Активность компонентов антиоксидантной защиты Raphanus sativus L. при выращивании на почве, загрязненной сульфатами свинца и кадмия

Автор: Еремченко О.З., Кусакина М.Г., Голева Т.Н.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 1, 2014 года.
Бесплатный доступ
Исследована активность антиоксидантной защиты в редисе Raphanus sativus L. var. radicula Pers., произрастающем в условиях загрязнения почвы сульфатами свинца и кадмия. Редокс-ак-тивность в листьях редиса усиливалась с ростом дозы загрязнения. Повышенная активность ка-талазы и пероксидаз в растениях отмечена на вариантах загрязнения почвы сульфатом кадмия.
Загрязнение почв, тяжелые металлы, активные формы кислорода, пероксид водорода, пероксидазы, каталаза, аскорбиновая кислота, редокс-активность
Короткий адрес: https://sciup.org/147204662
IDR: 147204662 | УДК: 581.632.1
The influence of soil pollution by pbso4 and cdso4 on the antioxidant system of Raphanus sativus L
Results of researches of the content of antioxidants in the plants of Raphanus sativus L. in the conditions of soil pollution PbSO4 and CdSO4 on the antioxidant system are presented. It is shown, that alterations in the plants antioxidant protection system are dependent on different levels of heavy metals in soil.
Текст научной статьи Активность компонентов антиоксидантной защиты Raphanus sativus L. при выращивании на почве, загрязненной сульфатами свинца и кадмия
Повышенное содержание тяжелых металлов в почвенной среде приводит к ингибированию роста, изменению интенсивности и направленности многих метаболических процессов в клетках растений. Известно, что тяжелые металлы при низкой концентрации выполняют роль важнейших микроэлементов, необходимых для нормального роста и развития растений. Однако остается неизвестной физиологическая роль Cd и Pb. При загрязнении растения поглощают эти элементы, избирательно аккумулируют в корнях и надземных вегетативных органах. При избыточном накоплении в растительных клетках свинца и кадмия нарушается гомеостаз и подавляются многие процессы метаболизма [Sanita di Toppi, Gabrielli, 1991; Серегин, Иванов, 2001]. Ингибирующее влияние Cd и Pb на активность большинства ферментов обусловлено как прямым связыванием с функциональными SH-группами белков, так и с нарушением ионного баланса за счет конкурентного снижения поглощения и нарушения транспорта Zn, Fe, Cu и др. [Титов, Таланова, Казнина, 2011].
Под влиянием неблагоприятных условий среды и, в частности, техногенного загрязнения в клетках растений происходит развитие окислительного стресса с избыточным образованием активных форм кислорода (АФК) [Зенков, Лапкин, Меньшикова, 2001; Чиркова, 2002; Га-рифзянов, 2012; Сигнальная роль ..., 2012]. К основным механизмам устойчивости растений к дей- ствию поллютантов относят активацию ферментов антиоксидантной защиты (супероксиддисмутаза, пероксидазы, каталаза и др.), а также накопление редокс-соединений (аскорбиновая кислота, глутатион, каротиноиды и др.), прерывающих цепи избыточного образования свободных радикалов [Fridovich, 1986, Кения, Лукаш, Гуськов, 1993; Mittler, 2002; Прадедова, Ишеева, Саляев, 2011].
Повышение активности компонентов антиоксидантной системы защиты может определять уровень адаптационных способностей растений к действию антропогенных загрязнителей. Однако особенности функционирования антиоксидантной системы растений в условиях загрязнения среды тяжелыми металлами изучены недостаточно. В связи с этим целью работы явилось исследование активности некоторых компонентов антиоксидантной защиты редиса посевного при выращивании на почве, загрязненной сульфатами свинца и кадмия.
Объект и методы исследования
Объектом исследований был редис посевной ( Raphanus sativus L. var. radicula Pers., Brassicaceae ) сорта Жара. Растения выращивали в условиях модельного эксперимента в течение 30 дней на агро-дерново-подзолистой почве на фоне внесения сульфатов свинца (1, 5 и 10 ПДК, при ПДК=100 мг/кг почвы) и кадмия (1, 50, 100 ПДК, при ПДК=3 мг/кг
почвы). Контролем служили растения, выращенные на незагрязненной почве. Повторность опытов – трехкратная. Определение массы и высоты надземной части растений проводили в 20-кратной повторности. Биохимические показатели устанавливали в 3-кратной повторности. Общее содержание перекисей определяли ферротиоционатным методом [Лу-каткин, 2002]; активность каталазы – газометрическим методом [Методы ..., 1987]; активность пероксидаз – по методу Бояркина [Методы ..., 1987]. Содержание аскорбиновой кислоты и общую редокс-активность изучали методом Петта – Прокошева [Практикум …, 1972].
Полученные результаты статистически обработаны с помощью компьютерной программы «Stadia 8.0»; были использованы дисперсионный и регрессионный анализы.
Результаты и их обсуждение
Загрязнение почвы солью свинца не оказало достоверного воздействия на высоту растений (рис. 1). В ряде работ показана низкая проникающая способность свинца в растительный организм [Ильин, 1991; Серегин, Иванов, 1997; Еремченко, Чудинова, 2012]. При невысоких концентрациях тяжелых металлов в клетке изменение физиологобиохимических реакций носит неповреждающий и обратимый характер. Согласованность метаболических процессов сохраняется, а иногда даже наблюдается интенсификация некоторых из них [Титов, Таланова, Казнина, 2011].
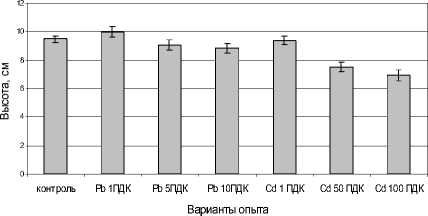
Рис. 1 . Влияние сульфатов свинца и кадмия на высоту растений редиса, см
Достоверное снижение высоты растений прослежено по вариантам Cd 50 и 100 ПДК (рис. 1). Торможение роста надземной части растений связывают как с непосредственным влиянием кадмия на клеточное деление, так и с нарушением общего метаболизма растений; отмечено уменьшение фотосинтезирующей поверхности, замедление скорости фотосинтеза, нарушение минерального питания, а также изменение гормонального баланса, в частности, увеличение количества абсцизовой кислоты [Иванов, Быстрова, Серегин, 2003; Титов, Казнина, Таланова, 2012].
Токсическое воздействие на массу растений оказало загрязнение Pb и Cd (табл. 2), однако исходя из внесенных доз солей, эффект действия кадмия проявлялся на порядок сильнее (рис. 2).
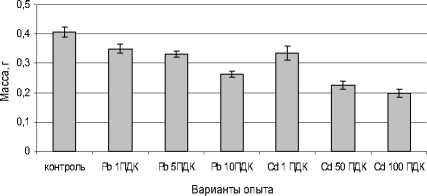
Рис. 2 . Влияние сульфатов свинца и кадмия на сырую массу надземной части растений, г
Как известно, поллютанты (в том числе тяжелые металлы) инициируют развитие в клетках живых организмов окислительного стресса, обязательным условием возникновения которого является избыточное образование активных форм кислорода, таких как супероксид-радикал (анион-ради-кал) О 2 •-, гидроперекисный радикал НО 2 •, гидрок-сил-радикал НО•, перекись водорода Н 2 О 2 , синглетный кислород 1О 2 . В результате повышенной генерации АФК в клетках происходят окисление липидов, углеводов, белков, повреждение ДНК и РНК, дезорганизация цитоскелета. Перекись водорода является относительно стабильным соединением (время жизни около 1 мс), способным диффундировать от места образования [Гарифзянов, 2012]. Её генерация может служить одним из биохимических показателей состояния растений в условиях нарушения внутриклеточных процессов.
Дисперсионный анализ подтвердил достоверное влияние фактора загрязнения почвы на содержание перекисей в листьях растений. В то же время количество перекисей зависело не только от их продуцирования, но и от эффективности работы антиоксидантной системы. На фоне действия соли свинца содержание перекисей в листьях растений повышено на вариантах 5 и 10 ПДК по сравнению с контрольными растениями (рис. 3).
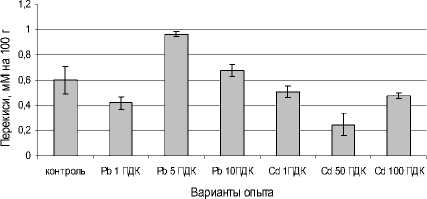
Рис. 3 . Влияние сульфатов свинца и кадмия на содержание перекисей в листьях редиса, мМ на 100 г сырой массы
Возможно, в этом случае накопление Н2О2 в клетках при стрессе связано с его сигнальной ро- лью, обеспечивающей включение защитных механизмов [Гамалей, Клюбин, 1996]. В вариантах с более токсичным кадмием содержание перекисей ниже контроля, по-видимому, из-за повышенной активности систем антиоксидантной защиты растений.
Избыток пероксида водорода, возникающий при действии стрессовых факторов, элиминируется каталазой и пероксидазами [Мирошниченко, 1992; Мерзляк, 1999; Роль систем …, 2008]. Наличие нескольких ферментов, выполняющих одну и ту же каталитическую функцию, расширяет адаптационный потенциал растений [Половникова, Воскресенская, 2008]. Каталаза преобразует перекись водорода в воду и молекулярный кислород. Она участвует в аскорбат-глутатионовом цикле, регулирующем окислительно-восстановительное равновесие в клетке. Поскольку каталаза и пероксидаза предотвращают избыточную аккумуляцию АФК и снижают вероятность развития окислительного стресса, изменения в их активности рассматривают как ранний и чувствительный индикатор токсичности тяжелых металлов [Иванов, Савочкин, Кузнецов, 2012].
В научной литературе отмечается разная реакция растений на воздействие поллютантов. В опытах М.Г. Половниковой и О.Л. Воскресенской [2008] при усилении загрязнения среды поллютантами наблюдалось снижение активности каталазы, особенно у менее устойчивого к антропогенному воздействию клевера лугового. В.О. Миттова и А.У. Игамбердиев [2000] считают, что в стрессовых условиях общая активность каталазы снижается, но увеличивается активность изоформ каталазы, обладающих пероксидазной активностью. Более высокий уровень активности каталазы в стрессовых условиях отмечен в исследованиях Т.В. Чирковой [2002] и П. Ахмада, К.А. Джамила, С. Шармы [2010].
Сульфат свинца в дозе Pb 1 ПДК не оказал достоверного влияния на активность каталазы в листьях растений (рис. 4). При повышенном загрязнении свинцом и наименьшей дозе кадмия зафиксировано снижение активности этого фермента. Вероятно, в адаптации растений на этом фоне имеют значения другие антиоксиданты. Под влиянием Cd 50 ПДК отмечено усиление активности каталазы в листьях растений в 1.5 раза по сравнению с контролем.
В результате поиска связей между биохимическими показателями установлена обратная экспоненциальная зависимость между общим количеством перекисей и активностью каталазы: y =e0,25 – 0.3х ; уровень значимости нулевой гипотезы P=0.0004; критерий Фишера F=11.3. Следовательно, чем выше активность каталазы, тем ниже содержание перекисей в листьях
В реализации адаптационного потенциала особая роль отводится пероксидазам (аскорбат-перок-сидаза, глутатион-пероксидаза, гваякол-пероксида-за и др.) – полифункциональной ферментной системе, которая способна реагировать на широкий спектр факторов, приводящих к нарушению обмена веществ у растений. Пероксидазы способны восстанавливать перекись водорода до воды, окисляя при этом различные соединения [Лукаткин, 2002].
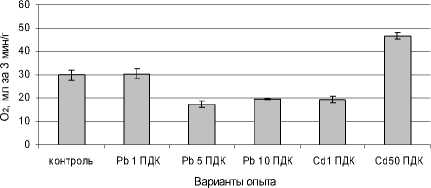
Рис. 4 . Влияние сульфатов свинца и кадмия на активность каталазы в листьях редиса, мл О 2 за 3 мин на 1 г сырой массы
Проведенные опыты показали, что внесение в почву сульфата свинца не сопровождалось усилением пероксидазной активности растений редиса (рис. 5). В варианте с внесением кадмия наблюдалось статистически достоверное увеличение активности пероксидаз по сравнению с контролем. Усиление активности пероксидаз при действии стресс-факторов и, в частности, тяжелых металлов, может быть связано с изменениями их компонентного состава. Ферменты находятся в клетке как в свободном, так и в связанном состоянии, а под влиянием стресса происходит увеличение доли свободных форм за счет освобождения из связанных форм [Рогожин, 2004].
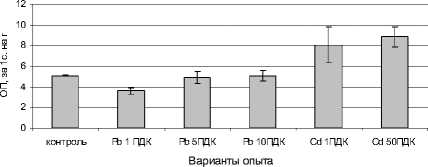
Рис. 5 . Влияние сульфатов свинца и кадмия на активность пероксидаз в листьях редиса, оптическая плотность (ОП) за 1 с на 1 г сырой массы
Повышение пероксидазной и каталазной активности в листьях редиса под влиянием CdSO 4 приводит к сохранению прооксидантно – оксидантного равновесия в биологических мембранах и свидетельствует об активации защитных механизмов от окислительного стресса, вызванного действием тяжелых металлов.
В целом следует отметить усиленные признаки развития окислительного стресса в редисе под влиянием соли кадмия. Кадмий обладает большой подвижностью в почве в широком диапазоне рН и способностью замещать в клеточном метаболизме такой важный биоэлемент, как цинк [Серегин, Иванов, 2001]. Ионы Cd чрезвычайно легко поглощаются растениями [Кабата-Пендиас, Пендиас, 1989], а Pb медленнее других тяжелых металлов поступает в растения и транспортируется в надземные органы [Серегин, Иванов, 1997]. Различия в степени ингибирующего действия исследуемых металлов могут быть связаны с эффективностью физиологических барьеров при поступлении микроэлементов в растение, с задержкой металлов в тканях корня или удалением их в вакуоли [Прасад, 2003].
Высокие параметры окислительно-восстановительных процессов в растениях являются показателем мобилизации защитных свойств организма. При окислительном стрессе ферментативная защита может дополняться протекторным действием низкомолекулярных антиоксидантов, таких как аскорбиновая кислота и глутатион, прерывающих цепь свободнорадикального окисления [Кения, Лукаш, Гуськов, 1993; Чиркова, 2002; Половникова, Воскресенская, 2008]. Аскорбиновая кислота реагирует с АФК, окисляясь при этом до моно - и де-гидроаскорбиновой кислоты.
В растениях редиса с увеличением загрязненности сульфатом свинца и кадмия повышалось содержание аскорбиновой кислоты, способной к восстановительным реакциям (рис. 6). Самый высокий уровень содержания аскорбиновой кислоты установлен при максимальных дозах тяжелых металлов – в 2 раза больше, чем в контроле. Накопление аскорбиновой кислоты может быть следствием интенсивности ее образования и/или освобождения из связанного состояния [Кения, Лукаш, Гуськов, 1993].
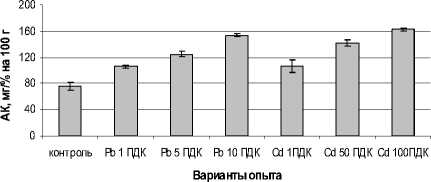
Рис. 6 . Влияние сульфатов свинца и кадмия на содержание свободной аскорбиновой кислоты (АК) в листьях редиса, мг/% сухой массы растений
С увеличением дозы кадмия и свинца повысилась и общая редокс-активность в листьях редиса по сравнению с контролем (рис. 7).
Выводы
-
1. Загрязнение почвы сульфатами свинца и кадмия оказало ингибирующее влияние на массу растений редиса. Достоверное уменьшение высоты растений отмечено в вариантах с солью кадмия, при дозах Cd 50 и 100 ПДК.
-
2. Достоверное накопление перекисей в надземных органах редиса установлено на вариантах загрязнения почвы сульфатом свинца в дозе 5 и 10 ПДК относительно контроля. На вариантах с сульфатом кадмия перекисей в листьях было меньше, чем на контроле.
-
3. При увеличении содержания сульфата кадмия в почве отмечено повышение активности каталазы и пероксидаз в листьях редиса по сравнению с контролем. При загрязнении почвы солью свинца активность каталазы в листьях редиса понижена, не отмечено усиления пероксидазной активности.
-
4. Установлено заметное усиление редокс-ак-тивности в листьях редиса в условиях загрязнения почвы свинцом и кадмием, что, вероятно, является одним из защитно-приспособительных реакций при избыточной генерации активных форм кислорода.
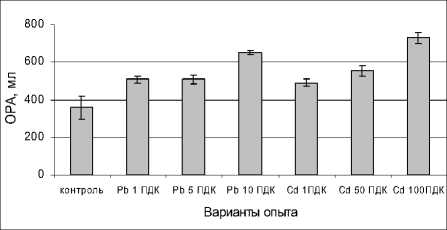
Рис. 7 . Влияние сульфатов свинца и кадмия на общую редокс-активность (ОРА) в листьях редиса, мл 0,001 н. KJO 3 /100г сухой массы
Список литературы Активность компонентов антиоксидантной защиты Raphanus sativus L. при выращивании на почве, загрязненной сульфатами свинца и кадмия
- Ахмад П., Джамил К.А., Шарма С. Влияние солевого стресса на систему антиоксидантной защиты, перекисное окисление липидов, ферментное окисление липидов, ферменты метаболизма пролина и метаболическую активность у двух генотипов шелковицы//Физиология растений. 2010. Т. 57, № 4. С. 547-555
- Гамалей И.А., Клюбин И.В. Перекись водорода как сигнальная молекула//Цитология. 1996. Т. 38. С. 1233-1247
- Гарифзянов А.Р. Образование перекиси водорода и проявление окислительного стресса в листьях древесных растений в условиях промышленного загрязнения//Фундаментальные исследования. Биологические науки. 2012. № 1. С. 151-155
- Еремченко О.З., Чудинова Л.А. Микроэлементный состав почв и растительности заповедной лесостепи в условиях техногенеза//Современные проблемы науки и образования. 2012. № 5. Статья № 279.
- Зенков Н.К., Лапкин В.З., Меньщикова Е.Б. Окислительный стресс: Биохимические и патофизиологические аспекты. М.: Наука, 2001. 343 с
- Иванов В.Б., Быстрова Е.И., Серегин И.В. Сравнение влияния тяжелых металлов на рост корня в связи с проблемой специфичности и избирательности их действия//Физиология растений. 2003. Т. 50, № 3. С. 445-454
- Иванов Ю.В., Савочкин Ю.В., Кузнецов Вл.В. Сосна обыкновенная как модельный объект для изучения механизмов адаптации хвойных к действию тяжелых металлов. 2. Функционирование антиоксидантных ферментов в сеянцах сосны в условиях хронического действия цинка//Физиология растений. 2012. Т. 59, № 1. С. 57-66
- Ильин В.Б. Тяжелые металлы в системе почва -растение. Новосибирск: Наука, 1991. 151 с.
- Кабата-Пендиас А., Пендиас Х. Микроэлементы в почвах и растениях. М.: Мир, 1989. 439 с.
- Кения М.В., Лукаш А.И., Гуськов Е.П. Роль низкомолекулярных антиоксидантов при окислительном стрессе//Успехи современной биологии. 1993. Т. 113, № 4. С. 456-470.
- Лукаткин А.С. Вклад окислительного стресса в развитие холодового повреждения в листьях теплолюбивых растений//Физиология растений. 2002. Т. 49, № 6. С. 878-885.
- Мерзляк М.Н. Активированный кислород и жизнедеятельность растений//Соросовский образовательный журнал. 1999. № 9. С. 20-26.
- Методы биохимического исследования растений/под ред. А.И. Ермакова. Л.: Агропромиздат, 1987. 430 с.
- Мирошниченко О.С. Биогенез, физиологическая роль и свойства каталазы//Биополимеры и клетка. 1992. Т. 8, № 6. С. 7-25.
- Миттова В.О., Игамбердиев У.А. Влияние солевого стресса на дыхательный метаболизм высших растений//Известия РАН. Сер. биол. 2000. №3. С. 322-328.
- Половникова М.Г., Воскресенская О.Л. Изменение активности компонентов системы антиоксидантной защиты и полифенолоксидазы у газонных растений на разных этапах онтогенеза в условиях городской среды//Физиология растений. 2008. Т. 55, № 5. С. 777-785.
- Прадедова Е.В., Ишеева О.Д., Саляев Р.К. Классификация системы антиоксидантной защиты как основа рациональной организации эксперимен-тального исследования окислительного стресса у растений//Физиология растений. 2011. Т. 58, № 2. С. 177-185.
- Практикум по физиологии растений. М.: Колос, 1972. 168 с.
- Прасад М.Н. Практическое использование растений для восстановления экосистем, загрязненных металлами//Физиология растений. 2003. Т. 50, № 5. С. 764-780.
- Рогожин В.В. Пероксидаза как компонент антиок-сидантной системы живых организмов. СПб.: ГЕОРД, 2004. 240 с.
- Роль систем антиоксидантной защиты при адаптации дикорастущих видов растений к солевому стрессу/А.В. Карташов, Н.Л. Радюкина, Ю.В. Иванов, П.П. Пашковский, Н.И. Шевякова, В.В. Кузнецов//Физиология растений. 2008. Т. 55, № 4. С. 516-522.
- Серегин И.В., Иванов В.Б. Гистохимические методы изучения распределения кадмия и свинца в растениях//Физиология растений. 1997. Т. 44, № 6. С. 915-921.
- Серегин И.В., Иванов В.Б. Физиологические аспекты токсического действия кадмия и свинца на высшие растения//Физиология растений. 2001. Т. 48, № 4. С. 606-630.
- Сигнальная роль активных форм кислорода при стрессе у растений/В.Д. Креславский, Д.А. Лось, С.Н. Аллахвердиев, Вл.В. Кузнецов//Физиология растений. 2012. Т. 59, № 2. С. 163-178.
- Титов А.Ф., Казнина Н.М., Таланова В.В. Устойчивость растений к кадмию. Петрозаводск: Карельский научный центр РАН, 2012. 55 с.
- Титов А.Ф., Таланова В.В., Казнина Н.М. Физиологические основы устойчивости растений к тяжелым металлам. Петрозаводск: Карельский научный центр РАН, 2011. 77 с.
- Чиркова Т.В. Физиологические основы устойчивости растений СПб.: Изд-во С.-Петерб. ун-та, 2002. 244 с.
- Fridovich I. Biological effects of the superoxide radical//Arch. Biochem. Biophys. 1986. Vol. 247, № 1. P. 1-11.
- Mittler R. Oxidative stress. Antioxidants and stress Tolerance//Trends plant Sci. 2002. Vol. 7. P. 405-410.
- Santa di Toppi, Gabrielli R. Response to Cadmium in Higher Plants/Environ//Exp. Bot. 1999. Vol. 41. P. 105-130.
