Активность гваякол-зависимой пероксидазы в листьях Plantago major L

Автор: Живетьев М.А., Граскова И.А., Войников В.К.
Журнал: Журнал стресс-физиологии и биохимии @jspb
Статья в выпуске: 3 т.9, 2013 года.
Бесплатный доступ
Впервые была изучена активность гваякол-зависимых пероксидаз при разных значениях рН в листовых пластинках Plantago major L.
Рн
Короткий адрес: https://sciup.org/14323777
IDR: 14323777
Текст научной статьи Активность гваякол-зависимой пероксидазы в листьях Plantago major L
Впервые была изучена активность гваякол-зависимых пероксидаз при разных значениях рН в листовых пластинках Plantago major L.
Оптимум рН катализируемой реакции – важная характеристика ферментов. В частности, пероксидазе, выделенной из томатов, свойственен оптимум рН равный 6,6 (Signoret, Crouzet, 1978), из полыни – 5,2 (Садвакасова, 1981). Пероксидаза хрена имеет несколько оптимумов – при рН 4,5 (Maehly, 1955) и 7,0 (Gaspar et al., 1977), что обусловлено наличием нескольких молекулярных форм фермента, проявляющих максимальную активность в более узких пределах рН (Газарян и др., 1998). Более того, в зависимости от рН среды фермент может катализировать разные реакции. Так, пероксидаза красной водоросли при рН 6,7 катализирует реакцию гидроксилирования фенолов, а при рН 5,4 – реакцию их бромирования в присутствии NaBr (Андреева, 1988). Максимальная активность пероксидазы перикарпа китайской сливы наблюдается при рН 6,5 и 70°С, а полифенолоксидазы – при рН 6,5 – 6,75 и 20°С (Mizobutsi et al., 2010). Кроме того, у растений с наступлением осени в период с июля по сентябрь может наблюдаться увеличение активности этого фермента (Zolfaghari et al., 2010). Вероятно, это связано с тем, что устойчивость растительных тканей к различным стрессовым воздействиям определяется в немалой степени активностью ферментов, обладающих пероксидазной активностью (Долгова, 2004; Янчевская и др., 2006; Ершова, 2007; Жуйкова и др., 2009; Алиева и др., 2010; Müftlügil, 1985; Thongsook, Barrett, 2005). В то же время, не смотря на интерес к этому ферменту, пероксидаза у многих растительных видов изучена не достаточно.
В этой связи целью наших исследований было проследить изменения в активности гваякол-зависимой пероксидазы подорожника большого в течение июня, июля и августа при значениях рН 4,6, 5,4 и 6,2 в двух различных точках ареала вида – г. Иркутске и на юговосточном побережье Байкала в районе устья речки Выдриная.
МАТЕРИАЛЫ И МЕТОДЫ
Материал для исследования. Исследовались листья подорожника большого Plantago major L., собранного в течение вегетационного периода 2009 года на территории стационара «Речка Выдринная» и в г. Иркутске.
Выделение слабосвязанной с клеточной стенкой пероксидазы. Для выделения пероксидаз слабосвязанных с клеточной стенкой, навеску (1 г) ткани листьев помещали в шприц с 5 мл холодного цитратно-фосфатного буфера (рН 5,5) и выдерживали при разреженном давлении 2–3 раза по 1 мин (Граскова и др., 2004). Полученный раствор центрифугировали 15 мин при 3 тыс. об./мин, отделяя от частиц ткани, и супернатант использовали для дальнейшего определения активности фермента.
Выделение растворимой пероксидазы. Навеску ткани после экстракции слабосвязанных пероксидаз помещали в 5 мл холодного цитратно-фосфатного буфера и растирали в фарфоровой ступке при 4 °С (Паду, 1995) для выделения растворимых пероксидаз Полученный гомогенат центрифугировали при 3 тыс. об./мин в течение 15 мин. Супернатант использовали для определения активности фермента.
Определение пероксидазной активности. Активность пероксидаз в листьях растений определяли по изменению оптической плотности (длина волны 580 нм) в реакционной смеси следующего состава: 0,5 мл 0,1 М цитратно-фосфатного буфера (рН от 4,6 до 6,2 с шагом 0,8), 0,5 мл 0,3 % перекиси водорода («Реахим», Россия), 0,5 мл 0,05 % гваякол (Sigma, США) и 0,5 мл пробы (1г пробы растирали в 10 мл Цитратно-фосфатного буфера рН 5,5).
Активность пероксидазы определяли при 25 °С сразу после выделения ферментов из образцов. Активность фермента рассчитывали по методу Бояркина (Бояркин, 1951) и выражали в условных единицах на мг сырого веса тканей по формуле: А = ∑ (α β γ)/d t, где ∑ - экстинция (0,125), α – отношение количества буфера, взятого для приготовления вытяжки в мл к весу сырой ткани, β – степень дополнительного разведения вытяжки в реакционной смеси, γ – степень постоянного разведения вытяжки в реакционной смеси, d – толщина поглощающего слоя кюветы, t – время реакции.
РЕЗУЛЬТАТЫ
В точке отбора проб на берегу озера Байкал активность слабосвязанной пероксидазы у подорожника большого была высокой в июне, заметно снижалась к июлю, но начинала возрастать в августе, причем главным образом за счет активности при рН 5,4 и 6,2 (рис. 1).
Для растворимой фракции гваякол-зависимой пероксидазы картина была обратная (рис. 2). Минимум активности при всех вариантах рН наблюдался в июне, а максимум – в июле.
Сходные данные были получены и по активности пероксидазы в июле и августе для г.
Иркутск (рис. 3, 4). В июле активность слабосвязанной с клеточной стенкой пероксидазы не зависела от рН в обоих точках сбора и составляла порядка 0,02 у.е./мг. В отличие от Байкала, в городе Иркутск повышение активности слабосвязанной с клеточной стенкой гваякол-зависимой пероксидазы в августе происходило за счет повышения ее активности при рН 6,2.
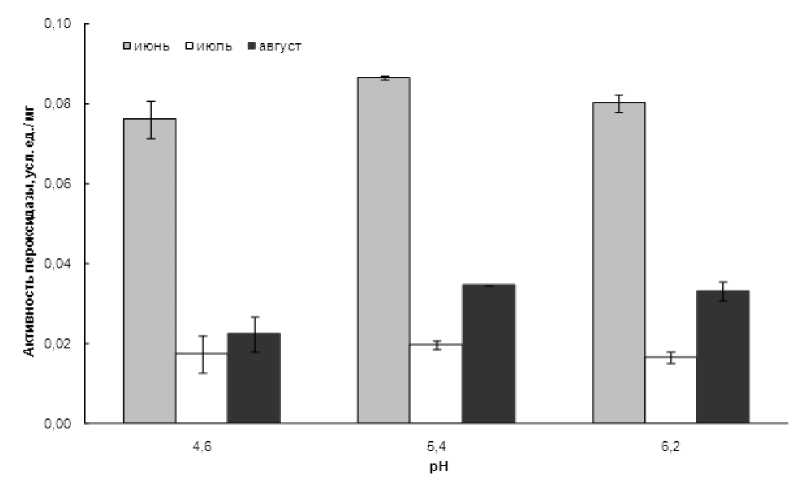
Рисунок 1 Активность слабосвязанной с клеточной стенкой пероксидазы листьев Plantago major в июне, июле и августе, Байкал
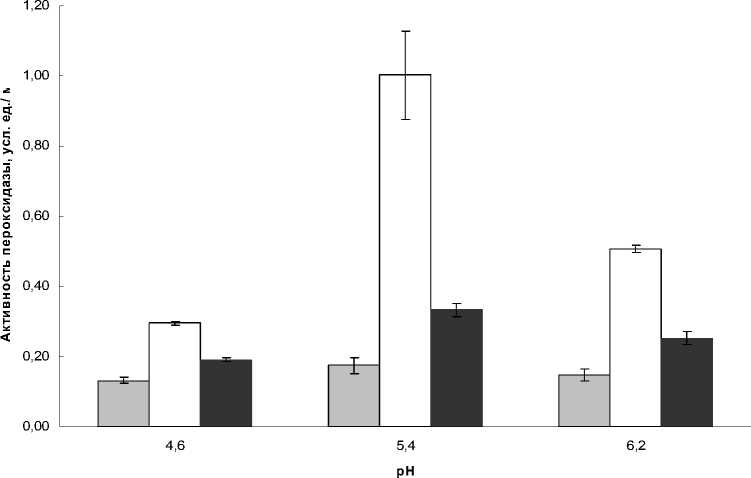
Рисунок 2 Активность растворимой пероксидазы листьев Plantago major в июне, июле и августе, Байкал
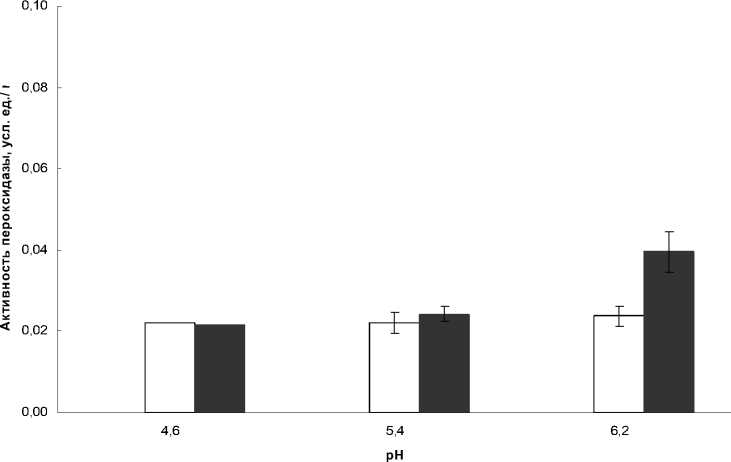
Рисунок 3 Активность слабосвязанной с клеточной стенкой пероксидазы листьев Plantago major в июле и августе, Иркутск
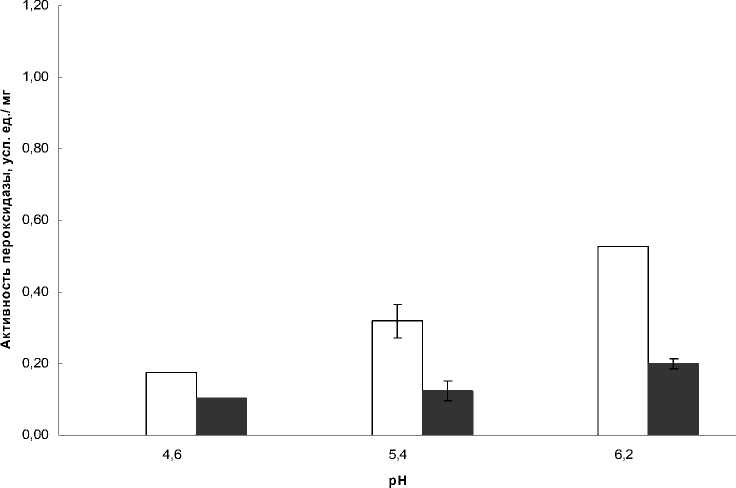
Рисунок 4 Активность растворимой пероксидазы листьев Plantago major в июле и августе Иркутск
Активность растворимой пероксидазы, не смотря на схожесть динамики, была выше на Байкале, чем в Иркутске, за исключением рН 6,2, когда данные по обоим географическим точкам совпали и составили 0,5 у.е./мг для июля и 0,2 для августа. Максимальные значения активности растворимой пероксидазы для Иркутска наблюдались при этой рН (6,2), а на побережье Байкала активность была максимальна при рН 5,4 и в июле, и в августе.
На основании полученных данных можно предполагать следующее: не смотря на то, что растворимая пероксидаза обычно доминирует в растительных тканях, адаптация к осенним сезонным изменениям у одуванчика связана главным образом с увеличением активности слабосвязанной пероксидазы как более подвижной фракции этого фермента. В то же время, влияние города Иркутск выражалось в уменьшении активности не слабосвязанной, а растворимой гваякол-зависимой пероксидазы подорожника при всех значениях рН как в июле, так и в августе по сравнению с побережьем Байкала.
Список литературы Активность гваякол-зависимой пероксидазы в листьях Plantago major L
- Алиева Д.Р., Бабаев Г.Г., Азизов И.В. (2010) Активность и изоферментный состав пероксидазы клеток Dunaliella salina при солевом стрессе. Вестник Днепропетровского университета. Биология. Медицина. 1(1), 16-21.
- Андреева В.А. (1988) Фермент пероксидаза. М.: Наука, 128 с.
- Бояркин А.Н. (1951) Быстрый метод определения активности пероксидазы Биохимия. 16. 352.
- Газарян И.Г., Упоров И.В., Чубарь Т.А., Федчина В.А., Мареева Е.А., Лагримини Л.М. (1988) Влияние рН на стабильность анионной пероксидазы табака и ее взаимодействие с перекисью водорода Биохимия. 63(5). 708-716.
- Граскова И.А., Боровский Г.Б., Колесниченко А.В., Войников В.Л. (2004) Пероксидаза как компонент сигнальной системы клеток картофеля при патогенезе кольцевой гнили Физиология растений 51(5). 692-697.
- Долгова Л.Г. (2004) Активность пероксидазы -показатель устойчивости растений-интродуцентов в условиях степной зоны Украины. Вестник Днепропетр. Ун-та. 1. 38-41.
- Ершова А.Н. (2007) Метаболическая адаптация растений к гипоксии и повышенному содержанию диоксида углерода. -Воронеж.: Изд-во Воронеж. гос. ун-та,. -264с.
- Жуйкова Т.В., Безель В.С. (2009) Адаптация растительных систем к химическому стрессу: популяционный аспект. Вестник Удмуртского университета, 1. 31-42.
- Паду Э.Х. (1995) Свойства пероксидазы и фенилаланин-аммиак-лиазы при образовании и лигнификации клеточных стенок стебля пшеницы. Физиология растений. 42. 408-415.
- Садвакасова Г.Г. (1981) Пероксидаза полыни и окислительный распад флавоноидов (на примере кверцитина): Автореф. дис. …канд. биол. наук. Алма-Ата,. 25 с.
- Янчевская Т.Г., Гриц А.Н, Ковалёва О.А. (2006) Влияние ультрафиолетового облучения суммарного диапазона на активность пероксидазы листьев меристемных регенерантов картофеля (Sоlanum tuberosum). Вести БГПУ., № 2. Серия 3. 38-40.
- Gaspar N., Wyndacker R., Bouchet M., Ceulemans E. (1977) Peroxidase and amylase activities in relation to germination of dormant and nondormant wheat. Physiol. Plant. 40(1). 11-14.
- Kareska S. (2009) Factors affecting hydrogen peroxidase activity. ESSAI. 7(27). 82-85.
- Maehly A.C. (1955) Plant peroxidase. Methods in Enzymology. 2. 801-813.
- Mizobutsi G.P., Finger F.L., Ribeiro R.A., Puschmann R., de Melo Neves L.L., da Mota W.F. (2010) Effect of pH and temperature on peroxidase and polyphenoloxidase activities of litchi pericarp. Sci. Agric. (Piracicaba, Braz.). 67(2). 213-217.
- Müftügil N. (1985) The peroxidase enzyme activity of some vegetables and its resistance to heat. J. Sci. Food Agric. 36. 877-880.
- Signoret A., Crouzed J. (1978) Activities polyphenoloxidasique et peroxidasigue du fruit de la tomate (Lycopersicum esculentum), purification et quelques properties. Arg. Biol. Chem. 42(10). 1527-1530.
- Thongsook T., Barrett D.M. (2005) Heat Inactivation and Reactivation of Broccoli Peroxidase. J. Agric. Food Chem. 53. 3215-3222.
- Zolfaghari R., Hosseini S.M., Korori S.A.A. (2010) Relationship between peroxidase and catalase with metabolism and environmental factors in Beech (Fagus orientalis Lipsky) in three different elevations. International J. of Environmental Sciences. 1(2). 243-252.
