Активность протолитических ферментов растений под воздействием лектинов эндофитных и эпифитных штаммов Azospirillum

Автор: Аленкина С.А., Никитина В.Е.
Журнал: Журнал стресс-физиологии и биохимии @jspb
Статья в выпуске: 2 т.12, 2016 года.
Бесплатный доступ
Мы изучили способность лектинов, изолированных от поверхности двух штаммов азотфиксирующих почвенных бактерий рода Azospirillum, A. brasilense Sp7 (эпифит) и A. brasilense Sp245 (эндофит), чтобы показать, оказывают регулирующее действие на активность пектинолитических ферментов в корнях саженцев пшеницы. Результаты исследований показали, что изучаемые лектины могут вызывать индукцию активности полигалактуроназы, пектинэстеразы, пектатлязы из клеточной стенки растения, тем самым обеспечивая проникновение бактерий в растительные ткани, а также индукцию ответов растений, которые в сочетании с стимулирующий рост бактерий, способствует формированию стабильности и продуктивности растений.
Короткий адрес: https://sciup.org/14323991
IDR: 14323991
Plant cell protolytic enzymes activity under exposure to lectins of endophytic and epiphytic Azospirillum strains
We studied the ability of lectins isolated from the surface of the two strains of nitrogen-fixing soil bacteria of the genus Azospirillum, A. brasilense Sp7 (epiphytic) and A. brasilense Sp245 (endophytic), to show have a regulating effect on the activity of pectinolytic enzymes in the roots of wheat seedlings. Research results showed that the lectins under study can cause the induction of the activity of polygalacturonase, pectinesterase, pectatlyase from the plant cell wall, thereby ensuring the bacteria penetration in the plant tissues, as well as the induction of plants responses which, being combined with growth-stimulating effect of bacteria, contributes to the formation of plants stability and productivity.
Текст научной статьи Активность протолитических ферментов растений под воздействием лектинов эндофитных и эпифитных штаммов Azospirillum
Бактерии рода Azospirillum (альфа-субкласс протеобактерий) известны много лет как улучшающие рост растений ризобактерии (plant growth promoting rhizobacteria - PGPR) (Okon, Vanderleyden, 1997).
Бактерии обладают рядом очень важных свойств для эффективного ассоциативного взаимодействия с растениями - способностью к азотфиксации, продукции фитогормонов, солюбилизации фосфатов, улучшению водного и минерального статуса, продукции ряда соединений, увеличивающих мембранную активность и пролиферацию тканей корневой системы, уменьшать влияние стрессоров на растение и осуществлять контроль многочисленных фитопатогенов (Bashan et al. , 2004). Они были изолированы из ризосферы многих трав и злаков, произрастающих как в тропическом так и в умеренном климате. Азоспириллы, в основном колонизируют поверхность корня, и лишь некоторые штаммы способны проникать во внутренние ткани корня (Steenhoudt, Vanderleyden, 2000; Saikia et al. , 2012). Вопросы, связанные с проникновением бактерий в ткани корня растений являются одними из невыясненных в исследовании азотфиксирующих бактериально-растительных ассоциаций.
Фитопатогенные и мутуалистические микроорганизмы, такие как Rhizobium и Azospirillum являются известными продуцентами ферментов деградирующих пектин клеточной стенки растений (Khammas et al., 1989). В тоже время показана способность ризобий за счет Nod фактора индуцировать активность этих ферментов в самой растительной клетке на начальных этапах взаимодействия (Xie et al., 2012).
С действием пектолитических ферментов связывают не только процессы разрушения пектиновых веществ при проникновении и продвижении по тканям растения, но и изменение проницаемости для воды и ионов. Повреждающее действие пектолитических ферментов вызывает сдвиг физиологических процессов и развитие в тканях растения ответных реакций, соотношение которых может определять исход взаимоотношений между организмами.
Как показано в большинстве случаев, пектинолитические ферменты растений, являются гликопротеинами и, следовательно, несут детерминанты специфичности (Albersheim, Valent, 1978). Присутствие в молекулах пектолитических ферментов углеводов (Cervone et al. , 1981) может обусловливать специфическое взаимодействие с лектинами, в том числе бактериальными.
Никитина с соавт. (1996) показала присутствие на поверхности клеток азоспирилл лектинов, вовлеченных в бактериальную адгезию к корням. С поверхности двух отличающихся по способу колонизации растений штаммов ассоциативных азотфиксирующих бактерий -A. brasilense Sp7 и A. brasilense Sp245 были изолированы лектины, являющиеся гликопротеинами с различными молекулярными массами и углеводной специфичностью (Никитина с соавт., 2005; Шелудько с соавт., 2009). Было показано, что лектины азоспирилл являются полифункциональными молекулами. Помимо адгезивной функции, они способны влиять на метаболизм растительной клетки - стимулировать прорастание семян (Никитина с соавт., 2004), проявлять по отношению к растительной клетке митогенную и ферментмодифицирующую активности (Alen’kina et al., 2006; Чернышева с соавт., 2005), изменять содержание стрессовых метаболитов в растительной клетке, что свидетельствует о способности лектинов выступать в качестве индукторов адаптационных процессов корней проростков пшеницы (Alen’kina et al., 2014).
В связи с вышесказанным, целью данной работы была сравнительная оценка способности лектинов A. brasilense Sp7 и A. brasilense Sp245 оказывать регулирующее воздействие на активность пектинолитических ферментов в корнях проростков пшеницы.
MATERIALS AND METHODS
В работе были использованы лектины двух штаммов бактерий рода Azospirillum – A. brasilense Sp7 (эпифитный) полученный из Института микробиологии им. С.Н. Виноградского РАН (г. Москва) и A. brasilense Sp245 (эндофитный) из коллекции микроорганизмов ИБФРМ РАН. Выращивание культур, выделение и очистку лектинов проводили, как описано ранее (Никитина с соавт. , 1996; Шелудько с соавт. , 2009).
В работе использовали 3-х суточные проростки пшеницы Саратовская 29. Корни после экспозиции с препаратами лектинов (концентрации лектинов 5-40 мкг/мл, максимальное время инкубации – 1 ч), гомогенизировали в 0.15 М фосфатном буфере (рН 7.8). Гомогенат центрифугировали при 7000g, надосадочную жидкость использовали для определения активности ферментов.
Активность полигалактуроназы, пектатлиазы и пектинэстеразы определяли по описанной методике (Ермаков с соавт. , 1987). Активность ферментов выражали числом единиц активности фермента в 1 г сырой массы корней.
Полученные результаты подвергали статистической обработке. Доверительные интервалы определяли для 95% уровня значимости.
RESULTS AND DISCUSSION
Результаты проведенных исследований показали, что изучаемые лектины в концентрациях от 5 до 20 мкг/мл после 1ч воздействия на корни проростков пшеницы способны вызывать индукцию активности полигалактуроназы, пектинэстеразы, пектатлиазы -ферментов пектинолитического комплекса клеточной стенки растений. Самой эффективной оказалась концентрация обоих лектинов 10 мкг/мл. Наибольший эффект наблюдался по отношению к полигалактуроназе, увеличение активности для лектинов A. brasilense Sp7 и Sp245 составляло 80 и 150%, соответственно. Несколько меньшим оказалось влияние лектинов на активность пектатлиазы. В данном случае увеличение активности составило 50% для лектина A. brasilense Sp7 и 100% для лектина A. brasilense Sp245. Самым незначительным было влияние лектинов на активность пектинэстеразы. Лектин эпифитного штамма вызывал увеличение активности фермента на 10%, эндофитного штамма на 50%. Результаты продемонстрировали наиболее высокую индуцирующую активность лектина эндофитного штамма Sp245 по отношению ко всем изучаемым ферментам, что по-видимому связано со способностью данного штамма проникать внутрь корня.
Таким образом, полученные данные свидетельствуют о том, что лектины азоспирилл способны вызывать индукцию пектинразрушающих ферментов самих растений, являющихся одним из основных факторов, способствующих разрушению клеточных стенок и внедрению микроорганизмов в ткани растения и в этом прослеживается сходство с бобово-ризобиальным симбиозом (Xie et al., 2012).
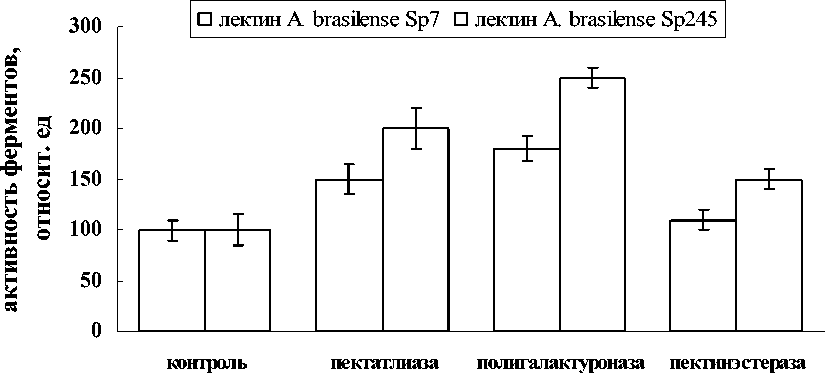
Figure 1. Влияние лектинов A. brasilense Sp7 и Sp245 на активность пектолитических ферментов корней проростков пшеницы. Контроль-корни: активность пектатлиазы – 20 ед/г сырой массы, полигалактуроназы – 10 ед/г сырой массы, пектинэстеразы – 25 ед/г сырой массы. Концентрация лектинов – 10 мкг/мл. Время инкубации – 1ч.
Роль пектолитических ферментов не ограничена только процессами разрушения пектиновых веществ, происходящих при проникновении и продвижении микроорганизмов по тканям растения. Повреждающее действие пектолитических ферментов вызывает в клетке сдвиг физиологических процессов и развитие ответных реакций в тканях растения. Соотношение этих реакций, скорость и интенсивность могут определять исход взаимоотношений между организмами. Клеточная стенка растений рассматривается как "депо" ферментов: в ней обнаружен целый ряд ферментов, в том числе и пероксидазы. Показано, что пероксидазы локализованы в свободном пространстве клеточной стенки, а также связаны ионными или ковалентными связями (Mader et al.,1980; Sasaki et al., 2008). Многие исследователи наблюдали активирование и освобождение пероксидаз, локализованных в клеточных стенках, при действии пектолитических ферментов (Гладких и др., 1979). С действием пероксидаз связывают образование этилена (Lund, Mapson, 1970) и перекиси водорода (Bhattachrjee, 2005), оказывающих токсическое действие на ткани растения, окисление индолилуксусной кислоты (ИУК), изменяющее гормональный баланс в клетках (Cheng, Song, 2006). Полагают, что растворенные и активированные пероксидазы, связанные с клеточными стенками, вызывают в клетках хозяина развитие метаболических изменений, нарушающих скоординированные физиологические процессы в клетках.
Вероятно, вышесказанное объясняет тот факт, что лектин A. brasilense Sp7, штамма, не обладающего способностью проникать в ткани растения, также как и лектин A. brasilense Sp245 (эндофитного штамма) способен вызывать, хотя и в меньшей степени, индукцию пектолитических ферментов растительной клетки.
Полученные результаты и предыдущие данные, свидетельствующие о способности лектинов азоспирилл вызывать спектр ответных биохимических реакций, являющихся частью сигнальных систем (Alen’kina et al. , 2014), дают основания полагать, что лектины азоспирилл могут индуцировать развитие ответных реакций в растениях, что в сочетании с ростстимулирующим эффектом бактерий способствует формированию устойчивости и продуктивности растений.
Список литературы Активность протолитических ферментов растений под воздействием лектинов эндофитных и эпифитных штаммов Azospirillum
- Ермаков А.И., Арасимович В.В., Ярош Н.П., Перуанский Ю.В., Луковникова Г.А., Иконникова М.И. (1987) Методы биохимического исследования растений Л.: Агропромиздат
- Гладких Т.А., Давыдова М.А., Молдосанова Г.А., Васильева К.В., Метлицкий Л.В. (1979). Действие препаратов пектин-трансэлиминазы Verticillium dahliae Klebahn на ткани и растения хлопчатника. Прикл. биохимия и микробиология, 15(2), 194-205
- Никитина В.Е., Аленькина С.А., Пономарева Е.Г., Савенкова Н.Н. (1996) Изучение роли лектинов клеточной поверхности азоспирилл во взаимодействии с корнями пшеницы. Микробиология, 65, 165-170
- Никитина В.Е., Богомолова Н.В., Пономарева Е.Г., Соколов О.И. (2004) Влияние лектинов азоспирилл на способность семян к прорастанию. Известия РАН. Серия биологическая, 4, 431-435
- Никитина В.Е., Пономарева Е.Г., Аленькина С.А. Лектины клеточной поверхности азоспирилл и их роль в ассоциативных взаимоотношениях с растениями. Молекулярные основы взаимоотношений ассоциативных микроорганизмов с растениями/Под ред. В.В. Игнатова.-М.: Наука, 2005. 70-97
- Чернышева М.П., Аленькина С.А., Никитина В.Е., Игнатов В.В. (2005) Внеклеточные протеолитические ферменты штамма Azospirillum brasilense Sp7 и регулирование их активности гомологичным лектином. Прикл. биохимия и микробиология, 41, 444-448
- Шелудько А.В., Пономарева Е.Г., Варшаломидзе О.Э., Ветчинкина Е.И., Кацы Е.И., Никитина В.Е. (2009) Гемагглютинирующая активность и подвижность бактерий Azospirillum brasilense в присутствии разных источников азота. Микробиология, 6, 749-756
- Albersheim P., Valent B. (1978) Host-pathogen interactions in plants. J Cell Biology, 78, 627-643
- Alen’kina S.A., Payusova O.A., Nikitina V.E. (2006) Effect of Azospirillum lectins on the activities of wheat-root hydrolytic enzymes. Plant and Soil, 283, 147-151
- Alen’kina S.A., Bogatyrev V.A., Matora L.Yu., Sokolova M.K., Chernysheva M.P., Trutneva K.A., Nikitina V.E. (2014) Signal effects of the lectin from the associative nitrogen-fixing bacterium Azospirillum brasilense Sp7 in bacterial-plant root interactions. Plant and Soil, 381, 337-349
- Bashan Y., Holguin G., de-Bashan L.E. (2004) Azospirillum-plant relationships: physiological, molecular, agricultural, and environmental advances (1997-2003). Can. J. Microbiol., 50, 521-577
- Bhattachrjee S (2005). Reactive oxygen species and oxidative burst: Roles in stress, senescence and signal transduction in plant. Current Science, 89, 1113-1121
- Cervone F., Anderbhan T., Corn R. H. A., Wood R. K. S. (1981) Effects of french bean tissue on Colletotrichum lindemuthianum polygalacturonase. Phytopathol. Z., 102, 238-246
- Cheng Y, Song C (2006). Hydrogen peroxide homeostasis and signaling in plant cells. Sci China C Life Sci., 49, 1-11
- Khammas K.M., Ageron., Grimond P.A., Kaiser P. (1989) Azospirillum irakense spp. nov., a nitrogen-fixing bacterium associated with rice roots and rhizosphere soil. Res. Microbiol., 140, 679-693
- Lund B.M., Mapson L.W. (1970) Stimulation by Erwinia carotovora of the synthesis of ethylene in cauliflower tissue. Biochem. J., 119, 251-263
- Mader M., Ungemach J., Schlop P. (1980) The role of peroxidase isozyme groups of Nicotiana tabacum in hydrogen peroxide formation. Planta, 147(5), 467-470
- Okon Y., Vanderleyden J. (1997) Root-associated Azospirillum species can stimulate plants. ASM News, 63, 366-370
- Sasaki S., Nonaka D., Wariishi H., Tsutsumi Y., Kondo R. (2008) Role of Tyr residues on the protein surface of cationic cell-wall-peroxidase (CWPO-C) from poplar: potential oxidation sites for oxidative polymerization of lignin. Phytochemistry, 69, 348-355
- Saikia S.P., Bora D., Goswami A., Mudoi K.D., Gogoi A. (2012) A review on the role of Azospirillum in the yield improvement of non leguminous crops. African J. Microbiol. Res., 6, 1085-1102
- Steenhoudt O., Vanderleyden J. (2000) Azospirillum, a free-living nitrogen-fixing bacterium closely associated with grasses: genetic, biochemical and ecological aspects. FEMS Microbiol. Rev., 24, 487-506
- Xie F., Murray J.D., Kim J., Heckmann A.B., Edwards A., Oldroyd G.E.D., Downiе J. A. (2012) Legume pectate lyase required for root infection by rhizobia. PNAS, 109, 633-638
