Активность канонической wnt сигнальной системы в артикулярных хондроцитах гиалинового хряща в процессе формирования синовиального сустава

Автор: Кузнецов С.Л., Молотков А.О.
Журнал: Саратовский научно-медицинский журнал @ssmj
Рубрика: Макро- и микроморфология
Статья в выпуске: 1 т.5, 2009 года.
Бесплатный доступ
Каноническая Wnt сигнальная система (^пГСС) играет важную роль в регуляции остеогенеза, однако, до настоящего времени остается мало изученной роль этой системы в развитии синовиальных суставов и гиалинового хряща. Мы установили: 1) ^пГСС неактивна в формирующейся промежуточной зоне сустава и в дифференцирующихся артикулярных хондроцитах на ранних эмбриональных стадиях развития (Е10.5 -Е14.5); 2) KWnt^ активна в артикулярных хондроцитах на этапах позднего эмбрионального развития (Е18.5) и в течение постнатального развития (Р7 - Р10); 3) ^пГСС снова неативна в артикулярных хондроцитах гиалинового хряща сустава взрослых животных. Механизмы и биологическое значения показанной нами регуляции активности ^пГСС на разных этапах развития и дифференцировки артикулярных хондроцитов требует дальнейшего изучения.
Артикулярные хондроциты, развитие сустава
Короткий адрес: https://sciup.org/14916847
IDR: 14916847
Activity of canonical wnt signal system in hyaline cartilage articular chondrocytes in process of synovial joint development
Canonical and non-canonical Wnt systems are essential regulators of chondrogenesis and bone development. However, the roles of these systems in synovial joint development are not well studied. To determine if canonical Wnt system is active in developing articular chondrocytes we used immunohistochemistry for в-galactosidase and doublecortin (cell-type specific marker for articular chondrocytes) to double label sections through joint regions of E14.5, E18.5, P10 and adult mice. Here the following results are presented. Canonical Wnt signal system does not work in developing articular chondrocytes at early embryonic stages (E14.5); it is active in the articular chondrocytes at late embryonic stages (E16.5-E18.5) and during postnatal development (P7-P10), but is turned off again in the adult articular chondrocytes. These results suggest that canonical Wnt signaling is being regulated during articular chondrocytes differentiation and joint formation.
Текст научной статьи Активность канонической wnt сигнальной системы в артикулярных хондроцитах гиалинового хряща в процессе формирования синовиального сустава
Генетичес^ие исследования ^становили, что Wnt си^нальная система и^рает важн^ю роль в ре^^ляции развития с^елетной т^ани [6]. Тр^бчатые ^ости ^онеч-ностей челове^а и животных формир^ются в процессе вторично^о (непрямо^о) остео^енеза [15]. Мно^очис-ленными исследованиями ^становлено, что ^анони-чес^ая и не^аноничес^ая Wnt си^нальные системы ^частв^ют в ре^^ляции эндохондрально^о и перихон-дрально^о о^остенения [26, 9, 14]. В отличие от хорошо из^ченных механизмов формирования ^остной т^ани, механизмы развития синовиальных с^ставов (с^ставы, соединяющие ^остные элементы ^онечно-стей) из^чены недостаточно. Развитие с^става начинается с формирования промеж^точной зоны (interzone), разделяющей ^омо^енн^ю (непрерывн^ю) ^онденсацию мезенхимы на месте б^д^ще^о с^ста-ва, и образования трехслойной промеж^точной зоны [16]. Клет^и мезенхимы, расположенные в промеж^-точной зоне, в ходе дальнейшей дифференциров^и приводят ^ образованию различных ^леточных стр^^-т^р с^става (с^ставные связ^и, ^иалиновый хрящ и др.), в то время ^а^ ^лет^и мезенхимы, прилежащие ^ про-меж^точной зоне, в дальнейшем ^частв^ют в формировании зоны роста тр^бчатых ^остей и ^остной т^ани [22]. Имеющиеся на настоящее время данные свидетельств^ют о возможной роли Wnt в формировании синовиальных с^ставов и в ре^^ляции диффе-ренциров^и мезенхимы промеж^точной зоны [13]. Пос^оль^^ была по^азана э^спрессия Wnt4, Wnt9A, Wnt11, Wnt16 в месте формирования с^ставов [24], это позволяет предположить роль ^аноничес^ой Wnt си^нальной системы (^WntCC) в ре^^ляции развития стр^^т^р с^ставов и в ре^енерации их т^аней, измененных в рез^льтате старения или болезни [25]. Отметим, что в последние ^оды все более широ^ое распространение пол^чает заместительная ^леточная терапия (stem cell therapy) та^их распространенных заболеваний с^ставов, ^а^ остеоартрит и ревматоидный артрит [23], что делает проблем^ из^чения роли Wnt в развитии стр^^т^р с^ставов высо^о а^т^альной.
Се^одня известны три си^нальных системы, а^ти-вир^емых Wnt си^нальными моле^^лами: Wnt/Ca2+ система, Wnt/PCP (planar cell polarity) и β-^атенин зависимая ^WntCC системы. Основная роль ^WntCC системы за^лючается в ре^^ляции ^ровня свободно^о вн^т-ри^леточно^о β-^атенина, ^оторый, в свою очередь, ре^^лир^ет э^спрессию Wnt-зависимых ^енов [12]. В отс^тствии Wnt си^нала, свободный цитоплазменный в-катенин связывается комплексом GSK3a/APC/Axin, фосфорилир^ется и, в послед^ющем, разр^шается под воздействием протеаз. А^тивация ^WntCC предотвращает фосфорилирование и де^радацию β-^ате-нина, что приводит ^ повышению ^ровня свободно^о β-^атенина в цитоплазме, е^о трансло^ации в ядро ^лет^и, с послед^ющей а^тивацией транс^рипции Wnt-зависимых ^енов.
В исследованиях на первичной ^^льт^ре хондроцитов было ^становлено, что Wnt9A, э^спрессия ^ото-ро^о по^азана в т^анях с^става, а^тивир^ет ^WntCC в хондроцитах [25]. Генетичес^ие исследования на мышах, но^а^тированных по Wnt4 и Wnt9A (Wnt4-/-;Wnt9A-/-) ^становили ^ооперативное взаимодействие межд^ Wnt4 и Wnt9A в ре^^ляции развития с^ставов и поддержании их целостности [25]. Это позволило предположить, что Wnt4 и Wnt9A действ^ют через ^WntСС, направляя дифференциров^^ хондроцитов в зоне с^става. Одна^о последние работы ^бедитель-но по^азали, что Wnt4 ре^^лир^ет вн^три^леточн^ю ло^ализацию β-^атенина, вызывая передисло^ацию свободно^о β-^атенина ^ ^леточной мембране, тем самым предотвращая е^о трансло^ацию в ядро ^ле-то^ и ин^ибир^я а^тивацию ^WntСС [8]. В дополнении ^ этом^, Wnt11, э^спрессир^емый в т^анях с^ставов, взаимодейств^я с Lrp6 (^о-рецептор ^WntСС), ре^^ли-р^ет ^ровень Axin в цитоплазме ^лет^и [17]. Axin – ре^^лятор ^WntСС, о^раничивает ^ровень свободно-^о β-^атенина, ^величивая е^о фосфорилирование и протеолиз, и, та^им образом, снижает а^тивность ^WntСС [17].
В работе из^чена а^тивность ^WntСС в стр^^т^рах синовиальных с^ставов в процессе эмбрионально^о (E11 – E18), постнатально^о развития (P3 -P28) и ^ взрослых мышей. Использ^я линию TOPgal (Wnt-репортер) мышей для обнар^жения а^тивности ^WntСС в т^анях [10] и anti-doublecortin (DCX) антитела в ^а-честве специфичес^ой мет^и для арти^^лярных хондроцитов [27], мы ^бедительно продемонстрировали, что а^тивность ^WntСС ре^^лир^ется в арти^^ляр-ных хондроцитах во время эмбрионально^о и пост-натально^о развития. По^азано, что ^WntСС неа^тивна в мезенхиме промеж^точной зоны формир^юще^о-ся с^става на ранних стадиях эмбрионально^о развития (Е10 – Е14); а^тивна в дифференцир^ющихся ар-ти^^лярных хондроцитых на поздних стадиях эмбри-онально^о развития (Е16 – Е18) и в течение постна-тально^о развития (Р7 – Р10), и снова неа^тивна в арти^^лярных хондроцитах взрослых животных.
Материалы и методы. Для имм^но^истохими-чес^ой оцен^и э^спрессии lacZ, DCX и Vinculin ^риос-татные срезы (10-12 m), пол^ченные ^ TOPgal мышей на разных стадиях развития (Е14.5, Е18.5, Р7, Р10 и врослых 6-месячных животных), использовали сле-д^ющие первичные антитела: 1) ^роличьи антитела ^ а-галокгозидазе (lacZ) (1:1000, Cappel, MP Biomedicals, Solon, Ohio, USA), 2) мышиные антитела ^ Vinculin (1:100, Santa Cruse Biotechnology, CA, USA), 3) антитела из морс^ой свин^и ^ Doublecortin (DCX) (1:100, Abcam). Вторичные антитела разводили в 1% сыво-рот^е в 0.9% NaCl; во всех сл^чаях вторичные антитела использовали в разведении 1:1000. Были использованы след^ющие вторичные антитела: 1) Alexa Fluor 488 goat anti-mouse, 2) Alexa Fluor 594 goat antimouse, 3) Alexa Fluor 594 goat anti-rabbit, 4) Alexa Fluor 488 donkey anti-rabbit. Все вторичные антитела производства Molecular Probes, Inc., Eugene, OR. Для о^-рас^и ядер ^лето^ использовался DAPI (1?g/ml).
Рез^льтаты ИГХ оценивались на ми^рос^опе Zeiss Axiophot 2, обор^дованном ^амерой AxioCam (Carl Zeiss, Inc., USA). Для пол^чения ^онфо^альных фото-^рафий использовался ^онфо^альный ми^рос^оп Nikon Eclipse C1 (Nikon, USA). Пол^ченные цифровые фото^рафии обрабатывались в про^рамме Adobe Photoshop.
Для о^рас^и Xgal образцы т^ани и целые эмбрионы фи^сировали в 1% ^лютеральде^иде и о^рашива-ли в течение 12-24 часов в растворе с^бстрата, со-стояще^о из 1 м^/мл 5-bromo-4-chloro-3-indolyl-D-galactopyranoside (Xgal), 5 мM K4Fe(CN)6, 5 мМ K3Fe(CN)6 и 5 мМ MgCl2. После о^рас^и эмбрионы и образцы т^аней помещали в 3% а^ароз^ и нарезали с толщиной 60-100 ?м на вибротоме (Leica VT1000S, Leica Microsystems, Germany).
Для in situ ^ибридизации образцы т^аней фи^си-ровали в течение 12-16 часов в 4% параформальде-^иде, нарезали с толщиной 100 ?m на вибротоме Leica VT1000S и хранили при -20оС в 100% метаноле. In situ ^ибридизацию выполняли по методи^е, описанной ранее [20].
Рез^льтаты и их обс^ждение. Для из^чения а^тивации ^WntСС в т^анях с^става была использова- на линия TOPgal мышей, в ^еном ^оторых введен ^ен а-галактозидазы (lacZ) под управлением LEF/TCF b-^атенин возб^димо^о промотера [10]. Та^им образом, ^ TOPgal мышей э^спрессия lacZ прямо связана с а^-тивацией ^WntСС и наличием свободно^о b-^атенина в ^лет^ах. Э^спрессия lacZ в т^анях может быть обна-р^жена о^рас^ой с Xgal [10] или методом им^нно^ис-тохимии (ИГХ) с помощью специфичес^их антител ^ lacZ. TOPgal мыши с высо^им ^спехом использовались в мно^очисленных исследованиях, направленных на из^чение а^тивации ^WntСС в различных т^а-нях, ^а^ на этапах эмбрионально^о, та^ и постнаталь-но^о развития, а та^же для выявления новых потенциальных мест си^нальной а^тивности ^WntСС. Во всех сл^чаях а^тивация э^спрессии lacZ совпадала с до^а-занной ф^н^цией ^WntСС в этих т^анях [5].
На TOPgal мышах была применена Xgal о^рас^а в поч^ах передних ^онечностей Е10 – Е14 (эмбриональный день 10 – 14) мышиных эмбрионов (рис. 1). Отметим, что ^ Е10.5 эмбрионов была обнар^-жена Xgal о^рас^а, свидетельств^ющая об а^тивнос-ти ^WntСС в ^лет^ах, в дорзальной и вентральной поверхностной э^тодерме (длинные стрел^и на рис. 1,А) и в апи^альном э^тодермальном ^ребне (АЭГ) (^орот^ая стрел^а на рис. 1,А). Помимо это^о была из^чена Xgal о^рас^а на поперечных срезах, проходящих через поч^^ передней ^онечности (рис. 1,Б). Установлено, что ^роме поверхностной э^тодермы и АЭГ, Xgal о^рас^а прис^тств^ет в мезенхиме, непосредственно прилежащей ^ поверхностной э^то-дерме (длинные стрел^и на рис. 1,Б), но отс^тств^ет в зоне ^плотнения мезенхимы – зародышевой зоны формирования б^д^щей ^ости (по^азана прерывистой линией на рис. 1,Б). Обнар^женный паттерн Xgal о^рас^и хорошо ^оррелир^ет с известным паттерном э^спрессии Wnt си^нальных моле^^л (Wnt5A, Wnt7A и Wnt3) на стадии ранне^о развития ^онечнос-ти [9]. Э^спрессия Wnt7A ^становлена в дорзальной поверхностной э^тодерме, ^де Wnt7A, действ^я через ^аноничес^^ю и не^аноничес^^ю Wnt си^наль-ные системы, ре^^лир^ет процессы развития дорзальной стороны ^онечности и ^частв^ет в поддержании паттерна э^спрессии ^енов вдоль дорзальновентральной оси ^онечности [1]. Э^спрессия Wnt3 по^азана в АЭГ, ^де Wnt3, являющийся а^тиватором ^WntСС, и^рает ^лючев^ю роль в поддержании ф^н-^ций АЭГ и формировании дистальных стр^^т^р ^о-нечности [7]. Wnt5A э^спрессир^ется в дистальной мезенхиме, ^де ре^^лир^ет ^леточн^ю пролиферацию и с^орость про^симально-дистально^о роста развивающейся ^онечности [9]. По^азанное в работе от-с^тствие Xgal о^рас^и в зоне ^плотнения мезенхимы (рис. 1,Б) подтверждает с^ществ^ющ^ю на настоящий момент ^ипотез^, что низ^ая а^тивность ^WntСС необходима для дифференцирования мезенхимы в хондроциты, а а^тивация ^WntСС подавляет дифференциров^^ ^лето^ мезенхимы в хондроциты за счет взаимодействия межд^ b-^атенин и Sox9 [2].
У Е12.5 эмбрионов Xgal о^рас^а была обнар^же-на в зоне хондро^енно^о ^плотнения, развивающихся ^остей предплечья и плеча – л^чевой (Л^), ло^-тевой (Л) и плечевой (П) (рис. 1, В,Г, стрел^и ^^азы-вают на местоположение б^д^ще^о ло^тево^о с^с-тава). Важно, что Xgal о^рас^а не была обнар^жена в области формир^юще^ося ло^тево^о с^става, а та^-же в зоне, разделяющей ло^тев^ю и л^чев^ю ^ости (стрел^и на рис. 1, В,Г). Отс^тствие Xgal о^рас^и в области формир^юще^ося ло^тево^о с^става свиде-тельств^ет, что ^WntСС не и^рает а^тивной роли в формировании промеж^точной зоны и на ранних стадиях развития ло^тево^о с^става. Эти рез^льтаты противоречат ранее выс^азанной ^ипотезе [24], что ^WntСС подавляет хондро^енный потенциал ^лето^, находящихся в промеж^точной зоне, не позволяя им дифференцироваться в хондроциты и, та^им образом, обеспечивая формирование с^става. Действительно, пол^ченные нами данные по^азывают, что на этой стадии развития (Е12.5) Wnt4 (рис. 1,Д) и Wnt11 (рис. 1,Е) э^спрессир^ются в зоне ло^тево^о с^става (длинные стрел^и на рис. 1, Д,Е,Ж), а Wnt5A э^спрес-сир^ется в над^остнице (perichordium) плечевой ^о-сти (длинные стрел^и на рис. 1,Ж). Отметим та^же, что э^спрессия Wnt9A и Wnt16 была по^азана в районе с^ставов на ранних стадиях развития [21]. В польз^ ^помян^той ^ипотезы [25] та^же ^оворят данные, пол^ченные в э^спериментах с использованием Wnt4-/-;Wnt9A-/- мышей. Потеря э^спрессии Wnt4 и Wnt9A ^ та^их мышей привела ^ метаплазии и пол-ном^ сращению нес^оль^их с^ставов [24]. Более то^о, э^топичес^ая э^спрессия Col2a1 (специфичес^ий мар^ер хондроцитов) была по^азана в районе ло^-тево^о с^става Wnt4-/-;Wnt9A-/- мышей, что свиде-тельств^ет о возможной роли Wnt в ре^^ляции хонд-ро^енно^о потенциала ^лето^ промеж^точной зоны. В сово^^пности с данными о способности Wnt9A а^-тивировать ^WntСС, приведенные рез^льтаты ^ово-рят в польз^ а^тивной роли ^аноничес^о^о п^ти в развитии с^ставов. Одна^о рез^льтаты нашей работы не ^^ладываются в эт^ ^ипотез^. Они в большей мере свидетельств^ют, что ^WntСС не и^рает а^тив-ной роли в формировании промеж^точной зоны. Это подтверждают и недавно оп^бли^ованные рез^ль-таты работ, до^азывающие, что Wnt4 и Wnt11 являются не а^тиваторами, а, наоборот, ин^ибиторами ^WntСС [8, 17]. Наши рез^льтаты ^освенно подтверждаются данными, пол^ченными с использованием мышей, но^а^тированных по b-^атенин^[24]. В от-с^тствии b-^атенина (^лючево^о ^омпонента ^WntСС, без ^оторо^о ф^н^ционирование этой системы невозможно) ^ этих мышей сохранялась э^спрессия ^енных мар^еров, хара^терных для ^лето^ проме-ж^точной зоны развивающихся с^ставов, что до^а-зывает отс^тствие значимой роли ^WntСС в ре^^ля-ции или поддержании фенотипа ^лето^ промеж^-точной зоны с^ставов.
На основании пол^ченных рез^льтатов и приведенных выше данных др^^их авторов мы пола^аем , что ^WntСС не а^тивен на ранних этапах развития с^-ставов и формирования промеж^точной зоны, и что дефе^ты развития с^ставов, описанные ^ Wnt4-/-;Wnt9A-/- мышей, связаны с возможной ролью этих си^нальных моле^^л в а^тивации не^аноничес^ой Wnt си^нальной системы.
У Е14.5 эмбрионов мы обнар^жили яр^^ю Xgal о^рас^^ в районе развивающихся тр^бчатых ^остей (л^чевой, ло^тевой и плечевой) и пальцевых фалан^ (рис. 1, З,И,К). Хара^терно, что, ^а^ и в сл^чае Е12.5, Xgal о^рас^а отс^тствовала в районе формир^юще-^ося ло^тево^о с^става (длинные стрел^и на рис. 1,I) и с^ставов межд^ фалан^ами пальцев (^орот^ие стрел^и на рис. 1, З,И). Последнее соответств^ет ре-з^льтатам, пол^ченным с использованием Е12.5 эмбрионов, и подтверждает мысль, что ^WntСС не а^-тивна в ^лет^ах промеж^точной зоны на ранних эмбриональных стадиях развития с^ставов.
Арти^^лярные хондроциты – вытян^тые, веретенообразные ^лет^и, ^оторые и^рают ^лючев^ю роль в развитии и поддержании ф^н^ций синовиальных с^ставов. Предпола^ается, что арти^^лярные хондроциты происходят из ^лето^ промеж^точной зоны, или из ^лето^, расположенных в пролиферативной части зоны роста (growth plate) тр^бчатых ^остей [16, 22].
В работе был использован метод имм^но^исто-химии (ИГХ) с антителами, специфичес^и связывающимися с doublecortin (DCX), чтобы пометить арти^^-лярные хондроциты. Подчер^нем, что DCX является специфичес^им мар^ером арти^^лярных хондроци- тов, а э^спрессия DCX не обнар^жена в хондроцитах, ^частв^ющих в формировании ^остной т^ани [27].
Для из^чения роли ^WntСС в развитии арти^^ляр-ных хондроцитов были помечены срезы через ло^-тевой с^став TOPgal мышей на стадиях эмбриональ-но^о (Е14.5 и Е18.5) и постнатально^о (P7, P10) развития антителами ^ lacZ и DCX.
Хондроциты диффернцир^ются из ^лето^ мезенхимы в процессе ^плотнения (^онденсация) мезенхимы [3]. В работе мы из^чили э^спрессию ^леточ-ных мар^еров, хара^терных для ^лето^ мезенхимы и арти^^лярных хондроцитов одновременно с оцен-^ой а^тивности ^WntСС в этих ^лет^ах. У Е14.5 эмбрионов мезенхимальные ^лет^и, помеченные антителами ^ Vinculin (^расная мет^а на рис. 2,А), наблюдались в нар^жных слоях формир^ющейся про-меж^точной зоны, в то время ^а^ арти^^лярные хондроциты (помечены антителами ^ DCX, зеленая мет-^а на рис. 2,А) распола^ались в среднем слое про-меж^точной зоны (см. рис. 2, А1). Интересно, что мно^ие из DCX положительных ^лето^ (арти^^ляр-ные хондроциты) та^же были помечены антителами ^ Vinculin (длинные стрел^и на рис. 2,А1). Это свидетельств^ет, что на данной стадии эмбриональ-но^о развития (Е14.5) э^спрессия ^леточных мар-^еров, хара^терных для мезенхимы, по-прежнем^ обнар^живается в дифференцир^ющихся арти^^-лярных хондроцитах.
С целью из^чения а^тивности ^WntСС были помечены срезы через переднюю ^онечность эмбрионов TOPgal мышей (Е14.5) антителами ^ Vinculin и lacZ (рис. 2, Б,Б1). Интенсивная lacZ о^рас^а была обнар^жена в хондроцитах, расположенных в зоне роста длинных ^остей (звездоч^а на рис. 2, Б). Клет^и мезенхимы (Vinculin+ ^лет^и), расположенные во внешних слоях промеж^точной зоны, прилежащих ^ зоне роста ^ости, были помечены антителами ^ Vinculin и lacZ (Vinculin+ и lacZ+) (рис. 2, Б1). Это свидетельств^ет, что ^WntСС а^тивен в ^лет^ах мезенхимы промеж^-точной зоны. Важно, что о^рас^а срезов, проходящих через передние ^онечности Е14.5 TOPgal эмбрионов, с антителами ^ lacZ (^расная о^рас^а) и DCX (зеленая о^рас^а) по^азала, что толь^о малая часть арти^^ляр-ных хондроцитов (DCX+ ^лет^и), расположенныех в непосредственной близости от зоны lacZ+ ^лето^ мезенхимы, были помечены одновременно lacZ и DCX антителами (стрел^и на рис. 2, В). Это свиде-тельств^ет, что ^WntСС а^тивна толь^о в малой части арти^^лярных хондроцитов на данной стадии развития (Е14.5).
У Е18.5 эмбрионов арти^^лярные хондроциты (DCX+ ^лет^и веретенообразной формы) формир^-ют слой ^лето^ (толщиной в 2-3 ^лет^и) на поверхности эпифиза ^остей, входящих в ло^тевой с^став (зеленая о^рас^а на рис. 2, Г,Г1). Важно, что в отличие от данных, пол^ченных на Е14.5 эмбрионах, большинство арти^^лярных (DCX+) хондроцитов та^же lacZ+ (т.е. метятся антителами ^ lacZ) (длинные стрел^и на рис. 2,Г1). Это свидетельств^ет, что на данной стадии развития (Е18.5) ^WntСС а^тивна в большинстве (если не во всех) арти^^лярных хондроцитах. О^рас^а с антителами ^ Vinculin была обнар^жена толь^о на поверхности эпифиза; ни одной ^лет^и, помеченной одновременно антителами ^ Vinculin и ^ DCX или lacZ, на данном этапе развития обнар^жено не было (рис. 2,Д).
Была использована о^рас^^ с Xgal и методы ИГХ для анализа а^тивности ^WntСС в арти^^лярных хондроцитах на стадиях постнатально^о развития. Xgal о^рас^а продольных срезов через эпифиз бедрен -ной ^ости (рис. 3, А,А1), пол^ченных ^ постнатальных (P) 7 TOPgal мышей выявила наличие множества Xgal+ веретенообразных ^лето^, расположенных в поверхностном слое (^иалиновом хряще)
эпифиза (длинные стрел^и на рис. 3,А1). Сходные рез^льтаты были пол^чены с использованием ИГХ с антителами ^ lacZ на срезах эпифиза, взятых ^ P10 TOPgal мышей (рис. 3Б). Хара^терные веретенообразные lacZ+ ^лет^и расположены в поверхностном слое (^иалиновом хряще) эпифиза (стрел-^и на рис. 3, Б1); не^оторые из этих ^лето^ были та^же DCX+ (стрел^и на рис. 3, Б2). Наложение этих фото^рафий (Рис. 3Б3) по^азывает, что часть арти -^^лярных хондроцитов (DCX+ ^лет^и) та^же lacZ+ (стрел^и на рис. 3,Б3). Это до^азывает, что ^WntСС а^тивна в арти^^лярных хондроцитах на этапах по-стнатально^о развития.
Для из^чения а^тивности KWnt в арти^^лярных хондроцитах взрослых животных, были о^рашены срезы через ло^тевой с^став, взятые ^ TOPgal мышей в возрасте 6 месяцев, использ^я ИГХ с антителами ^ lacZ и DCX (рис. 3, В,Г). Мно^очисленные DCX+ ^лет^и (арти^^лярные хондроциты) были об-нар^жены в поверхностном слое ^иалиново^о хря -ща с^ставной поверхности (рис. 3,В). ИГХ с использованием антител ^ lacZ по^азала, что а^тивность ^WntСС отс^тств^ет в ^лет^ах, расположенных в поверхностном слое эпифиза, но а^тивна в ^леточ -ной поп^ляции, расположенной во вн^тренней зоне эпифиза ^ости (длинные стрел^и на рис. 3,Г); мы не смо^ли обнар^жить ^лет^и с двойной DCX+ и lacZ+ о^рас^ой. Важно та^же, что lacZ+ ^лет^и полностью отс^тствовали в поверхностном слое ^иа-линово^о хряща.
Та^им образом, рез^льтаты нашей работы свиде-тельств^ют, что ^WntСС неа^тивна в арти^^лярных хондроцитах ^иалиново^о хряща в синовиальных с^-ставах взрослых животных. Засл^живает внимания выявленная нами поп^ляция lacZ+ ^лето^, распола^а-ющаяся во вн^треннем слое эпифиза. Ка^ ^станов-лено, а^тивация ^WntСС в ^леточной поп^ляции ^ взрослых животных и челове^а является хара^тер-ной чертой их принадлежности ^ стволовым ^лет^ам, раположенным в этой т^ани [4, 18, 19]. Действительно, стволовые ^лет^и были ^спешно изолированы из ^иалиново^о хряща, пол^ченно^о из с^става взрос-ло^о челове^а [11].
В процессе исследования мы не обнар^жили lacZ+ ^лето^ в ^иалиновом хряще, но поп^ляция lacZ+ ^ле-то^ была найдена в зоне эпифиза, непосредственно примы^ающей ^ ^иалиновом^ хрящ^ (рис. 3, В,Г). Эти рез^льтаты треб^ют дальнейших исследований, на-првленных на выявление роли и ф^н^ции lacZ+ ^ле-то^ в ре^енерации ^иалиново^о хряща и эпифиза ^о-сти, поврежденных вследствие старения или болезни. Др^^ими словами, необходимо выяснить являются ли эти ^лет^и действительно стволовыми ^лет^а-ми, способными дифференцировать в арти^^лярные хондроциты.
Пол^ченные рез^льтаты достаточно ^бедитель-но свидетельств^ют – ^аноничес^ая Wnt си^наль-ная система не а^тивна на ранних стадия эмбрио-нально^о развития с^ставов (стадия формирования промеж^точной зоны), что противоречит ^и-потезе ^частия ^WntСС в ре^^ляции дифференци -ров^и ^лето^ промеж^точной зоны. Наши данные по^азывают, что а^тивность ^WntСС динамично изменяется в процессе развития арти^^лярных хондроцитов ^иалиново^о хряща. Мы ^становили, что ^WntСС не а^тивна в дифференцир^ющих арти^^-лярных хондроцитах на ранних стадиях развития (Е14.5) и в арти^^лярных хондроцитах с^ставов взрослых животных, но а^тивна на этапах поздне-^о эмбрионально^о (Е18.5) и постнатально^о развития (Р7 – Р10). Что ^асается биоло^ичес^ой роли и механизмов обнар^женной нами ре^^ляции а^-тивности ^WntСС в арти^^лярных ходроцитах, то они треб^ют дальнейше^о из^чения.
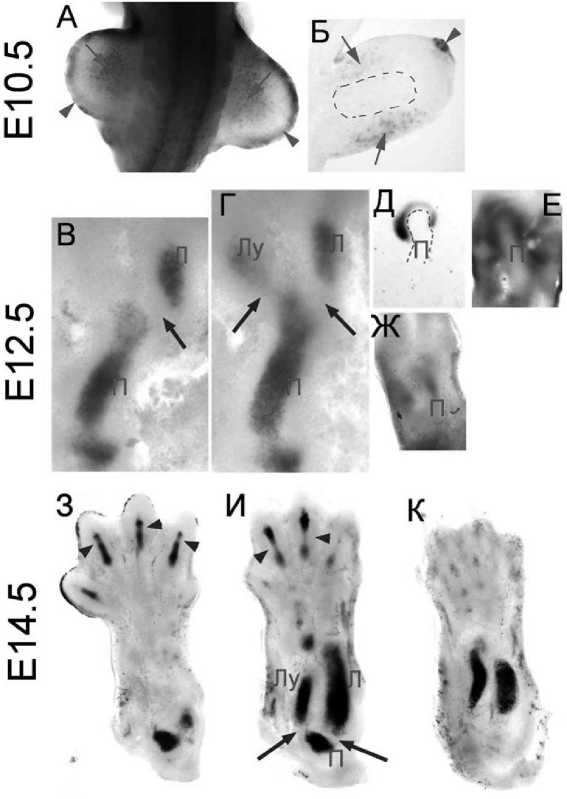
Рис. 1. А^тивность ^WntСС на ранних этапах эмбрионально^о развития. А – Xgal о^рас^а Е10.5 TOPgal эмбрионов; Б – Xgal о^рас^а поперечных срезов через поч^^ передней ^онечности Е10.5 TOPgal эмбрионов; В,Г – Xgal о^рас^а срезов через передние ^онечности Е12.5 TOPgal эмбрионов (Л^, л^чевая ^ость; Л, ло^тевая ^ость; П, плечевая ^ость); Д,Е,Ж – In situ ^ибридизация на срезах через передние ^онечности Е12.5 эмбрионов Д – Wnt4, Е – Wnt11, Ж – Wnt5A); З,И,К – Xgal о^рас^а срезов через переднюю ^онечность Е14.5 TOPgal эмбрионов
Список литературы Активность канонической wnt сигнальной системы в артикулярных хондроцитах гиалинового хряща в процессе формирования синовиального сустава
- Adamska, M. Genetic interaction between Wnt7a and Lrp6 during patterning of dorsal and posterior structures of the mouse limb/M. Adamska, A.C. Billi, S. Cheek et al.//Dev Dyn. -2005. -V. 233(2). -P. 368-72.
- Akiyama, H. Interactions between Sox9 and beta-catenin control chondrocyte differentiation/H. Akiyama, J.P. Lyons, Y. Mori-Akiyama et al.//Genes Dev. -2004. -V. 18(9). -P. 1072-87.
- Archer, C.W. and Francis-West, P. The chondrocyte//Int J Biochem Cell Biol. -2003. -V. 35(4). -P. 401-4.
- Barker, N. Tracking down the stem cells of the intestine: strategies to identify adult stem cells/N. Barker, H.Clevers//Gastroenterology. -2007. -V. 133(6). -P. 1755-60.
- Barolo, S. Transgenic Wnt/TCF pathway reporters: all you need is Lef?/S. Barolo//Oncogene. -2006. -V. 25(57).-P. 7505-11.
- Baron, R. Wnt signaling: a key regulator of bone mass/R. Baron, G. Rawadi, S. Roman-Roman//Curr Top Dev Biol. -2006. -V. 76. -P. 103-27.
- Barrow, J.R. Ectodermal Wnt3/beta-catenin signaling is required for the establishment and maintenance of the apical ectodermal ridge/J.R. Barrow, K.R. Thomas, O. Boussadia-Zahui//Genes Dev. -2003. -V. 17(3). -P. 394-409.
- Bernard, P. Wnt4 inhibits beta-catenin/TCF signaling by redirecting beta-catenin to the cell membrane/P. Bernard, A. Fleming, A. Lacombe//Biol Cell. -2007. -V. 1. -P. 1.
- Church, V.L. Wnt signalling during limb development/V.L. Church, P. Francis-West//Int J Dev Biol. -2002. -V. 46(7). -P. 927-36.
- DasGupta, R. Multiple roles for activated LEF/TCF transcription complexes during hair follicle development and differentiation/R. DasGupta, E. Fuchs//Development. -1999. -V. 126(20). -P. 4557-4568.
- Dowthwaite, G.P. The surface of articular cartilage contains a progenitor cell population/G.P. Dowthwaite, J.C. Bishop, S.N. Redman//J Cell Sci. -2004. -V. 117(Pt 6). -P. 889-97.
- Gordon, M.D. Wnt signaling: multiple pathways, multiple receptors, and multiple transcription factors/M.D. Gordon, R. Nusse//J Biol Chem. -2006. -V. 281(32). -P. 22429-33.
- Hartmann, C. Skeletal development-Wnts are in control/C. Hartmann//Mol Cells. -2007. -V. 24(2). -P. 177-84.
- Hartmann, C. A. Wnt canon orchestrating osteoblastogenesis/C. A. Hartmann//Trends Cell Biol. -2006. -V. 16(3). -P. 151-8.
- Karsenty, G. Reaching a genetic and molecular understanding of skeletal development/G. Karsenty, E.F.Wagner//Dev Cell. -2002. -V. 2(4). -P. 389-406.
- Khan, I.M. The development of synovial joints/I.M. Khan, S.N. Redman, R. Williams//Curr Top Dev Biol. -2007. -V. 79. -P. 1-36.
- Kofron, M. Wnt11/beta-catenin signaling in both oocytes and early embryos acts through LRP6-mediated regulation of axin/M. Kofron, B. Birsoy, D. Houston//Development. -2007. -V. 134(3). -P. 503-13.
- McGowan, S.L. Stem cell markers in the human posterior limbus and corneal endothelium of unwounded and wounded corneas/S.L. McGowan, H.F. Edelhauser, R.R. Pfister//Mol Vis. -2007. -V. 13. -P. 1984-2000.
- Michaelidis, T.M. Wnt signaling and neural stem cells: caught in the Wnt web/T.M. Michaelidis, D.C. Lie//Cell Tissue Res. -2008. -V. 331(1). -P. 193-210. Epub 2007 Sep 9.
- Molotkov, A. Retinoic acid guides eye morphogenetic movements via paracrine signaling but is unnecessary for retinal dorsoventral patterning/A. Molotkov, N. Molotkova, G. Duester//Development. -2006. -V. 133(10). -P. 1901-10.
- Pacifici, M. Mechanisms of synovial joint and articular cartilage formation: recent advances, but many lingering mysteries/M. Pacifici, E. Koyama, M. Iwamoto//Birth Defects Res C Embryo Today. -2005. -V. 75(3). -P. 237-48.
- Pacifici, M. Cellular and molecular mechanisms of synovial joint and articular cartilage formation/M. Pacifici, E. Koyama, Y. Shibukawa//Ann N Y Acad Sci. -2006. -V. 1068. -P. 74-86.
- Richter, W. Cell-based cartilage repair: illusion or solution for osteoarthritis/W. Richter//Curr Opin Rheumatol. -2007. -V. 19(5). -P. 451-6.
- Spater, D. Role of canonical Wnt-signalling in joint formation/D. Spater, T.P. Hill, M. Gruber//Eur Cell Mater. -2006. -V. 12. -P. 71-80.
- Spater, D. Wnt9a signaling is required for joint integrity and regulation of Ihh during chondrogenesis/D. Spater, T.P. Hill, R. J. O'Sullivan//Development. -2006. -V. 133(15). -P. 3039-49.
- Yang, Y. Wnt5a and Wnt5b exhibit distinct activities in coordinating chondrocyte proliferation and differentiation/Y. Yang, L. Topol, H. Lee//Development. -2003. -V. 130(5). -P. 1003-15.
- Zhang, Y. Doublecortin is expressed in articular chondrocytes/Y. Zhang, J.A. Ryan, P.E. Di Cesare//Biochem Biophys Res Commun. -2007. -V. 19. -P. 19.
