Активность основных ферментов синтеза полиаминов в природных изолятах Escherichia coli

Автор: Ахова Анна Викторовна, Шегина Елена Сергеевна, Лаврикова Анастасия Леонидовна, Кузнецова Марина Валентиновна, Ткаченко Александр Георгиевич
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 3, 2019 года.
Бесплатный доступ
Исследована способность природных изолятов Escherichia coli, выделенных из разных мест обитания, декарбоксилировать лизин, орнитин и аргинин. Данный признак является широко распространенным среди E. coli, поскольку лишь у 7% изолятов не было выявлено способности декарбоксилировать ни одну из трех аминокислот. Способность декарбоксилировать аргинин встречалась реже, чем способность декарбоксилировать лизин или орнитин. Природные изоляты характеризовались разными комбинациями аргинин-, орнитин- и лизиндекарбоксилазной активностей. Наиболее часто встречалось сочетание лизиндекарбоксилазной и орнитиндекарбоксилазной активности, в то время как сочетание орнитиндекарбоксилазной и аргининдекарбоксилазной активности в отсутствие способности декарбоксилировать лизин не выявлено. Зависимости наличия способности декарбоксилировать ту или иную аминокислоту изолятом от источника выделения не установлено.
Кадаверин, путресцин, спермидин, декарбоксилаза, лизин, аргинин
Короткий адрес: https://sciup.org/147227091
IDR: 147227091 | УДК: 579.22 | DOI: 10.17072/1994-9952-2019-3-291-299
Activity of polyamine biosynthesis enzymes in Escherichia coli isolates
The ability of Escherichia coli isolated from different habitats to decarboxylate lysine, ornithine, and arginine was studied. The decarboxylase activity is widespread among Escherichia coli, since only 7% of the isolates showed no ability to decarboxylate any of the three amino acids. The ability to decarboxylate arginine was less common than the ability to decarboxylate lysine or ornithine. Natural isolates were characterized by different combinations of arginine, ornithine, and lysine decarboxylase activity. The most common combination was lysine decarboxylase and ornithine decarboxylase activity, while the combination of ornithine decarboxylase and arginine decarboxylase activity, in the absence of decarboxylation of lysine, was not detected. There were no differences in the decarboxylase activity patterns of isolates from different sources.
Текст научной статьи Активность основных ферментов синтеза полиаминов в природных изолятах Escherichia coli
Полиамины – природные поликатионы, необходимые для нормального развития эукариот, прокариот и вирусов, относятся к классу алифатических углеводородов с двумя и более амино- или иминогруппами в составе молекулы [Tabor, Tabor, 1985; Kusano et al., 2008; Pegg, 2016].
Качественный и количественный состав полиаминов варьирует у различных видов микроорганизмов, а также зависит от возраста культуры и условий культивирования [Tabor, Tabor, 1985]. Основными полиаминами Escherichia coli являются путресцин (1,4-диаминобутан), кадаверин (1,5-диаминопентан) и спермидин (N-(3-аминопропил)-1,4-диаминобутан) [Igarashi, Kashiwagi, 2010]. Кроме того, в клетках
E. coli при специфических условиях роста могут образовываться аминопропилкадаверин (N-(3-аминопропил)-1,5-диаминопентан), ацетилированные формы спермидина и путресцина, а также глутатио-нилспермидин [Tabor, Tabor, 1985; Chattopadhyay, Chen, Tabor, 2013]. Основные пути синтеза полиаминов в клетках E. coli представлены на рис. 1.
Первым этапом синтеза полиаминов является декарбоксилирование аминокислот-предшественников. Путресцин в клетках E. coli может синтезироваться двумя путями: 1) прямое декарбоксилирование орнитина при участии орнитиндекарбоксилазы (ОДК); 2) декарбоксилирование аргинина аргининдекарбоксилазой (АДК) с образованием
агматина, который затем гидролизуется агматину-реогидролазой (агматиназой) до мочевины и путресцина [Tabor, Tabor, 1985]. Кадаверин синтезируется в результате реакции декарбоксилирования лизина, катализируемой лизиндекарбоксилазой (ЛДК) [Sabo et al., 1974]. Спермидин синтезируется из L-метионина, который вначале аденилируется, а затем декарбоксилируется. В завершении полиамин-аминопропилтрансфераза синтеза
(спермидинсинтаза) переносит аминопропильную
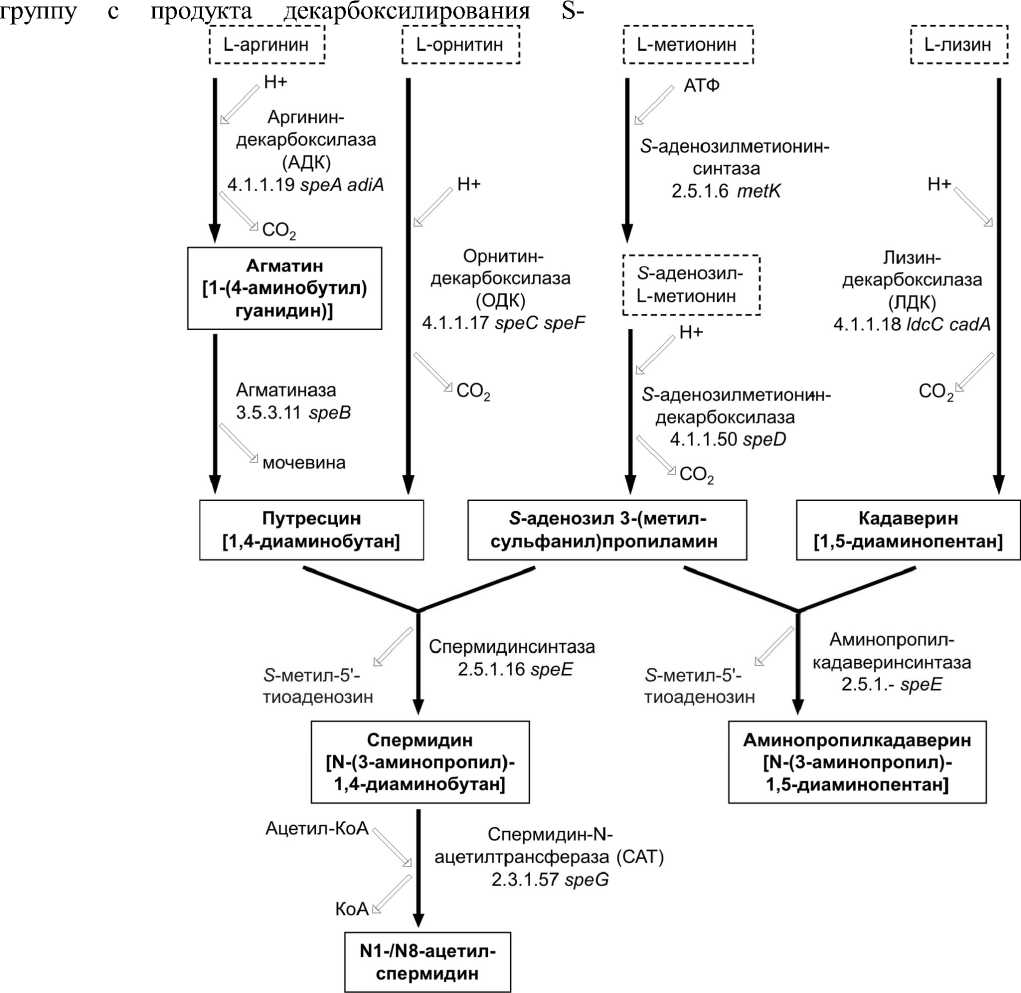
Рис. 1 . Основные метаболические пути синтеза полиаминов E. coli K12
Ферменты, катализирующие реакции декарбоксилирования аминокислот-предшественников, могут быть представлены в клетках в двух формах: биосинтетические (конститутивные) и биодеграда-тивные (индуцибельные) [Morris, Fillingame, 1974]. Изоформы различаются биохимическими свойствами, условиями активации и физиологическими функциями.
Индуцибельная форма АДК кодируется геном adiA и представляет собой гомодекамер, максимально активный при низких значениях рН (оптимум рН = 5.2) [Blethen, Boeker, Snell, 1968]. Повышение активности АДК AdiA наблюдается в клетках, выращенных на богатой среде (в присут- аденозилметионина на путресцин с образованием спермидина [Markham et al., 1980]. Полиамин-аминопропилтрансфераза также может катализировать присоединение аминопропильной группы к кадаверину с образованием аминопропилкадаве-рина и к спермидину с образованием спермина [Bowman, Tabor, Tabor, 1973]. Последняя реакция протекает с относительно низкой скоростью, и в обычных условиях накопление спермина за счет синтеза в клетках E. coli не происходит.
ствии избытка субстрата) при низких значениях рН и в условиях низкой аэрации. Физиологической ролью данного фермента является противодействие закислению цитоплазмы и поддержание гомеостаза рН внутри бактериальной клетки. По всей вероятности, аргинин-зависимая система кислото-устойчивости функционирует за счет удаления протонов водорода из клетки в составе продукта реакции декарбоксилирования аргинина – путресцина [Richard, Foster, 2004]. Ген speA кодирует конститутивную форму АДК, которая активна в форме гомотетрамера в слабощелочной и нейтральной среде (оптимум рН = 8.4). Для формирования тетрамера и проявления активности необхо- димы ионы Mg2+. Ингибитором АДК SpeA, как на уровне транскрипции, так и на уровне активности белка, выступает продукт катализируемой ею реакции – путресцин [Wu, Morris, 1973]. Биосинтетическая форма фермента экспрессируется вне зависимости от изменения рН и вовлечена в процесс биосинтеза полиаминов. АДК SpeA локализуется в районе клеточной оболочки и предпочтительно утилизирует аргинин, поступающий из среды [Buch, Boyle, 1985].
В клетках E. coli обнаружено две изоформы лизиндекарбоксилазы – LdcC и CadA (LdcI). Инду-цибельная форма ЛДК кодируется геном cadA и активна в форме гомодекамера. Индукция данной изоформы происходит при культивировании на богатой среде (в присутствии избытка субстрата) в условиях слабой аэрации и низких значений рН [Auger, Bennett, 1989; Meng, Bennett, 1992]. Кроме того, на Vibrio vulnificus показано, что данная лизиндекарбоксилаза входит в soxRS -регулон и индуцируется в ответ на супероксидный окислительный стресс [Pomposiello, Bennik, Demple, 2001; Kim, Choi, Lee, 2006]. Оптимум рН ЛДК CadA равняется 5.5, и данный фермент является частью лизин-зависимой системы кислотоустойчивости, которая работает по принципу, описанному выше для системы защиты, основанной на декарбоксилировании аргинина [Kanjee, Houry, 2013]. В регуляцию экспрессии cadA вовлечены различные транскрипционные регуляторы, а также алармон ppGpp, отрицательно регулирующий активность ЛДК CadA как на уровне экспрессии гена, так и на уровне активности белка [Kanjee et al., 2011]. Описанная несколько позднее лизиндекарбоксилаза LdcC, считающаяся конститутивным ферментом, проявляет активность в широком диапазоне рН, максимум которой приходится на значения, близкие к нейтральным (оптимум рН = 7.6) [Kikuchi, Kurahashi, Nagano, 1997; Lemonnier, Lane, 1998]. ЛДК LdcC, как и индуцибельная форма фермента, активна в виде гомодекамера. Известно, что промотор гена ldcC специфичен для σS-субъединицы РНК-полимеразы, и экспрессия данного гена повышается в стационарной фазе роста и под действием фторхинолоновых антибиотиков [Kikuchi, Kurahashi, Nagano, 1997; Шумков и др., 2010]. Ингибиторами активности данного белка являются путресцин и спермидин, а также (p)ppGpp [Wertheimer, Leifer, 1983; Kanjee et al., 2011].
Биосинтетическая и биодеградативная формы орнитиндекарбоксилазы кодируются генами speС и speF , соответственно, и функционируют в форме димеров. Обе изоформы ОДК наиболее активны в нейтральных и слабощелочных условиях (для SpeC оптимум рН = 8.1; для SpeF оптимум рН = 7) [Applebaum, Dunlap, Morris, 1977; Kanjee et al., 2011].
Экспрессия гена speC находится под контролем
RpoS, а также отрицательно регулируется ц-АМФ, Crp и ppGpp [Wright, Boyle, 1982]. Активность данной изоформы ОДК стимулируется ГТФ и ингибируется путресцином, спермидином и ppGpp [Holtta, Janne, Pispa, 1972; Applebaum, Dunlap, Morris, 1977]. Кроме того, в регуляцию активности ОДК вовлечена двухкомпонентная сигнал-проводящая система atoC-atoS [Kyriakidis, Tiligada, 2009]. Активность фермента SpeF положительно регулируется ди-, три-, тетра- и пентафосфатами гуанозина [Kanjee et al., 2011]. Считается, что ОДК SpeC вовлечена в процесс биосинтеза полиаминов, а физиологической функцией ОДК SpeF является деградация орнитина. Кроме того, предполагается, что данная орнитиндекарбоксилаза может быть частью системы кислотоустойчиво-сти, функционирующей при слабом кислотном стрессе [Kanjee, Houry, 2013].
В данной работе проведен анализ действия природных изолятов E. coli на способность декарбоксилировать лизин, орнитин и аргинин.
Материалы и методы исследования
В качестве объектов исследования использованы 62 природных изолята E. coli и штамм E. coli К12, полученный из Всероссийской коллекции микроорганизмов.
Природные изоляты E. coli были выделены из фекалий здоровых животных фермерских хозяйств Пермского края (5 культур выделены от коров, 5 – от кроликов и 4 – от свиней). Птичьи патогенные E. coli (n=33) изолированы из внутренних органов вынужденно убитой птицы (цыплят-бройлеров) в период 2016–2018 гг. (крупное птицеводческое хозяйство Пермского края). Клинические уропато-генные E. coli (n = 5) получены из материала (моча, катетеры) пациентов с инфекцией мочевыводящих путей (ИМВП), находившихся на стационарном (9 медицинских организаций) лечении в г. Перми в 2017 г. Симбионтные эшерихии (n = 10) изолированы от людей при диспансеризации в 2018 г. Все изоляты были идентифицированы согласно Приказу МЗ СССР №535 от 22.04.1985 г.
Бактериологическое исследование проводили с использованием среды МакКонки («Merck», США), идентифицируя бактерии с помощью диагностической системы «Еnterotest16» («Lachema», Чехия).
Определение декарбоксилазной активности проводили методом Мёллера с использованием декарбоксилазного бульона (ДК-бульона) («Sigma», Индия) – полноценной питательной среды, не содержащей изначально лизин, аргинин и орнитин [Moeller, 1955].
Для получения инокулята микроорганизмы, сохраняемые в полужидком Luria-Bertani-агаре («Amresco», США) засевали в 5 мл Luria-Bertani-бульона («Amresco», США) и культивировали без перемешивания при 37ºС в течение 18–20 ч. Полученные бактериальные культуры разводили в ДК-бульоне, содержащем 10 г/л лизина, аргинина или орнитина («Panreac», Испания), в соотношении 1:10. Культивирование проводили в лунках 96-луночного планшета (общий объем культуры – 200 мкл) без перемешивания при 37ºС. Изменение окраски регистрировали визуально каждый час в течение 6 ч. В качестве контроля бактерии параллельно выращивали на ДК-бульоне без добавления аминокислот.
Результаты
Наличие основных ферментов синтеза полиаминов было проанализировано у представителей E. coli , выделенных из разных мест обитания. В частности, исследованы обитатели кишечника здоровых сельскохозяйственных животных, культуры, изолированные из разных органов птицы при ко-либактериозе, а также симбионтные и уропатоген-ные эшерихии человека.
Определение декарбоксилазной активности проводили методом Мёллера с использованием в качестве селективной среды декарбоксилазного бульона (ДК-бульона) с добавкой лизина, аргинина или орнитина. Данный метод был разработан для идентификации энтеробактерий на основе их способности декарбоксилировать перечисленные выше аминокислоты. За счет наличия в составе ДК-бульона индикаторов рН бромкрезолового фиолетового и крезолового красного его цвет в процессе культивирования может изменяться от желтого (рН<7) до фиолетового (рН>7). В состав ДК-бульона входит декстроза, ферментация которой бактериями приводит к накоплению кислых продуктов обмена и снижению рН среды, что сопровождается появлением желтой окраски. Закисление среды стимулирует декарбоксилазную активность бактерий, что приводит к накоплению полиаминов и защелачиванию среды культивирования, которая в результате окрашивается в фиолетовой цвет [Moeller, 1955]. Таким образом, поскольку на первых этапах культивирования происходит закисление среды, данным методом определяется, в первую очередь, активность индуцибельных форм ферментов.
Способность декарбоксилировать орнитин, аргинин или лизин является широко распространенным признаком среди Escherichia coli , поскольку лишь у 7% изолятов не было выявлено способности декарбоксилировать ни одну из трех аминокислот. Из 62 исследованных изолятов лизиндекарбоксилазной активностью обладали 53, орнитиндекарбоксилазную активность проявляли 43 изолята, а способность декарбоксилировать аргинин была выявлена у 24 изолятов (таблица). Лабораторный штамм К12 проявлял лизиндекарбоксилазную активность.
Проявление декарбоксилазной активности (изменение окраски среды культивирования) природными изолятами E. coli
|
№ изо-лята / штамм |
ДК-бульон |
ДК-бульон + лизин |
ДК-бульон + орнитин |
ДК-бульон + аргинин |
|
Время, ч. |
||||
|
1 2 3 4 5 6 |
1 2 3 4 5 6 |
1 2 3 4 5 6 |
1 2 3 4 5 6 |
|
|
1 |
------ |
------ |
------ |
------ |
|
2 |
------ |
------ |
------ |
- - - + + + |
|
3 |
------ |
+ + + + + + |
------ |
------ |
|
4 |
------ |
------ |
------ |
------ |
|
5 |
------ |
------ |
------ |
------ |
|
6 |
------ |
------ |
------ |
- - - + + + |
|
7 |
------ |
+ + + + + + |
------ |
------ |
|
8 |
------ |
------ |
------ |
-----+ |
|
9 |
------ |
+ + + + + + |
- - + + + + |
------ |
|
10 |
------ |
- + + + + + |
- + + + + + |
------ |
|
11 |
------ |
- + + + + + |
------ |
------ |
|
12 |
------ |
------ |
------ |
------ |
|
13 |
------ |
------ |
------ |
-----+ |
Окончание таблицы
|
№ изо-лята / |
ДК-бульон |
ДК-бульон + лизин |
ДК-бульон + орнитин |
ДК-бульон + аргинин |
||||||||||||||||||||
|
штамм |
Время, ч. |
|||||||||||||||||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
1 |
2 |
3 |
4 |
5 |
6 |
1 |
2 |
3 |
4 |
5 |
6 |
1 |
2 |
3 |
4 |
5 |
6 |
|
|
14 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
|
15 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
|
16 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
+ |
|
17 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
|||||
|
18 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
|||||
|
19 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
|||||
|
20 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
------ |
------ |
|||||||||||||||
|
21 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
------ |
------ |
|||||||||||||||
|
22 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
------ |
------ |
||||||||||
|
23 |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
+ |
+ |
+ |
- |
- |
- |
+ |
+ |
+ |
|
24 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
|
25 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- - - - - + |
------ |
||||||||||
|
26 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
- |
- |
- |
- |
- |
- |
|
27 |
------ |
- |
- |
- |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
+ |
+ |
+ |
|||||
|
28 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
------ |
------ |
|||||||||||||||
|
29 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
|
30 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
|
31 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
|
32 |
------ |
------ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
||||||||||
|
33 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
- |
+ |
+ |
+ |
|
34 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
|
35 |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
+ |
+ |
+ |
|
36 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
------ |
- |
- |
- |
+ |
+ |
+ |
|||||
|
37 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
|||||
|
38 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
|||||
|
39 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
|||||
|
40 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
|||||
|
41 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
|||||
|
42 |
------ |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
+ |
- |
- |
- |
- |
+ |
+ |
|||||
|
43 |
------ |
- |
- |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
|||||
|
44 |
------ |
- |
- |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
|||||
|
45 |
------ |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
|||||
|
46 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
|||||
|
47 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
|||||
|
48 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
|
49 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
|
50 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
|
51 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
|||||
|
52 |
------ |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
|||||
|
53 |
------ |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
|||||
|
54 |
------ |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
|||||
|
55 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
------ |
- |
- |
+ |
+ |
+ |
+ |
||||||||||
|
56 |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
|
57 |
------ |
- |
- |
+ |
+ |
+ |
+ |
------ |
------ |
|||||||||||||||
|
58 |
- |
- |
- |
- |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
|
59 |
- |
- |
- |
- |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
|
60 |
- |
- |
- |
- |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
|
61 |
------ |
- |
- |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
- |
- |
|||||
|
62 |
- |
- |
- |
- |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
- |
- |
- |
+ |
+ |
|
K12 |
------ |
- |
+ |
+ |
+ |
+ |
+ |
------ |
------ |
|||||||||||||||
Способность декарбоксилировать лизин у большинства исследованных изолятов определялась уже на 1–2 ч. культивирования. Орнитиндекарбоксилазная активность регистрировалась после двух часов культивирования, а способность декарбоксилировать аргинин – после 3 ч. с момента засева инокулята. Разница во времени активации для разных декарбоксилаз может быть связана с состоянием инокулята, в качестве которого использовались клетки в стационарной фазе роста, переход к которой является одним из условий индукции лизиндекарбоксилазы [Kikuchi, Kurahashi, Nagano, 1997]. С другой стороны, при культивировании бактерий на ДК-бульоне закисление среды происходит постепенно и индуцибельная аргининдекарбоксилаза, обладающая наиболее низким значением оптимума рН, должна активироваться позже других ферментов.
Природные изоляты характеризовались разными комбинациями аргинин-, орнитин- и лизиндекарбоксилазной активностей (рис. 2). Из всех исследованных изолятов 17 были способны декарбоксилировать все три аминокислоты. Два изолята проявляли лизиндекарбоксилазную и аргининдекарбоксилазную активности. Наиболее часто у природных изолятов встречалось сочетание лизиндекарбоксилазной и орнитиндекарбоксилазной активностей (24 изолята). Изолятов, обладающих орнитиндекарбоксилазной и аргининдекарбоксилазной активностью в отсутствие способности декарбоксилировать лизин, не выявлено.
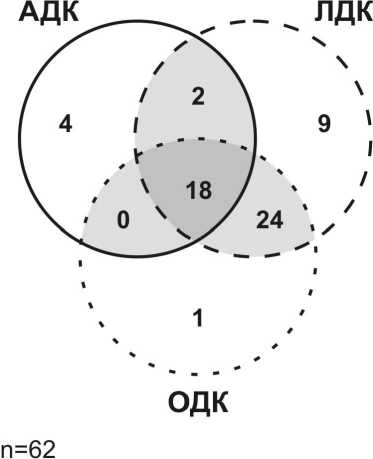
Рис. 2. Встречаемость аргинин-, орнитин- и лизиндекарбоксилазной активности в природных изолятах E. coli
Следует отметить, что использованный метод определения декарбоксилазной активности, по-видимому, применим только для высокоактивных ферментов. Об этом свидетельствует отсутствие изменения окраски среды при культивировании лабораторного штамма К12 на среде, содержащей орнитин и аргинин, несмотря на то, что в геноме данного микроорганизма присутствуют гены, кодирующие орнитин- и аргининдекарбоксилазы. Вероятно, в условиях наших экспериментов активность ферментов в клетках лабораторного штамма и накопление полиаминов в среде культивирования недостаточно велики для того, чтобы вызвать изменение ее окраски.
Таким образом, активность ключевых ферментов синтеза полиаминов обнаруживается у большинства представителей E. coli , изолированных из разных мест обитания. Способность декарбоксилировать аргинин встречается реже, чем способность декарбоксилировать лизин или орнитин. Зависимости наличия способности декарбоксилировать ту или иную аминокислоту изолятом от источника выделения не установлено. В результате работы были выявлены обладающие способностью декарбоксилировать лизин, аргинин и/или орнитин микроорганизмы, которые потенциально могут быть использованы в качестве продуцентов 1,5-диаминопентана и 1,4-диаминобутана.
Работа выполнена в рамках государственного задания, номер госрегистрации темы 01201353249.
Список литературы Активность основных ферментов синтеза полиаминов в природных изолятах Escherichia coli
- Шумков М.С. и др. Изменение экспрессии ldcC Escherichia coli как фактор адаптации к антибиотикам // Вестник Пермского университета. Сер. Биология. 2010. Вып. 1. С. 36-40.
- Applebaum D., Dunlap J., Morris D. Comparison of the biosynthetic and biodegradative ornithine decarboxylases of Escherichia coli // Biochemistry. 1977. Vol. 16, № 8. P. 1580-1584.
- Auger E., Bennett G. Regulation of lysine decarboxylase activity in Escherichia coli K-12 // Archives of Microbiology. 1989. Vol. 151, № 5. P. 466-468.
- Blethen S., Boeker E., Snell E. Arginine decarboxylase from Escherichia coli. I. Purification and specificity for substrates and coenzyme // Journal of Biological Chemistry. 1968. Vol. 243, № 8. P. 1671-1677.
- Bowman W., Tabor C., Tabor H. Spermidine biosynthesis. Purification and properties of propylamine transferase from Escherichia coli // Journal of Biological Chemistry.1973. Vol. 248, № 7. P. 2480-2486.
- Buch J., Boyle S. Biosynthetic arginine decarboxylase in Escherichia coli is synthesized as a precursor and located in the cell envelope // Journal of Bacteriology. 1985. Vol. 163, № 2. P. 522-527.
- Castanie-Cornet M. et al. Control of acid resistance in Escherichia coli // Journal of Bacteriology. 1999. Vol. 181. P. 3525-3535.
- Chattopadhyay M., Chen W., Tabor H. Escherichia coli glutathionylspermidine synthetase/amidase: phylogeny and effect on regulation of gene expression // FEMS Microbiology Letters. 2013. Vol. 338, № 2. P. 132-140.
- Höltta E., Jänne J., Pispa J. Ornitine decarboxylase from Escherichia coli: stimulation of the enzyme activity by nucleotides // Biochemical and Biophysical Research Communications. 1972. Vol. 47. P. 1165-1171.
- Igarashi K., Kashiwagi K. Modulation of cellular function by polyamines // International Journal of Biochemistry and Cell Biology. 2010. Vol. 42, № 1. P. 39-51.
- Kanjee U. et al. The enzymatic activities of the Escherichia coli basic aliphatic amino acid decarboxylases exhibit a pH zone of inhibition // Biochemistry. 2011. Vol. 50, № 43. P. 9388-9398.
- Kanjee U., Houry W.A. Mechanisms of acid resistance in Escherichia coli // Annual Review of Microbiology. 2013. Vol. 67. P. 65-81.
- Kikuchi Y., Kurahashi O., Nagano T. RpoSdependent expression of the second lysine decarboxylase gene in Escherichia coli // Bioscience, Biotechnology, and Biochemistry. 1997. Vol. 62, № 6. P. 1267-1270.
- Kim J., Choi S., Lee J. Lysine decarboxylase expression by Vibrio vulnificus is induced by SoxR in response to superoxide stress // Journal of Bacteriology. 2006. Vol. 188, № 24. P. 8586-8592.
- Kusano T. et al. Polyamines: essential factors for growth and survival // Planta. 2008. Vol. 228, № 3. P. 367-381.
- Kyriakidis D., Tiligada E. Signal transduction and adaptive regulation through bacterial two-component systems: the Escherichia coli AtoSC paradigm // Amino Acids. 2009. Vol. 37, № 3. P. 443-458.
- Lemonnier M., Lane D. Expression of the second lysine decarboxylase gene of Escherichia coli // Microbiology. 1998. Vol. 144, № 3. P. 751-760.
- Markham G. et al. S-adenosylmethionine synthetase from Eschericgia coli // Journal of Biological Chemistry. 1980. Vol. 255. P. 9082-9092.
- Meng S., Bennett G. Nucleotide sequence of the Escherichia coli cad operon: a system for neutralization of low extracellular pH // Journal of Bacteriology. 1992. Vol. 174, № 8. P. 2659-2669.
- Moeller V. Simplified tests for some amino acid decarboxylases and for the arginine dihydrolase system // Acta Pathologica et Microbiologica Scandinavica. 1955. Vol. 36. P. 158-172.
- Morris D., Fillingame R. Regulation of amino acid decarboxylation // Annual Review of Biochemistry. 1974. Vol. 43. P. 303-321.
- Pegg A. Functions of polyamines in mammals // Journal of Biological Chemistry. 2016. Vol. 291, № 29. P. 14904-14912.
- Pomposiello P., Bennik M., Demple B. Genome-wide transcriptional profiling of the Escherichia coli responses to superoxide stress and sodium salicylate // Journal of Bacteriology. 2001. Vol. 183, № 13. P. 3890-3902.
- Richard H., Foster J. Escherichia coli glutamate- and arginine-dependent acid resistance systems increase internal pH and reverse transmembrane potential // Journal of Bacteriology. 2004. Vol. 186, № 18. P. 6032-6041.
- Sabo D. et al. Purification and physical properties of inducible Escherichia coli lysine decarboxylase // Biochemistry. 1974. Vol. 13. P. 662-670.
- Tabor C.W., Tabor H. Polyamines in microorganisms // Microbiological Reviews. 1985. Vol. 49. P. 81-99.
- Wu W., Morris D. Biosynthetic arginine decarboxylase from Escherichia coli. Purification and properties // Journal of Biological Chemistry. 1973. Vol. 248, № 5. P. 1687-1695.
- Wendisch V. Microbial production of amino acidrelated compounds // Advances in Biochemical Engineering/Biotechnology. 2017. Vol. 159. P. 255-269.
- Wertheimer S., Leifer Z. Putrescine and spermidine sensitivity of lysine decarboxylase in Escherichia coli: evidence for a constitutive enzyme and its mode of regulation // Biochemical and Biophysical Research Communications. 1983. Vol. 114, № 2. P. 882-888.
- Wright J., Boyle S. Negative control of ornithine decarboxylase and arginine decarboxylase by adenosine-3,5-monophosphate in E. coli // Molecular and General Genetics. 1982. Vol. 186. P. 482-487.
