5), что указывает на активацию данных ферментов в мицелии. Вероятно, наблюдаемое повышение общей пероксидазной активности может быть вызвано активацией образования в грибе пероксида водорода (см. предыдущий раздел) и, как следствие, необходимостью нейтрализации этой АФК. В условиях острого опыта этот эффект должен проявляться на ранних временных стадиях, что и наблюдалось экспериментально.
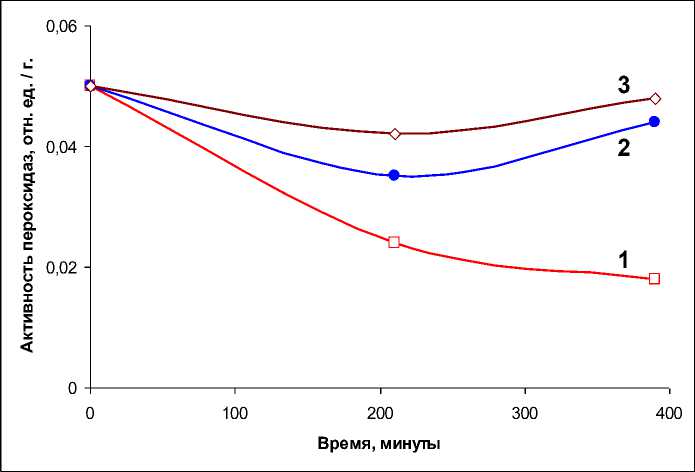
Рис. 5. Общая пероксидазная активность в образцах мицелия N. nambi в зависимости от времени инкубации без (1) и при добавках (2–4) СК в инкубационную среду до концентрации 0,5 и 1 мМ, соответственно. Пероксидазная активность представлена в относительных единицах на 1 грамм сырой биомассы мицелия.
Показано, что активность каталазы (рис. 6) в контрольных образцах механически поврежденного мицелия незначительно повышается через 1,5 часа инкубации. Затем активность фермента снижается и в последующие 3–4 часа инкубации практически не изменяется, хотя и отмечается некоторая тенденция к повышению уровня каталазной активности. Из представленных данных видно, что добавка СК в среду инкубации приводит к снижению активности фермента. По крайней мере, через 1,5 часа инкубации мицелия в присутствии СК наблюдается незначительное понижение уровня каталазной активности. Видно (рис. 6), что эффект является дозозависимым – при большей концентрации СК регистрируется меньшая активность фермента. В течение последующего периода инкубации активность фермента в контрольных и опытных образцах мицелия практически не различается.
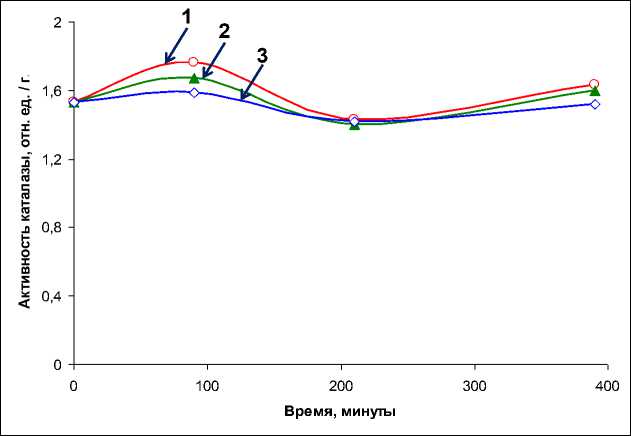
Рис. 6. Активность каталазы в образцах мицелия N. nambi в зависимости от времени инкубации без (1) и при добавках (2–3) СК в инкубационную среду до концентрации 0,5 и 5 мМ соответственно. Активность фермента представлена в относительных единицах на 1 грамм сырой биомассы мицелия
В завершение работы следует отметить, что во всех исследованных случаях при инкубации образцов мицелия в присутствии СК наблюдалось желто-коричневое окрашивание питательной среды, интенсивность которого возрастала в зависимости от времени инкубации и концентрации СК.
Заключение. Совокупность представленных в работе данных позволяет высказать несколько общих суждений и выводов. Показано, что в условиях острого эксперимента, после механического повреждения мицелия N. nambi и последующей его инкубации в питательной среде в течение короткого (6–7 часов) интервала времени, наблюдается значительное (на 2 порядка и более) увеличение уровня световой эмиссии гриба. Повышение интенсивности свечения регистрируется на фоне существенного снижения в мицелии активности СОД и общей пероксидазной активности. При этом уровень активности НАДФН-оксидазы в грибе практически не меняется в течение всего эксперимента, а активность каталазы незначительно возрастает. Установлено, что добавка экзогенной СК в среду инкубации заметным образом подавляет световую эмиссию мицелия N. nambi. При этом выявлено, что экзогенная СК активирует все изучаемые ферменты метаболизма АФК, за ис- ключением каталазы, активность которой несущественно снижается. Установлено, что эффекты СК являются дозозависимыми и нарастают с увеличением концентрации реагента. В целом представленные в работе данные согласуются с результатами других авторов, которые показали в исследованиях на растениях изменение активности ферментов метаболизма АФК под действием СК. В то же время полученные данные согласуются с высказанной нами ранее гипотезой, что грибное свечение является дополнительным механизмом антиоксидантной защиты от повреждающего действия АФК. Наблюдаемое в экспериментах значительное увеличение световой эмиссии мицелия при снижении активности СОД, и особенно общей пероксидазной активности, может свидетельствовать о запуске в грибе этого механизма защиты от АФК (прежде всего от избытка Н2О2 и иных перок-сидных соединений), которые могут нейтрализоваться в реакции свечения. В присутствии экзогенной СК ферменты метаболизма АФК активируются, следовательно, лидирующая роль в нейтрализации АФК осуществляется уже классической системой антиоксидантной защиты. Защитная функция системы излучения становится второстепенной, и свечение гриба снижается. Иным объяснением снижения свечения мицелия под действием СК может быть следующее. Общеизвестно, что СК является фенольным соединением и это позволяет рассматривать ее как субстрат для пероксидазных ферментов. Исходя из этого, экзогенная СК может осуществлять не только свою регуляторную функцию, активируя в мицелии ферменты метаболизма АФК, но и участвовать в нейтрализации активных радикалов кислорода (прежде всего, пероксида водорода) как субстрат в ферментативных реакциях с участием грибных пероксидаз (например, экстраклеточных). Известно, что защитное действие пероксидаз в значительной степени определяется их способностью окислять соединения фенольной природы до хинонов [33], которые являются хромогенами и составляют структурную основу пигментов. Вероятно, наблюдаемое нами в экспериментах окрашивание питательной среды при инкубации поврежденного мицелия в присутствии СК могло являться следствием окисления салициловой кислоты грибными пероксидазами.
Список литературы Активность ферментов метаболизма активных форм кислорода и уровень световой эмиссии мицелия гриба Neonothopanus nambi при действии салициловой кислоты
- Бондарь В.С., Пузырь А.П., Пуртов К.В. . О люминесцентной системе светя-щегося гриба Neonothopanus nambi//ДАН. -2011. -Т. 438. -№ 5. -С. 705-707.
- Bondar V.S., Shimomura O., Gitelson J.I. Lu-minescence of higher mushrooms//J. Sib. Fed. Univ. Biol. -2012. -V. 5. -№. 4. -P. 331-351.
- Бондарь В.С., Родичева Э.К., Медведева С.Е. . О механизме свечения гриба Neonothopanus nambi//ДАН. -2013. -Т. 449. -№ 2. -С. 223-227.
- Бондарь В.С., Пузырь А.П., Пуртов К.В. . Выделение люминесцентной систе-мы из светящегося гриба Neonothopanus nambi//ДАН. -2014. -Т. 455. -№ 3. -С. 346-348.
- Могильная О.А., Ронжин Н.О., Медведева С.Е. . Общая пероксидазная и ката-лазная активности светящихся базидиоми-цетов Armillaria borealis и Neonothopanus nambi в сравнении с уровнем световой эмиссии//Прикладная биохимия и микро-биология. -2015. -Т. 51. -№ 4. -С. 395-401.
- Medvedeva S.E., Artemenko K.S., Krivosheenko A.A. . Growth and light emission of luminous basidiomycetes cultivat-ed on solid media and in submerged culture//Mycosphere. -2014. -V. 5. -P. 565-577.
- Kobzeva T.V., Melnikov A.R., Karogodina T.Y. . Stimulation of Luminescence of myceli-um of luminous fungus Neonothopanus nambi by ionizing radiation//Luminescence. -2014. -V. 29. -P. 703-710.
- Тюлькова Н.А., Медведева С.Е., Бондарь В.С. Сравнительная оценка интенсивностей перекисного окисления липидов и свечения гриба Neonothopanus nambi//Вестник КрасГАУ. -2016. -№ 1. -С. 21-28.
- Mogilnaya O.A., Ronzhin N.O., Bondar V.S. Comparative evaluation of total peroxidase and catalase activities during light emission of luminous fungus Neonothopanus nambi//Mycosphere. -2016. -V. 7. -P. 499-510.
- Desican R., Mackerness S.A., Hancock J.T. . Regulation of the Arabidopsis transcriptome by oxidative stress//Plant Physiol. -2001. -V. 127. -P. 159-172.
- Тарчевский И.А. Сигнальные системы кле-ток растений. -М.: Наука, 2002. -294 с.
- Гесслер Н.Н., Аверянов А.А., Белозерская Т.А. Активные формы кислорода в регуля-ции развития грибов//Биохимия. -2007. -Т. 72. -№ 10. -С. 1091-1109.
- Vranova E., Inze D., Van Breuegegem F. Sig-nal transduction during oxidative stress//J. Exp. Bot. -2002. -V. 53. -P. 1227-1236.
- Jaspers P., Kangasjarvi J. Reactive oxygen species in abiotic stress signaling//Physiol. Plant. -2010. -V. 138. -P. 405-413.
- Колупаев Ю.Е., Карпец Ю.В. Активные формы кислорода при адаптации растений к стрессовым температурам//Физиология и биохимия культурных растений. -2009. -Т. 41. -№ 2. -С. 95-108.
- Карпун Н.Н., Янушевская Э.Б., Михайлова Е.В. Механизмы формирования неспеци-фического индуцированного иммунитета у растений при биогенном стрессе//Сель-скохозяйственная биология. -2015. -Т. 50. -№ 5. -С. 540-549.
- Apel K., Hirt H. Reactive oxygen species: me-tabolism, oxidative stress, and signal transduction//Annu. Rev. Plant Biol. -2004. -V. 55. -P. 373-399.
- Колупаев Ю.Е., Ястреб Т.О. Стресс-протекторные эффекты салициловой ки-слоты и ее структурных аналогов//Физио-логия и биохимия культурных растений. -2013. -Т. 45. -№ 2. -С. 113-126.
- Белых Ю.В., Кириллова Н.В., Спасенков А.И. Влияние салициловой кислоты на ан-тиоксидантную и прооксидантную активно-сти в растительных клетках//Вестник Санкт-Петербургского университета. -2009. -Вып. 2. -С. 145-151.
- Молодченкова О.О. Влияние салициловой кислоты на ответные реакции проростков кукурузы при абиотических стрессах//Вiсник Харкiвського нацiонального аграрно-го унiверситету. -2008. -Вип. 3(15). -С. 24-32.
- Абилова Г.А. Участие салициловой кислоты в системе антиоксидантной защиты у три-тикале при действии ZnSO4//Вестник Да-гестанского государственного университе-та. -2013. -Вып. 1. -С. 124-127.
- Фенько А.А., Репкина Н.С., Таланова В.В. Влияние салициловой кислоты на холодо-устойчивость проростков огурца//Тр. Ка-рельского научного центра РАН. -2015. -№ 11. -С. 26-34.
- Fujita M., Fujita Y., Noutoshi Y. . Cross-talk between abiotic and biotic stress respons-es: a current view from the points of conver-gence in the stress signalling networks//Curr. Opin. Plant Biol. -2006. -№ 9. -P. 436-442.
- Kaur N., Gupta A.K. Signal transduction path-ways under abiotic stresses in plant//Curr. Sci. -2005. -V. 88. -№ 11. -P. 1771-1780.
- Lushchak V.I. Adaptive response to oxidative stress: Bacteria, fungi, plants and animals//Comp. Biochem. Physiol. Part C. -2011. -V. 153. -P. 175-190.
- Chen Z., Silva H., Klessig D.F. Active oxygen species in the induction of plant systemic ac-quired resistance by salicylic acid//Science. -1993. -V. 262. -№ 5141. -P. 1883-1886.
- Barna B., Adam A.L., Gulner G. . Role of antioxidant systems and juvenility in tolerance of plants to diseases and abiotic stresses//Acta Phytopathol. Entomol. Hung. -1995. -V. 30. -P. 39-45.
- Rao M.V., Paliyaht G., Ormrod D.P. . Influence of salicylic acid on H2O2 production, oxidative stress, and H2O2-metabolizing en-zymes (salicylic acid-mediated oxidative dam-age requires H2O2)//Plant Physiol. -1997. -V. 115. -P. 137-149.
- Полесская О.Г., Каширина Е.И., Алехина Н.Д. Изменение активности антиоксидант-ных ферментов в листьях и корнях пшени-цы в зависимости от формы и дозы азота в среде//Физиология растений. -2004. -Т. 51. -С. 686-691.
- Aeby H. Catalase in vitro//Methods Enzymol. -1984. -V. 105. -P. 121-126.
- Вольский H.H., Козлов В.А., Лозовой В.П. Влияние гидрокортизона на продукцию су-пероксидного радикала фагоцитирующими клетками селезенки//Бюл. эксперимен-тальной биологии и медицины. -1987. -Т. 103. -№ 6. -С. 694-696.
- Wi S.J., Ji N.R., Park K.Y. Synergistic biosyn-thesis of biphasic ethylene and reactive oxy-gen species in response to hemibiotrophic phytophthora parasitica in tobacco plants//Physiol. Plant. -2012. -V. 159. -№ 1. -P. 251-265.
- Okey E.N., Duncan E.J., Sirju-Charran G. Phy-tophthora canker resistance in cacao: Role of peroxidase, polyphenoloxidase and phenylal-anine ammonialyase//J. Phytopathol. -1997. -V. 145. -№ 7. -P. 295-299.
