Активность и состав лектинов клеточной стенки пшеницы при инфицировании грибными патогенами

Автор: Шаймуллина Гульназ Хидиятовна, Хусаинова Раиля Рафаиловна, Невмержицкая Юлия Юрьевна, Тимофеева Ольга Арнольдовна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биотехнология
Статья в выпуске: 3-5 т.15, 2013 года.
Бесплатный доступ
Исследована гемагглютинирующая активность и молекулярный состав лектинов из клеток корней проростков озимой пшеницы сорта Мироновская 808 и Казанская 560, инфицированных возбудителями грибных заболеваний. Методами очистки и фракционирования белков выявлено увеличение количества белков клеточной стенки с агглютинирующей активностью у двух сортов озимой пшеницы, что свидетельствует об участии этих белков в механизмах защитного действия при патогенезе. Показано, что увеличение лектиновой активности является одной из защитных реакций растительного организма на действие патогенов и зависит от устойчивости сорта и воздействующего инфекционного агента.
Лектины, очистка, патоген
Короткий адрес: https://sciup.org/148202077
IDR: 148202077 | УДК: 581.1
Lectin activity and composition in the wheat cell walls during infection fungal pathogens
Hemagglutinating activity and the molecular composition of the lectin wheat germ cells Mironovskaya 808 and Ka- zankaya 560 infected with pathogens of fungal diseases are investigated. The methods of purification and fractionation of proteins revealed an increase in the number of cell wall proteins with agglutinating activity in two varieties of winter wheat, which indicates the involvement of these proteins in the mechanisms of the protective effect of the pathogenesis. It is shown that an increase in activity of the lectin is one of the protective reaction of the organism to the action of plant pathogens and stability depend on the variety and exposure to infectious agent.
Текст научной статьи Активность и состав лектинов клеточной стенки пшеницы при инфицировании грибными патогенами
Проблема инфекционных болезней растений пшеницы, приводящая к потерям урожая и ограничению объемов производства сельскохозяйственных культур, не нова, но в последнее время приобрела глобальный характер. Поэтому важное значение приобретает изучение молекулярных основ устойчивости растений к фитозаболеваниям, в частности, к грибным патогенам [1].
При инфицировании растений фитопатогенами лектинам отводят роль первичных агентов в узнавании участков сложных углеводов на поверхности клеток патогенов и дальнейшем развитии реакций сверхчувствительности. Гипотезы об участии лектинов в защите растений от болезнетворных агентов подкреплены данными о способности этих белков специфически взаимодействовать с поверхностью бактериальных клеток, спор и гиф грибов [2], что приводит к несовместимому или совместимому взаимодействию организмов, которое соответственно проявляется в индукции или в отсутствии защитной реакции растений на атаку патогена [3].
Одним из признаков активной реакции растений на инфицирование является количественное изменение содержания или активности лектиновых белков. В связи с этим, цель данной работы состояла в анализе уровня активности и молекулярного состава лектинов при формировании иммунного ответа разных сортов озимой пшеницы ( Triticum aestivum L.), инфицированных фитопатогенами Fusarium ( F. ) oxysporum Schlectend.:Fr., Cladosporium ( C .) graminum Cda., Alternaria spp . и Aspergillus ( A. ) niger .
Объектом исследования служили проростки озимой пшеницы (Triticum aestivum L.) сорта Мироновская 808 и Казанская 560. Семена перед
посевом стерилизовали в 96% этаноле и промывали стерильной дистиллированной водой. Далее семена опытного варианта замачивали на сутки в суспензии конидий с исходным титром (1-3)*104 КОЕ/см3 (КОЕ - колониеобразующие единицы). Инфицирование проводили двумя специфическими для пшеницы фитопатогенными грибами Fusarium (F.) oxysporum Schlectend.:Fr. и Cladosporium (C.) graminum Cda. и двумя неспецифическими – Alternaria spp . и Aspergillus (A.) niger . Растения выращивали в лабораторных условиях в кюветах на водопроводной воде при освещенности 100 Вт/м2 и 12-часовом фотопериоде в течение 7 сут.
Растворимые лектины экстрагировали 0,05н HCl, лектины клеточной стенки – 0,05% раствором тритона Х-100 по методу, описанному в ранее опубликованной работе [4]. Для определения количества белка использовали метод Bradford [5]. Активность лектинов определяли в планшетах для иммунологических исследований по их способности агглютинировать трипсинизированные эритроциты 1 группы крови. Лектиновую активность рассчитывали по минимальному количеству белка, вызывающему агглютинацию эритроцитов (мкг бел-ка/мл)-1. Эритроциты получали и трипсинизирова-ли по методу Луцика [6]. Гель-фильтрацию проводили на колонке с Sephadex G-150, которая позволила отделить лектины от значительного количества низкомолекулярных примесей непротеиновой природы. Результаты обработали статистически, их считали достоверными при уровне погрешности ≤5% по критерию Стьюдента.
Во время прорастания семян на ранних этапах развития проростка синтезируется значительное количество лектинов [7], также как это происходит в ходе нормального онтогенеза растений. В наших экспериментах при инфицировании фитопатогены уменьшали активность растворимых лектинов у двух сортов озимой пшеницы (рис. 1). Вероятно, ингибирующее действие фитопатогенов на актив- ность растворимых лектинов может быть обусловлено взаимодействием углеводсвязывающих центров этих белков с инфекционными структурами патогенов, в результате которого лектины не могут участвовать в агглютинации эритроцитов. При этом чувствительность активности растворимых лектинов к патогенам была меньше у Мироновской 808. Возможно, у Казанской 560 происходит более эффективная инактивация фитопатогенов за счет усиления взаимодействия лектиновых белков с углеводными гаптенами грибов.
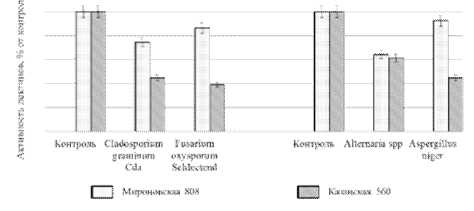
Рис. 1 . Активность растворимых лектинов озимой пшеницы сорта Мироновская 808 и Казанская 560 при инфицировании патогенными грибами
В литературе накоплено достаточное количество сведений, подтверждающих, что накопление лектинов тесно связано с индукцией устойчивого состояния растения-хозяина [8]. В наших опытах заражение растений специфическим фитопатогеном Cladosporium graminum Cda. приводило к значительному повышению активности лектинов клеточной стенки у растений двух сортов (рис. 2). При этом ответная реакция растений на Cladosporium graminum Cda. была выражена сильнее по сравнению с Fusarium spp. Fusarium spp. усиливал активность лектинов клеточной стенки только у сорта Мироновская 808. По-видимому, этот сорт более восприимчив к Fusarium spp. по сравнению с Казанской 560. Следует отметить, что и степень поражения этим патогеном растений Мироновской 808 была выше по сравнению с Казанской 560. При действии неспецифического возбудителя Alternaria spp. активность лектинов клеточной стенки снижалась, а Aspergillus niger не оказывал влияния на активность лектинов у двух сортов (рис. 2).
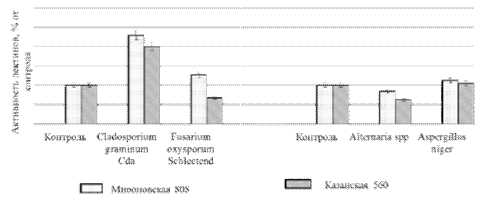
Рис. 2 . Активность связанных с клеточной стенкой лектинов озимой пшеницы сорта Мироновская 808 и Казанская 560 при инфицировании патогенными грибами
В следующей части работы с помощью гель- фильтрации мы провели хроматографическое разделение белков клеточной стенки корней проростков озимой пшеницы сортов Мироновская 808 и Казанская 560, инфицированных патогенными грибами.
У растений Мироновской 808 в контрольном варианте среди полученных белковых фракций лектиновая активность обнаруживалась во фракциях, содержащих белки с молекулярной массой 79, 69, 55, 45, 35 кДа (рис. 3). На основании работы Chivasa et al. [9], которые проанализировали белки клеточной стенки Arabidopsis thaliana , можно сделать некоторые предположения относительно обнаруженных нами лектинов клеточной стенки, поскольку белки арабидопсиса имеют состав, близкий к составу белков, которые характерны для других растений. Белок с молекулярной массой 35 кДа может быть классическим лектином пшеницы – агглютинином зародыша пшеницы (АЗП). АЗП, резкое накопление которого происходит при разных неблагоприятных условиях, относится к экскретируемым белкам, выделение которого в наружную среду может иметь значение для предохранения ослабленных растений от почвенной инфекции [8]. Обнаруженные нами белки наряду с лектиновой активностью, могут обладать ферментативной активностью. Например, белки с молекулярной массой 79 кДа и 45 кДа могут быть ГТФ-связывающими [9].
Патогенные микроорганизмы по-разному влияли на профиль элюции белков Мироновской 808 (рис. 3). Инфицирование неспецифическими грибами рода Alternaria приводило к исчезновению белков с молекулярной массой 79, 69 кДа и 45кДа. При этом в профиле элюции появились белки 85, 83, 71, 61, 59 кДа. В варианте с грибом Aspergillus niger не было только белков 69 кДа и 55 кДа, характерных для контрольных растений, при этом лектиновая активность обнаруживалась во фракциях 85, 83, 71, 61 кДа (эти же белки появлялись при действии Alternaria spp. ), а также белки 79, 51, 47, 38, 37, 33кДа и 29 кДа.
Кроме белков, выявленных у контрольных растений, в варианте с заражением Cladosporium gra-minum Cda. появились новые белки: 85, 81, 79, 51, 33 кДа и 29 кДа, а при действии Fusarium spp. мы обнаружили белки с молекулярной массой 87, 85, 75, 57, 55, 53, 45, 41, 35, 31 кДа (рис. 3).
В профиле элюции белков клеточной стенки контрольных растений сорта Казанская 560 лекти-новая активность была обнаружена во фракциях белков с молекулярной массой 79, 69, 61, 55, 45, 35 кДа. Заражение семян сорта Казанская 560 как специфическими, так и неспецифическими грибами вызывало изменение всего профиля элюции белков клеточной стенки (рис. 4).
В варианте с авирулентными микроорганизмами Alternaria spp. и Aspergillus niger значительно снизилось содержание высокомолекулярных белков, но возросла концентрация среднемолекулярных
а)
а)
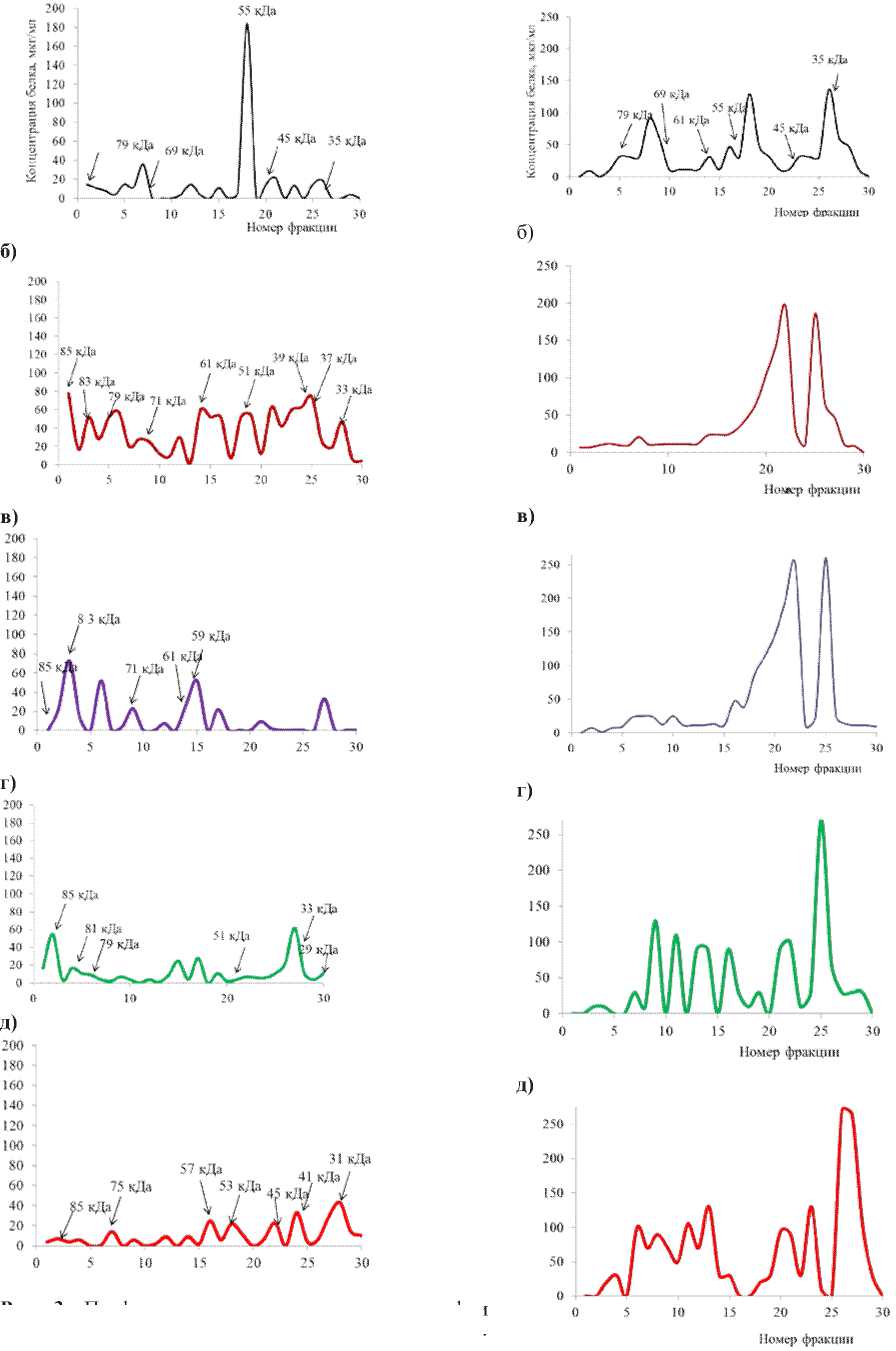
элюции при гель-хроматографии стенки проростков озимой пшени-
Рис. 3. Профиль белков клеточной цы Мироновская 808: а) контрольные растения, б) Aspergillus niger, в) Alternaria spp., г) Cladosporium gra-minum Cda. , д) Fusarium spp.
Рис. 4 . Профиль элюции при гель-хроматографии белков клеточной стенки проростков озимой пшеницы Казанская 560: а) контрольные растения, б) Aspergillus niger , в) Alternaria spp. , г) Cladosporium grami-num Cda., д) Fusarium spp .
полипептидов, которые формировали два характерных пика. При этом во всех выявленных фракциях наблюдалась лектиновая активность. Можно предположить, что индукция образования среднемолекулярных белков, обладающих лектиновой активностью, необходима для эффективного взаимодействия с авирулентными грибными патогенами.
Таким образом, обнаружено, что неспецифические микроорганизмы Alternaria spp. и Aspergillus niger вызывают у растений сорта Казанская 560 увеличение содержания среднемолекулярных лектиновых белков клеточной стенки, а специфические фитопатогены Cladosporium graminum Cda. и Fusarium spp. – высокомолекулярных лектиновых белков. По-видимому, эти две группы белков участвуют в узнавании и инактивации патогенов: среднемолекулярные взаимодействуют с авиру-лентными возбудителями, высокомолекулярные – с вирулентными.
-
1. Молодченкова О.О., Адамовская В.Г., Досенко В.Е., Тихонов П.С. Лектиновая активность и экспрессия генов лектина проростков пшеницы при инфицировании грибными патогенами и действии салициловой кислоты.// Вiсник Харкiвс. Нац. Агр. Ун-ту. Серiя Бiологiя.
2012. Т. 26. Bип. 2. С. 54-60.
-
2. Etzler M.E. Are lectins involved in plant-fungus interactions? // Phytopathology. 1981. V. 71. N 7. P. 744-746.
-
3. Белава В.Н., Зеленый С.Б., Панюта О.А.,Таран Н.Ю., Погребной П.В . Экспрессия генов лектинов и дефенсина у сортов пшеницы Мироновская 808 и Roazon при инфицировании Pseudocercosporella herpotrichoides // Biopolymers and Cell. 2010. V. 26. N. 1. P. 45-50.
-
4. Тимофеева О.А., Невмержицкая Ю.Ю., Московкина М.А. Активность и состав лектинов клеточной стенки пшеницы при действии низких температур и ингибиторов кальциевой сигнальной системы // Физиол. раст. 2010. Т. 57. № 2. С. 209-216.
-
5. Bradford M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding // Anal. Biochem. 1976. V. 72. P. 248-254.
-
6. Луцик М.Д., Панасюк Е.Н., Луцик А.Д . Лектины. Львов: Вища шк. Изд-во при Львов. ун-те, 1981. 156 с.
-
7. Lepekhin E.A., Yalovoi A.I., Rybak V.I . Activity and specificity carbohydrates in lectins of germinating maize grains // Russ. J. Plant Physiol. 1986. V. 33. N 2. P. 390-394.
-
8. Шакирова Ф.М. Неспецифическая устойчивость растений к стрессовым факторам и ее регуляция. Уфа: Гилем, 2001. 159 с.
-
9. Chivasa S., Ndimba B. K., Simon W . Proteomic analysis of the Arabidopsis thaliana cell wall // Electrophoresis. 2002. V. 23. P. 1754-1765.
LECTIN ACTIVITY AND COMPOSITION IN THE WHEAT CELL WALLS DURING INFECTION FUNGAL PATHOGENS
Список литературы Активность и состав лектинов клеточной стенки пшеницы при инфицировании грибными патогенами
- Молодченкова О.О., Адамовская В.Г., Досенко В.Е., Тихонов П.С. Лектиновая активность и экспрессия генов лектина проростков пшеницы при инфицировании грибными патогенами и действии салициловой кислоты.//Вiсник Харкiвс. Нац. Агр. Ун-ту. Серiя Бiологiя. 2012. Т. 26. Bип. 2. С. 54-60.
- Etzler M.E. Are lectins involved in plant-fungus interactions?//Phytopathology. 1981. V. 71. N 7. P. 744-746.
- Белава В.Н., Зеленый С.Б., Панюта О.А.,Таран Н.Ю., Погребной П.В. Экспрессия генов лектинов и дефенсина у сортов пшеницы Мироновская 808 и Roazon при инфицировании Pseudocercosporella herpotrichoides//Biopolymers and Cell. 2010. V. 26. N. 1. P. 45-50.
- Тимофеева О.А., Невмержицкая Ю.Ю., Московкина М.А. Активность и состав лектинов клеточной стенки пшеницы при действии низких температур и ингибиторов кальциевой сигнальной системы//Физиол. раст. 2010. Т. 57. № 2. С. 209-216.
- Bradford M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding//Anal. Biochem. 1976. V. 72. P. 248-254.
- Луцик М.Д., Панасюк Е.Н., Луцик А.Д. Лектины. Львов: Вища шк. Изд-во при Львов. ун-те, 1981. 156 с.
- Lepekhin E.A., Yalovoi A.I., Rybak V.I. Activity and specificity carbohydrates in lectins of germinating maize grains//Russ. J. Plant Physiol. 1986. V. 33. N 2. P. 390-394.
- Шакирова Ф.М. Неспецифическая устойчивость растений к стрессовым факторам и ее регуляция. Уфа: Гилем, 2001. 159 с.
- Chivasa S., Ndimba B. K., Simon W. Proteomic analysis of the Arabidopsis thaliana cell wall//Electrophoresis. 2002. V. 23. P. 1754-1765.
