Активность лизосомальных гидролаз у молоди кумжи (Salmo trutta L.) из приполярной реки Ольховка
 из приполярной реки Ольховка")
Автор: Немова Нина Николаевна, Крупнова Марина Юрьевна, Вдовиченко Елизавета Андреевна, Ефремов Денис Александрович, Веселов Алексей Елпидифорович
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 4 (157), 2016 года.
Бесплатный доступ
Исследовали динамику активности лизосомальных ферментов (катепсинов В и D, ДНКазы и РНКазы) у молоди кумжи Salmo trutta L. возраста 0+, 1+, 2+, 3+ и 4+, отловленной в летний сезон (июнь 2015 года) в р. Ольховка, расположенной на Терском берегу Кольского полуострова (бассейн Белого моря). Показано, что динамика удельной активности лизосомальных гидролаз у молоди кумжи в речной период жизни коррелирует с интенсивностью роста молоди на разных этапах ее развития. Сравнительно высокие значения исследуемых ферментов обнаружены у сеголеток (личинок возраста 0+). У мальков кумжи возрастных групп 1+, 2+, 3+ и 4+ более высокие значения активности лизосомальных гидролаз (за исключением ДНКазы) обнаружены в печени в сравнении со скелетными мышцами. Эти результаты отражают участие лизосомальных ферментов нуклеинового и тесно связанного с ним белкового метаболизма в процессах роста и раннего развития лососевых рыб, особенно на этапе их выхода из нерестовых гнезд (0+).
Молодь кумжи, раннее развитие, лизосомальные протеиназы, лизосомальные нуклеазы
Короткий адрес: https://sciup.org/14751068
IDR: 14751068 | УДК: 577.152.34.032:597.553.2_113.32
Activity of lysosomal hydrolases in juvenile trout (Salmo trutta L.) from the subpolar river Ol’hovka
The dynamics of the activity of lysosomal enzymes (cathepsins B and D, DNAse and RNAse) in young salmon trout Salmo trutta L. (0+, 1+, 2+, 3+ and 4+) collected during the summer (June, 2015) from the Ol’khovka River, located on the Tersky Coast of the Kola Peninsula (the White Sea basin) was studied. It is shown that the dynamics of the activity of lysosomal hydrolases in juvenile trout during the river period of life points to a correlation between the specific activity (per mg protein) and the growth and development stage of juveniles. Relatively high values of the studied enzymes were found in the young’s-of-the-year (0+).The activity of lysosomal hydrolases was studied in the liver and muscles of trout juveniles (1+, 2+, 3+ and 4+) and the highest enzyme activity values (except DNAse) were detected in the liver. These results indicate that the lysosomal enzymes are involved in compensatory transformations of the nucleic metabolism and in closely related protein metabolism during the growth and development processes of these salmonids, especially when leaving their spawning nests (0+).
Текст научной статьи Активность лизосомальных гидролаз у молоди кумжи (Salmo trutta L.) из приполярной реки Ольховка
Кумжа (Salmo trutta L., сем. Salmonidae) – пластичный вид со сложной популяционно-генетической структурой, представленный проходной, озерной и ручьевой формами, обитает в водоемах северного полушария (бассейн Атлантического океана) и является одним из ценнейших промысловых видов лососевых рыб [14]. Условия жизни в речной период чрезвычайно важны для пополнения популяции кумжи. Период раннего онтогенеза рыб характеризуется существенными морфологическими и функциональными преобразованиями, сопровождающи- мися кардинальными перестройками клеточного метаболизма [8]. Процессы роста и развития обеспечиваются деятельностью целого комплекса механизмов, среди которых особую роль играют биохимические механизмы обмена белковых и нуклеиновых компонентов клетки [22]. Лизосомы, содержащие комплекс гидролитических ферментов, способных расщеплять все органические компоненты клетки, выполняют важную реконструктивную и защитную функцию, обеспечивая метаболические превращения в организме, в том числе в периоды раннего развития и интенсивного роста [1], [3], [10]. Поэтому представлялось
интересным провести исследование динамики изменения активности лизосомальных ферментов в процессе роста молоди кумжи начиная с этапа выхода личинок из нерестовых гнезд и до возраста 4+.
В настоящей статье приводятся данные о динамике активности лизосомальных нуклеаз (РНКазы и ДНКазы) и протеиназ (катепсинов В и D) у кумжи разных возрастных групп (0+, 1+, 2+, 3+ и 4+), отловленной в одном биотопе – приполярной реке Ольховка (бассейн Белого моря).
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
В работе были использованы химические реагенты и субстраты протеиназ и нуклеаз, произведенные «Sigma-Aldrich» (США); приборы ЦКП НО ИБ КарНЦ РАН: гомогенизатор Tissue Lyser LT (Qiagen, Германия), центрифуга Allegra 64R (Beckman Coulter, США), термостатирующая водяная баня UT-4334 (Россия), спектрофотометр СФ-2000 (ЗАО «ОКБ-Спектр», Россия).
Материал
Отлов молоди кумжи разных возрастов проводили на пороговых и перекатных участках реки Ольховка в середине июня (температура воды 11,5 °С) с помощью аппарата электролова Fa-2 («Фирма», Норвегия). Состав и объем кормовой базы для молоди кумжи, ведущей в речных условиях оседлый образ жизни на индивидуальных микростациях на порогах и перекатах, идентичен и содержит различные виды воздушных и наземных насекомых, а также водных организмов – личинок ручейников, нимф поденок и веснянок [14]. Основной прирост молоди происходит летом, при температуре воды 10–17 °С. После вылова молодь выдерживали около суток в садках, затем помещали в жидкий азот и транспортировали в лабораторию для дальнейших биохимических исследований.
Количественное содержание растворимого белка в тканях (мг/г ткани) определяли по методу М. Бредфорд [19], используя в качестве стандарта бычий сывороточный альбумин.
Определение активности лизосомальных ферментов
Активность лизосомальных протеиназ. Цистеиновая протеиназа, катепсин В (КФ 3.4.22.1), и аспартатная, катепсин D (КФ 3.4.23.5), – основные протеиназы лизосом [18]. После гомогенизации образцов в соотношении 1 : 10 (вес/об.) в растворе 0,25 М сахарозы с добавлением 0,01 % Тритона Х-100 (Merck) (1200 об./мин, 60 с) и их центрифугирования (10 000 g, 30 мин), в супернатанте определяли активность катепсина В по расщеплению 0,065 M раствора этилового эфира гидрохлорида N-бензоил L-аргинина в 0,1 M ацетатном буфере (pH 5,0) [24] и катепсина D по гидролизу 1 %-го бычьего гемоглобина в 0,1 М ацетатном буфере (рН 3,6) согласно модифицированному методу Ансона [16]. Активность катепсинов B и D (ед. акт.) выражали в единицах изменения оптического поглощения при Е525 и Е280 соответственно, на 1 мг белка за 1 ч инкубации (37 °C).
Активность лизосомальных нуклеаз. Определение активности РНКазы (КФ 3.1.4.23) и ДНКазы (КФ 3.1.4.6) проводили по методам А. П. Левицкого с соавт. [4] и А. А. Покровского с соавт. [9]. В качестве субстратов использовали 0,1 % растворы РНК и ДНК в 0,2 М ацетатном буфере (pH 5,2 и 5,0 соответственно). Количество низкомолекулярных продуктов гидролиза нуклеиновых кислот определяли спектрофотометрически при 260 нм после осаждения нерасщепленных нуклеиновых кислот и их крупных фрагментов 0,5 М раствором хлорной кислоты или 0,25%-ным уранилацетатом в 0,5 М хлорной кислоте (для ДНК и РНК соответственно). Активность ферментов выражали в условных единицах (ед. акт.) изменения оптического поглощения при 260 нм в расчете на 1 мг белка за 1 ч инкубации (30 °C).
Полученные данные обработаны статистически с помощью Microsoft Office Excel 2007. Результаты представлены в виде средних и их ошибок (М ± m). Достоверность различий оценивалась по непараметрическому критерию U Вилкоксо-на – Манна – Уитни при уровне значимости р ≤ 0,05 [2].
Экспериментальные работы выполнены с использованием оборудования ЦКП Института биологии КарНЦ РАН.
РЕЗУЛЬТАТЫ
В табл. 1 приведены данные, свидетельствующие о том, что в течение первого года жизни
Таблица 1
Размерно-весовые характеристики молоди кумжи Salmo trutta L . , n = 5
|
Показатели |
Возраст |
||||
|
0+ |
1+ |
2+ |
3+ |
4+ |
|
|
Длина тела, см |
2,6 ± 0,10 |
5,4 ± 0,3 |
9,1 ± 0,4 |
13,3 ± 0,6 |
16,5 ± 0,2 |
|
Масса тела, г |
0,11 ± 0,1 |
1,5 ± 0,1 |
7,28 ± 0,6 |
22,56 ± 0,4 |
46,4 ± 0,5 |
|
n |
6 |
5 |
5 |
5 |
5 |
Таблица 2
Активность лизосомальных гидролаз (катепсинов и нуклеаз) у молоди кумжи разных возрастов; n = 5
|
Орган |
Возраст |
Активность ферментов, ед. акт. |
|||
|
Катепсин B |
Катепсин D |
Кислая РНКаза |
Кислая ДНКаза |
||
|
Личинка целиком |
0+ |
1,80 ± 0,10 |
2,00 ± 0,15 |
0,46 ± 0,06 |
0,16 ± 0,05 |
|
Печень |
1+ |
0,60 ± 0,0 |
0,30 ± 0,0 |
0,31 ± 0,00 |
0,07 ± 0,00 |
|
2+ |
0,50 ± 0,10 |
0,14 ± 0,03 |
0,20 ± 0,02 |
0,07 ± 0,02 |
|
|
3+ |
0,50 ± 0,02 |
0,25 ± 0,04 |
0,18 ± 0,01 |
0,07 ± 0,02 |
|
|
4+ |
0,55 ± 0,02 |
0,20 ± 0,06 |
0,14 ± 0,01 |
0,05 ± 0,00 |
|
|
Мышцы |
1+ |
0,47 ± 0,06 |
0,07 ± 0,01 |
0,17 ± 0,02 |
0,07 ± 0,02 |
|
2+ |
0,37 ± 0,06 |
0,04 ± 0,0 |
0,14 ± 0,03 |
0,07 ± 0,03 |
|
|
3+ |
0,33 ± 0,08 |
0,02 ± 0,0 |
0,11 ± 0,03 |
0,06 ± 0,01 |
|
|
4+ |
0,50 ± 0,07 |
0,003 ± 0,0 |
0,11 ± 0,01 |
0,05 ± 0,00 |
|
(от 0+ до 1+) масса тела молоди кумжи возрастает более чем в 10 раз. Затем масса исследуемой молоди увеличивается меньшими темпами: в возрасте 2+, 3+ и 4+ она выше по сравнению с сеголетками в 5, 3 и 2 раза соответственно. Длина молоди кумжи от стадии 0+ до 4+ увеличивается линейно примерно в 8 раз – от 2,6 до 16,5 см.
Результаты изучения динамики активности исследуемых лизосомальных гидролаз (катепсинов и нуклеаз) у молоди кумжи различных возрастов представлены в табл. 2.
Максимальная активность исследуемых ферментов обнаружена у молоди кумжи возраста 0+. В данной группе активность ферментов определяли в целом организме личинки, поскольку ее вес составлял всего 0,11 г (табл. 1), что не давало возможности препарировать ее органы. На последующих стадиях роста и развития молоди активность ферментов определяли в печени и скелетных мышцах, которые, как известно, составляют до 70 % веса тела личинки кумжи. Активность катепсинов В и D в печени и мышцах молоди кумжи по мере роста и развития от стадии 1+ до стадии 4+ изменялась незначительно с тенденцией к снижению и составляла от таковой на стадии 0+ (сразу после выхода из нерестовых гнезд) примерно 25 % для катепсина В, 2 и 11 % в печени и мышцах соответственно для катепсина D. Такие же возрастные тенденции были обнаружены в изменении активности лизосомальных нуклеаз: активность РНКазы и ДНКазы снижалась приблизительно на 43 и 28 % в печени и на 43 и 38 % в мышцах соответственно по отношению к таковой у личинок возраста 0+. При этом значения ДНКазной активности у молоди кумжи всех исследуемых возрастов были примерно втрое ниже РНКазной. В печени молоди всех возрастных групп была выявлена сравнительно более высокая активность изученных гидролаз, чем в мышцах. Эти изменения активности лизосомальных ферментов сопровождались увеличением содержания растворимого белка по мере роста и развития рыб (рисунок).
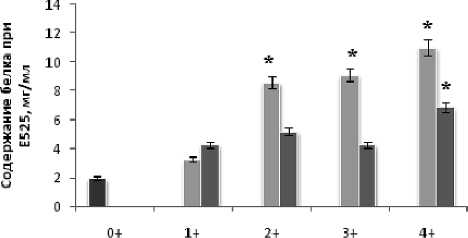
личинка целиком "печень "мышцы
Содержание растворимого белка у молоди кумжи разных возрастов (0+, 1+, 2+, 3+, 4+), n = 5
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Показанное увеличение массы тела кумжи по мере развития от сеголеток до пестряток и смолтов (2+…5+) отражает интенсивность роста молоди, напрямую связанную с накоплением костной и мышечной тканей, причем последняя играет особенно важную роль в ростовых процессах у рыб [20]. В данном исследовании показано, что наиболее значительный прирост массы тела наблюдается на первом году жизни молоди. Анализ данных показал, что темп роста молоди кумжи в данном регионе значительно выше, чем молоди атлантического лосося. Например, сеголетки рыб, выловленные в одни и те же сроки (сентябрь 2004 года) из близко расположенных водоемов, значительно различались по длине и массе. Длина АВ сеголетков кумжи из ручья Ольховка на 25 %, а вес на 133 % превышали аналогичные показатели сеголетков лосося из р. Ин- деры. Сходная картина наблюдалась и для более старших возрастных групп лососевых рыб. Так, пестрятки кумжи (1+) из ручья Собачий были больше по длине и массе (на 49 и 190 % соответственно) пестряток лосося из близко расположенного ручья Пятка. Ранее было показано [15], что в возрасте 2+ у молоди кумжи показатели длины и массы также выше, чем у лосося (на 58 и 338 % соответственно).
Процесс роста – преобразования пищевых веществ и энергии – имеет у рыб свои особенности, связанные с его недетерминированностью, в основе которой лежит преобладание синтетических процессов над катаболическими на протяжении всего жизненного цикла [5], [21], [23], [25]. Интенсивность белковой деградации служит регуляторным фактором недетерминированного роста рыб, свойственного большинству представителей класса Костистых рыб (Teleostei), включая лососевых. Синтезированные белки используются не только для построения структур и тканей молоди рыб, но и выполняют каталитическую и регуляторную функции в организме. Уровень активности основных лизосомальных протеиназ рыб – катепсинов В и D – у сеголеток (0+) кумжи имеет сравнительно высокие значения, свидетельствующие о высокой интенсивности лизосомального протеолиза, связанной, скорее всего, с задачами обеспечения «строительными блоками» (аминокислотами и пептидами) биосинтетических процессов накопления белка, необходимого для последующих структурных преобразований растущей молоди в период, когда завершается эндогенное питание и начинается переход на смешанное питание (при повышении температуры воды выше 11 °С, июнь, 500 градусо-дней). Далее, по мере роста и развития молоди кумжи, активность катепсинов в исследуемых органах изменяется незначительно (вариабельность 15–20 %), оставаясь на более низком уровне по сравнению с таковой у личинок после выклева и выхода из нерестовых гнезд (возраста 0+). В печени молоди кумжи всех изученных возрастов активность лизосомальных протеиназ, особенно катепсина D (в 5 раз), выше, чем в мышцах, что связано, вероятнее всего, с более высокой метаболической и функциональной (прежде всего синтетической) активностью печени.
Кислые нуклеазы (РНКаза и ДНКаза), локализованные в лизосомах, участвуют в расщеплении межнуклеотидных фосфодиэфирных связей в молекулах нуклеиновых кислот, которые, как известно, отвечают за передачу и хранение наследственной информации, а также ее реализацию в процессе трансляции, то есть биосинтеза белка в организме [11], [12], [17], [26]. Кислые нуклеазы осуществляют деградацию выполнивших свои функции биополимеров, тем самым обеспечивая организм компонентами для синтеза новых нуклеиновых кислот, принимающих участие в биосинтезе необходимых на новом этапе белковых веществ. Максимальная активность нуклеаз, так же как и протеиназ, обнаружена у сеголеток (0+) кумжи после их выхода из нерестовых гнезд. Ранее для молоди лосося было показано, что наиболее активные метаболические изменения связаны именно с ростом личинок в первый год жизни [5], [6], [7], [8], [13]. Можно полагать, что изменения уровня активности лизосомальных протеиназ (катепсинов В и D) и нуклеаз (РНКазы и ДНКазы) у молоди кумжи связаны с различиями в соотношениях процессов синтеза и деградации на разных стадиях развития личинок. В ранний период развития у рыб, как уже указывалось выше, преобладают синтетические процессы. Продукты, образующиеся при действии лизосомальных гидролаз у сеголеток кумжи, используются для компенсации пластических и энергетических затрат растущего организма, необходимых для метаболической перестройки при переходе от одной стадии развития мальков к другой. Успешная трансформация парров кумжи ведет к формированию смолтов, способных к миграции в море для последующего нагула.
ЗАКЛЮЧЕНИЕ
Выявленная динамика активности лизосомальных гидролаз у молоди кумжи в речной период жизни свидетельствует о наличии корреляции между активностью исследуемых ферментов и возрастом рыб, что указывает на участие лизосомальных ферментов в перестройках нуклеинового и тесно связанного с ним белкового обмена, сопровождающих процессы роста и раннего развития лососевых рыб, особенно на этапе выхода личинок из нерестовых гнезд.
* Работа выполнена при финансовой поддержке Российского научного фонда, проект №14-24-00102 «Лососевые рыбы Северо-Запада России: эколого-биохимические механизмы раннего развития».
Список литературы Активность лизосомальных гидролаз у молоди кумжи (Salmo trutta L.) из приполярной реки Ольховка
- Высоцкая Р. У., Немова Н. Н. Лизосомы и лизосомальные ферменты рыб. М.: Наука, 2008. 284 с.
- Гублер Е. В., Генкин А. А. Применение критериев непараметрической статистики для оценки различий двух групп наблюдений в медико-биологических исследованиях. М.: Медицина, 1969. 29 с.
- Дин Р. Процессы распада в клетке. М.: Мир, 1980. 120 с.
- Левицкий А. П., Барабаш Р Д., Коновец В. М. Сезонные особенности активности рибонуклеазы и α-амилазы слюны и слюнных желез у крыс линии Вистар//Биохимическая эволюция. Л.: Наука, 1973. С. 192-195.
- Лысенко Л. А., Канцерова Н. П., Крупнова М. Ю., Веселов А. Е., Немова Н. Н. Внутриклеточная белковая деградация в процессе роста Атлантического лосося Salmo salar L.//Биоорганическая химия. 2015. Т. 41. № 6. С. 717-724.
- Немова Н. Н., Крупнова М. Ю., Ефремов Д. А., Веселов А. Е. Активность лизосомальных протеиназ (катепсинов B и D) в мышцах молоди (0+, 1+, 2+) атлантического лосося из р. Варзуга//Труды КарНЦ РАН. Сер. Экспериментальная биология. 2015. № 11. С. 85-92.
- Немова Н. Н., Нефедова З. А., Мурзина С. А., Веселов А. Е., Рипатти П. О., Павлов Д. С. Влияние экологических условий обитания на динамику жирных кислот у молоди Атлантического лосося (Salmo salar L.)//Экология. 2015. № 3. С. 1-6.
- Озернюк Н. Д. Онтогенетические температурные адаптации ферментов пойкилотермных животных//Успехи современной биологии. 2004. № 6. С. 534-541.
- Покровский А. А., Арчаков А. И. Методы разделения и ферментной идентификации субклеточных фракций//Современные методы в биохимии. М.: Медицина, 1968. С. 5-59.
- Покровский А. А., Тутельян В. А. Изменение ферментов лизосом при белковой недостаточности//Биохимия. 1968. Т. 33. № 4. С. 809-816.
- Покровский А. А., Тутельян В. А. Лизосомы. М.: Наука, 1976. 351 с.
- Спирин А. С. Биосинтез белков, мир РНК и происхождение жизни//Вестник Российской академии наук. 2001. Т. 71. № 4. С. 320-328.
- Чурова М. В., Мещерякова О. В., Веселов А. Е., Немова Н. Н. Активность ферментов энергетического и углеводного обмена и уровень некоторых молекулярно-генетических показателей у молоди лосося (Salmo salar L.), различающейся возрастом и массой//Онтогенез. 2015. Т. 46. № 5. С. 304-312.
- Шустов Ю. А., Веселов А. Е. Питание и рост молоди озерной кумжи (Salmo trutta L. morphalacustris) в водоемах национального парка «Паанаярви»//Труды Карельского научного центра РАН. Сер. Экспериментальная биология. 2007. № 11. С. 142-146.
- Шустов Ю. А., Барышев И. А., Белякова Е. Н. Особенности питания молоди атлантического лосося Salmo salar L. в субарктической реке Варзуга и ее малых притоках (Кольский полуостров)//Биология внутренних вод. 2012. № 3. С. 66-70.
- Barrett A. J., Heath M. Lysosomal enzymes//Dingle J. T. (ed.). Lysosomes. A laboratory handbook. Amsterdam, 1977. P. 19-27.
- Bashan A., Yonath A. Correlating ribosome function with high-resolution structures//Trends in Microbiology. 2008. Vol. 16. № 7. P. 326-335.
- Bohley P. Intracellular proteolysis//Hydrolytic enzymes. Biomedical division. 1987. P. 307-332.
- Bradford M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding//Analit. Biochem. 1976. Vol. 72. P. 248-254.
- Bureau D., Hua K., Cho C. Y. Effect of feeding level on growth and nutrient deposition in rainbow trout (Oncorhynchus mykiss Walbaum) growing from 150 to 600 g.//Aquac. Res. 2006. Vol. 37. P. 1090-1098.
- Hagen O., Vieira V. L. A., Solberg S., Johnston I. A. Myotube production in fast myotomal muscle is switched-off at shorter body lengths in male than female Atlantic halibut (Hippoglossus hippoglossus L.) resulting in a lower final fiber number//J. Fish Biol. 2008. Vol. 73. P. 139-152.
- Hochachka P. W., Somero G. N. Biochemical adaptation/mechanisms and process in physiological evolution. 2nd edition. New York: Oxford University Press, 2002. 466 p.
- Lepage S. E., Bruce A. E. Characterization and comparative expression of zebrafish calpain system genes during early development//Dev. Dyn. 2008. Vol. 237. P. 819-829.
- Matsuda K., Misaka E. Studies on cathepsins of rat liver lysosomes. I. Purification and multiple forms//J. Biochem. 1974. Vol. 76. № 3. P. 639-649.
- Overturf K., Gaylor T. G. Determination of relative protein degradation activity at different life stages in rainbow trout (Oncorhynchus mykiss)//Comp. Biochem. Physiol. 2009. Vol. 152B. P. 150-160.
- Pizzo E., Varcamonti M., Maro A. D., Zanfardino A., Giancola C., D’Alessio G. Ribonucleases with angiogenic and bactericidal activities from the Atlantic salmon//FEBS Journal. 2008. Vol. 275. P. 1283-1295.
