Альгофлора рек Пермского края в условиях антропогенного засоления отходами калийного производства

Автор: Мартыненко Н.А.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 2, 2017 года.
Бесплатный доступ
Изучен видовой состав альгофлоры р. Яйвы, Волима, Чёрной и Лёнвы Пермского края. Представлен таксономический список водорослей и цианопрокариот, включающий в себя 300 видов и разновидностей из 106 родов, 59 семейств, 33 порядков и 7 отделов. Основу видового разнообразия слагают диатомовые водоросли; ведущие семейства: Fragilariaceae, Bacillariaceae и Cymbellaceae. Большинство идентифицированных таксонов относятся к бентосным формам с широким ареалом. По отношению к галобности большинство принадлежат к индифферентам и галофилам, по отношению к кислотности среды - к алкалифилам и индифферентам, а по отношению к сапробности -к в-мезо и олигосапробам. Представлены дендрограммы исследованных рек и участков, основанные на индексе сходства биоценозов Чекановского-Серенсена.
Видовой состав, альгофлора, реки, повышенная минерализация
Короткий адрес: https://sciup.org/147204820
IDR: 147204820 | УДК: 574.58
Algoflora of the rivers of Permskiy krai under anthropogenic salinization of potassium production wastes
We studied the species composition of algoflora of Yaiva, Volim, Chernaya and Lеnva rivers of Perm krai in this article. A check-list of algae and cyanoprokaryota presented, it includes 300 taxa below the genus from 106 genera, 59 families, 33 orders and 7 divisions. The diatoms have composed most of the species diversity; the leading families: Fragilariaceae, Bacillariaceae and Cymbellaceae. Most of identified taxa are benthic forms with a world-wide area. Most of them is indifferent and halophilic in relation to salinity, alkaliphilic and indifferent with respect to pH and в-meso and oligosaprobic towards saprobity. Dendrograms of studied rivers, based on Sorensen-Czekanowski's index of similarity of biocenosis, presented in this paper.
Текст научной статьи Альгофлора рек Пермского края в условиях антропогенного засоления отходами калийного производства
Среди водных объектов проточного типа самыми многочисленными, разнообразными и уязвимыми являются небольшие по протяженности водотоки (средние и малые реки). Именно они, несмотря на размеры, составляют основу гидрографической сети, формируют крупные реки, определяя их гидрологический и биохимический режимы. Но эти водные экосистемы и их компоненты до сих пор остаются наименее изученными [Фролова, 2004]. Важным компонентом водных экосистем является фитопланктон. Иденти- фикация того или иного индикаторного вида может служить одним из важных показателей состояния водотока, поэтому изучение видового состава водного объекта имеет огромное значение для оценки степени загрязнения и, в частности, засоления водоёмов и водотоков.
Первые сведения о видовом составе альгофло-ры бассейна р. Камы на территории Пермского края ещё до её зарегулирования содержатся в работе А.О. Таусон [1947]. После появления среднекамских водохранилищ практически важным стало изучение их автотрофного звена. Данные о видовом составе водорослей и цианопрокариот Воткин-
ского водохранилища приведены в работе С.А. Третьяковой, С.И. Головачевой, Е.М. Батовой [1988], фитопланктона Камского водохранилища – в работе П.Г. Беляевой [2013]. Также был изучен видовой состав фитоперифитона р. Сылвы [Беляева, 2004] и состав доминантных комплексов р. Чусовой [Беляева, Поздеев, 2005]. Был уточнён видовой состав центрических диатомовых водорослей, роль и функционирование цианопрокариот Камского и Воткинского водохранилищ, а также десмидиевых водорослей некоторых водоёмов Пермского края [Тарасова, 2005; Генкал, Охапкин, 2010; Беляева, 2011; Генкал, Беляева, 2011; Мартыненко, 2015].
Водоёмы с повышенной минерализацией традиционно рассматриваются как местообитания с экстремальными условиями существования. Лимитирующее влияние высокой солености способствует формированию специфического биоценоза, структура которого значительно отличается от пресноводной биоты [Ventosa, Arahal, 1999]. Необходимым условием для понимания механизмов функционирования сообществ водоемов в условиях лимитирующего влияния солености является определение качественного и количественного состава водорослей и цианопрокариот и их связи с абиотическими факторами [Селиванова, Немцева, 2011].
Цель данной работы – изучение видового состава альгофлоры рек в условиях антропогенного засоления на примере р. Яйвы, Волима, Чёрной и Лёнвыа Камского бассейна.
Физико-географическая характеристика района исследований
Под малыми реками мы, вслед за А.М. Комлевым и Е.А. Черных [1984], подразумеваем водотоки длиной от 10 до 100 км, под средними – от 100 до 500 км.
Река Яйва – средняя река длиной 304 км, левый приток р. Камы, а в настоящее время – Камского водохранилища. Её исток расположен вблизи границы Свердловской области на хр. Кваркуш. Верхнее течение этой реки лежит в Александровском р-не, нижнее течение проходит по Усольскому р-ну. Река впадает в Камское водохранилище на 879 км от устья у пос. Яйвинский Рейд, южнее г. Березники. Благодаря подпору водохранилища на последних километрах образуется залив. В р. Яйву впадают 267 притоков общей протяжённостью 506 км, общая площадь водосбора – 6 250 км². На исследованных нами участках вода в реке, согласно классификации С.П. Китаева [2007], относится к классу особо пресных (0.05–0.09‰). Минерализация вод реки повышается до уровня среднепресной (0.13‰) после впадения р. Волима.
Река Волим – малая река (12 км), впадающая в р. Яйву на 7.8 км от устья по правому берегу у д. Володин Камень. Река Волим протекает в пределах Березниковского и Усольского р-нов Пермского края в юго-западном направлении, затем поворачивает на северо-запад. Русло извилистое, количество притоков – 31 с общей протяжённостью 38 км. Река протекает в непосредственной близости от Второго калийного комбината группы «Уралкалий», её воды на значительном протяжении сильно засолены отходами калийного производства [Формирование загрязнения…, 2015]. Подпор Камского водохранилища, в которое впадает р. Яйва, периодически распространяется и на р. Волим, на последнем километре течения реки образуется залив. В верхнем течении воды Волима относятся к классам среднепресных и пресноватых (0.09–1.01‰). После впадения р. Чёрной вода в р. Волиме становится средне- и сильносолоноватой (7.54–18.64 ‰).Р ека Чёрная – правый приток р. Волима, длиной около 4.8 км. В связи с тем, что солеотвалы Второго калийного комбината расположены на площади её водосбора, она засолена от истока до устья. Вода в ней сильносолёная (44.58–51.31‰).
Река Лёнва – левый приток Камского водохранилища протяжённостью 21 км, впадающий в него на 881 км от устья, у пос. Яйвинский Рейд. Верхнее и нижнее течение проходит по городскому округу Березники, среднее – по территории Усольского р-на. Исток реки находится на территории городского округа Березники к югу от платформы «Калийная», рядом со шламохранилищем и солеотвалами рудника БКПРУ-2 («Уралкалий»). В связи с этим р. Лёнва засолена от истока до устья, вода в ней средне- и сильносолоноватая (6.56–10.08 ‰).
Все изученные реки являются нейтральнощелочными с высоким содержанием кислорода. Данные о гидрохимии изученных рек приведены в табл. 1.
Таблица 1
Гидрохимические показатели воды: температура (t°С), окислительно-восстановительный потенциал (ОВП), реакция среды ( рН ), содержание кислорода в воде ( O 2 ), электропроводность ( S ), содержание растворённых веществ ( С ), солёность (PSU) рек на исследованных участках
|
Река |
t°С |
ОВП (мВ) |
pH |
O 2 (%Sat) |
S (мСм/см) |
С (мг/л) |
PSU |
|
Яйва |
14.3 |
44.8 |
8.0 |
96.3 |
286 |
186 |
0.10 |
|
13.3-15.7 |
29.7-57.7 |
7.8-8.2 |
93.2-101.5 |
160-408 |
104-265 |
0.05-0.13 |
|
|
Волим |
9.3 |
25.9 |
7.8 |
110.4 |
8879 |
5772 |
5.17 |
|
6.6-10.3 |
6.1-58.5 |
7.5-8.2 |
92.9-131.9 |
351-30090 |
228-19560 |
0.11-18.64 |
Окончание табл. 1
|
Река |
t°С |
ОВП ( мВ ) |
pH |
O 2 (%Sat) |
S ( мСм /см) |
С (мг/л) |
PSU |
|
Чёрная |
10.3 |
9.6 |
7.2 |
118.5 |
70983 |
46136 |
48.79 |
|
8.7-12.7 |
2.8-15.8 |
7.2-7.3 |
109.5-129.1 |
65710-74140 |
42710-48190 |
44.58-51.31 |
|
|
Лёнва |
11.9 |
34.3 |
7.5 |
104.7 |
14340 |
9318 |
8.32 |
|
11.6-12.6 |
28.8-39.8 |
7.5-7.6 |
98.3-111.1 |
11530-17150 |
7495-11140 |
6.56-10.08 |
Примечание. Над чертой – средняя величина, под чертой – min и max значения
Материал и методы
Всего была собрана и обработана 31 проба: 7 проб на 4 участках р. Яйвы, 19 проб на 8 участках р. Волима, 3 пробы на 1 участке р. Чёрной, 2 пробы на 2 участках р. Лёнвы.
Пробы были отобраны в течение двух вегетационных сезонов: в сентябре 2013 г. на станциях № 1, 4, 7 и 12; в мае 2014 г. – на станциях № 1–9 и сентябре 2014 г. – на всех вышеперечисленных станциях (рис. 1).
Пробы фитопланктона объёмом 1 л отбирали с поверхностного слоя воды. Сгущение проведено фильтрационным методом. Качественные пробы макрофитовых водорослей брали 15 или 50 мл пробиркой. Обработка проб проводилась под микроскопом «Ломо-Микмед 1» на базе Пермского отделения ФГБНУ «ГосНИОРХ». Идентификацию проводили с помощью определителей из серии «Определители пресноводных водорослей СССР» [1951–1986], а также с помощью «Краткого определителя…» [Царенко, 1990].
Систематическое положение таксонов принимали по справочнику «Водоросли-индикаторы в оценке качества окружающей среды» [Баринова, Медведева, Анисимова, 2000], с уточнениями для Cyanophyta, Chlorophyta и Streptophyta [Царенко, 2005; Taxonomic …, 2014]. Классификация высших таксонов диатомовых водорослей (Bacillario-phyta) приведена по системе Ф. Раунда с соавторами [Round, Crawford, Mann, 1990]. Для уточнения классификации были использованы сводки по отдельным группам [Round, Bukhtiyarova, 1996; Diatoms of Europe, 2001; 2003; 2009; Diatom monographs, 2003]. Сокращения авторов таксонов приводили по рекомендациям П.М. Царенко [2010].
Для сравнения альгофлор изученных рек использовали родовой коэффициент (Кр) – отношение количества внутриродовых таксонов к количеству родов [Охапкин, 1997]. Эколого-географические характеристики внутриродовых таксонов приведены по литературным данным [Водоросли, 1989; Баринова, Медведева, Анисимова, 2000; Diversity …, 2004]. Сходство альгоценозов оценивали посредством коэффициента Чекановского-Серенсена (Kч-с) [Sörensen, 1948]. С помощью полученной матрицы сходства методом невзвешенных парногрупповых средних (UPGMA) была по- строена дендрограмма в программе STATISTICA v. 10.
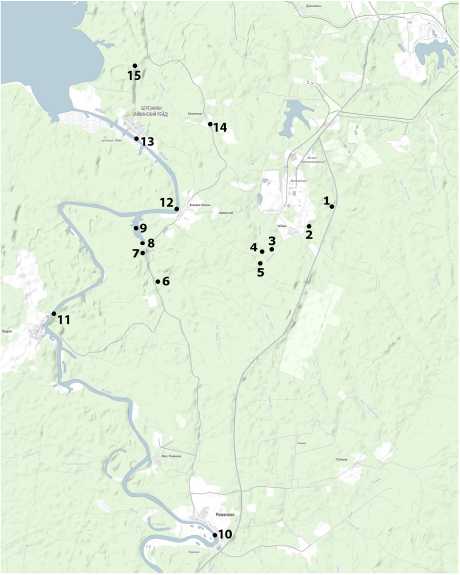
Рис. 1 . Карта-схема рек в районе исследования:
1-3 и 5-9 – точки сбора материала на р. Волиме : 1 – ниже моста на трассе «Пермь-Березники» на 950 м; 2 – ниже моста на трассе в д. Сибирь на 10 м; 3 – выше устья р. Чёрной на 500 м; 5 – ниже устья р. Чёрной на 50 м; 6 – выше моста на трассе «Романово-Володин Камень» на 50 м; 7 – ниже залива на 500 м; 8 – ниже залива на 200 м; 9 – залив в устье р. Волима; 4 – на р. Чёрной ; 10-13 – на р. Яйве : 10 – у с. Романово, 11 – у д. Белая Пашня, 12 – у д. Володин Камень, 13 – у пос. Яйвинский Рейд; 14-15 – на р. Лёнве : 14 – выше моста у д. Балахонцы на 50 м; 15 – ниже моста на трассе «Березники – Яйвинский Рейд» на 50 м
Результаты и их обсуждение
Общая характеристика альгофлоры
В результате проведённых работ в альгофлоре района исследования было идентифицировано 300 видов и разновидностей водорослей и цианопрокариот, принадлежащих к 106 родам, 59 семействам, 33 порядкам и 7 отделам.
Диатомовые водоросли составляют 73% таксономического списка альгофлоры изученных рек. Наибольшим видовым богатством отличаются семейства Fragilariaceae (25 видов и разновидностей), Bacillariaceae (25) и Cymbellaceae (23); рода Navicula (17), Nitzschia (17), Cymbella (12) и Gom-phonema (12).
Зелёные водоросли формируют 14% видового богатства альгофлоры. Более половины их видового богатства приходится на семейство Scenedes- maceae (25), а наиболее богатые видами рода – Desmodesmus (11), Scenedesmus (6), Monoraph-idium (4).
Цианопрокариоты составляют около 7% видового списка альгофлоры. Ведущим семейством по числу таксонов является Oscillatoriaceae (10), а лидирующим родом – Oscillatoria (5). Представители отделов Chrysophyta, Dinophyta, Euglenophyta и Streptophyta, в сумме составляют около 6% общего видового богатства (табл. 2).
Таблица 2
Таксономическая структура альгофлоры исследованных рек
|
Отдел |
Число |
% от общего числа |
|||||
|
порядков |
семейств |
родов |
видов |
внутривидовых таксонов |
таксонов, определённых до рода |
||
|
Bacillariophyta |
15 |
33 |
63 |
179 |
25 |
15 |
73.0 |
|
Chlorophyta |
7 |
9 |
18 |
40 |
– |
2 |
14.0 |
|
Chrysophyta |
2 |
2 |
2 |
– |
– |
2 |
0.7 |
|
Cyanoprocaryota |
4 |
8 |
12 |
16 |
– |
4 |
6.7 |
|
Dinophyta |
1 |
1 |
1 |
– |
– |
1 |
0.3 |
|
Euglenophyta |
1 |
1 |
4 |
7 |
– |
3 |
3.3 |
|
Streptophyta |
3 |
5 |
6 |
1 |
1 |
4 |
2.0 |
|
Всего |
33 |
59 |
106 |
243 |
26 |
31 |
100.0 |
Видами, которые встречаются на каждом из исследованных участков, являются диатомеи из семейства Fragilariaceae : Fragilaria capucina Desm., Diatoma vulgare Bory и Ulnaria ulna (Nitzsch) Compère. Всего около половины (49%) идентифицированных внутриродовых таксонов отмечены более чем на одном участке, и только 9% – более чем в половине изученных участков.
Далее приведён список водорослей и цианопрокариот, идентифицированных в р. Яйве, Волиме, Чёрной и Лёнве.
Отдел Dinophyta
Порядок Peridiniales
Peridinium sp.
Отдел Chrysophyta
Порядок Chromulinales
Mallomonas sp.
Порядок Ochromonadales
Ochromonas sp.
Отдел Chlorophyta
Порядок Chlamydomonadales
Chlamydomonas sp.
Порядок Chlorococcales
Botryococcus braunii Kütz.
Порядок Sphaeropleales
Acutodesmus acuminatus (Lagerh.) E. Hegew.
-
A. falcatus (Lagerh.) P. Tsarenko
-
A. acutiformis (Schröd.) P. Tsarenko et D.M. John Ankistrodesmus falcatus (Corda) Ralfs
-
A. fusiformis Corda
Coelastrum astroideum De Not.
-
C. microporum Nägeli
Desmodesmus abundans (Kirchn.) E. Hegew.
-
D. bicaudatus (Dedus.) P. Tsarenko
-
D. denticulatus (Lagerh.) An, Friedl et E. Hegew.
-
D. dispar (Bréb.) E. Hegew.
-
D. intermedius (Chodat) E. Hegew.
-
D. magnus (Meyen) P. Tsarenko
-
D. microspina (Chodat) P. Tsarenko
-
D. granulatus (West et G.S. West) P. Tsarenko
-
D. quadricaudatus (Turpin) Bréb.
-
D. esmodesmus sp.
D. spinosus (Chodat) E. Hegew.
Enallax costatus (Schmidle) Pascher
Monoraphidium arcuatum (Korschikov) Hindák
M. contortum (Thur.) Komárk.-Legn.
M. griffithii (Berk.) Komárk.-Legn.
M. tortile Komárk.-Legn.
Pediastrum boryanum (Turpin) Menegh.
P. tetras (Ehrenb.) Ralfs
Pseudodidymocystis lineata (Korsсhikov) Hindák
Scenedesmus apiculatus (West et G.S. West) Chodat
S. disciformis (Chodat) Fott et Komárek
S. ellipticus Corda
S. grahneisii (Heynig) Fott
S. obliquus (Turpin) Kütz.
S. obtusus Meyen
Tetraëdron minimum (A. Braun) Hansg.
Tetrastrum triangulare (Chodat) Komárek
Порядок Chlorellales
Dictyosphaerium pulchellum H.C. Wood
Порядок Treubouxiales
Microthamnion strictissimum Rabenh.
Порядок Ulotrichales
Enteromorpha intestinalis (L.) Link
Порядок Siphonocladales
Cladophora glomerata (L.) Kütz.
C. rivularis (L.) Hoek
Rhizoclonium hieroglyphicum (C. Agardh) Kütz.
Отдел Streptophyta
Порядок Zygnematales
Mougeotia sp.
Spirogyra sp.
Порядок Desmidiales
Closterium pronum var. brevius W. West
Cosmarium sp.
Staurastrum sp.
Порядок Klebsormidiales
Klebsormidium subtile (Kütz.) Tracanna
Отдел Euglenophyta
Порядок Euglenales
Euglena limnophila Lemmerm.
-
E. oblonga Schmitz
-
E. sp.
Lepocinclis sp.
Phacus hamatus Pochm.
Trachelomonas mirabilis Svirenko
T. oblonga Lemmerm.
T. ovalis Daday
T. planctonica Svirenko
T. sp.
Отдел Cyanoprocaryota
Порядок Chroococcales
Chroococcus turgidus (Kütz.) Nägeli
Порядок Synechococcales
Merismopedia punctata Meyen
Limnothrix planctonica (Wolosz.) Meffert
Порядок Nostocales
Anabaena sp.
Aphanizomenon flos-aquae (L.) Ralfs
Calothrix sp.
Dolichospermum flosaquae (Bréb.) P. Wacklin,
-
L. Hoffm. et Komárek
Trichormus variabilis (Kütz.) Komárek et Anagn.
Порядок Oscillatoriales
Lyngbya sp.
Oscillatoria limosa C. Agardh
-
O. major Vaucher
O. nitida Schkorb.
O. sp.
O. tenuis C. Agardh
Phormidium tambii (Woron.) Anagn. et Komárek
-
P. terebriforme (C. Agardh) Anagn. et Komárek
P. ingricum (Woron.) Anagn. et Komárek
P. willei (Gardner) Anagn. et Komárek
Planktothrix agardhii (Gomont) Anagn.
P. rubescens (DC.) Anagn. et Komárek
Отдел Bacillariophyta
Порядок Thalassiosirales
Cyclotella comta (Ehrenb.) Kütz.
-
C. kuetzingiana Thw.
-
C. meneghiniana Kütz.
-
C. sp.
Stephanodiscus astraea (Ehrenb.) Grunow
S. binderanus (Kütz.) Krieg.
-
S. hantzschii Grunow
Порядок Coscinodiscales
Actinocyclus normanii (W. Greg.) Hust.
Порядок Melosirales
Melosira moniliformis (O. Müll.) C. Agardh
M. undulata (Ehrenb.) Kütz.
M. varians C. Agardh
Порядок Aulacoseirales
Aulacoseira distans (Ehrenb.) Simonsen
-
A. granulata (Ehrenb.) Simonsen
-
A. sp.
Порядок Fragilariales
Asterionella formosa Hassal
Ctenophora pulchella var. lanceolata (O'Meara) Bukht.
-
C. pulchella (Ralfs) D.M. Williams et Round Diatoma anceps (Ehrenb.) Kirchn.
-
D. elongatum (Lyngb.) C. Agardh
-
D. hiemale (Lyngb.) Heib.
-
D. mesodon (Ehrenb.) Kütz.
-
D. vulgare Bory
Fragilaria capucina Desm.
-
F. crotonensis Kitton
-
F. inflata (Heiden) Hust.
-
F. sp.
-
F. vaucheriae (Kütz.) J.B. Petersen
Fragilariforma virescens (Ralfs) D.M. Williams et Round
-
F. virescens var. capitata (Østrup) B. Hartley Meridion circulare (Grev.) C. Agardh Staurosira construens Ehrenb.
Staurosirella leptostauron (Ehrenb.) D.M. Williams et Round
S. pinnata (Ehrenb.) D.M. Williams et Round
Tabularia fasciculata (C. Agardh) D.M. Williams et Round
Ulnaria acus (Kütz.) Aboal
U. biceps (Kütz.) Compère
U. capitata (Ehrenb.) Compère
U. ulna (Nitzsch) Compère
U. ulna var. amphirhynchus (Ehrenb.) Aboal
Порядок Tabellariales
Tabellaria fenestrata (Lyngb.) Kütz.
T. flocculosa (Roth) Kütz.
Tetracyclus rupestris (A. Braun) Grunow
Порядок Eunotiales
Eunotia bilunaris (Ehrenb.) Mills
-
E. exigua (Bréb.) Rabenh.
-
E. faba (Ehrenb.) Grunow
-
E. formica Ehrenb.
-
E. gracilis (Ehrenb.) Rabenh.
-
E. minor (Kütz.) Grunow
-
E. praerupta Ehrenb.
-
E. sp.
Порядок Mastogloiales
Aneumastus tuscula (Ehrenb.) D.G. Mann et Stickle
Порядок Cymbellales
Anomoeoneis sphaerophora (Ehrenb.) Pfitzer Brebissonia lanceolata (C. Agardh) Mahoney et Reimer Cymbella affinis Kütz.
-
C. amphioxys (Kütz.) Cleve
-
C. aspera (Ehrenb.) Cleve
-
C. balatonis Grunow
-
C. cistula (Hemprich) Grunow
-
C. cymbiformis C. Agardh
-
C. falaisensis (Grunow) Krammer et Lange-Bert.
-
C. laevis Nägeli
-
C. sp.
-
C. tumida (Bréb.) van Heurck
-
C. tumidula Grunow
-
C. turgidula Grunow
Cymbopleura naviculiformis (Auerswald) Krammer
Didymosphenia geminata (Lyngb.) Schmidt Encyonema gracile Kirchn.
-
E. hebridicum (W. Greg.) Grunow
-
E. ventricosum (C. Agardh) Grunow
Gomphoneis olivaceum (Hornem.) P. Dawson Gomphonema abbreviatum C. Agardh
G. acuminatum Ehrenb.
-
G. acuminatum var. coronatum (Ehrenb.) W. Sm.
-
G. angustatum Kütz.
-
G. intricatum Kütz.
-
G. lanceolatum Ehrenb.
-
G. longiceps Ehrenb.
-
G. longiceps var. montanum (Schum.) Cleve
-
G. parvulum Kütz.
-
G. parvulum var. lagenulum (Kütz.) Freng.
-
G. sp.
-
G. truncatum Ehrenb.
Navicymbula pusilla (Grunow) Krammer
Placoneis dicephala (W. Sm.) Mereschk.
P. elginensis (W. Greg.) E.J. Cox
P. elginensis f. exigua (W. Greg) Bukht.
P. placentula (Ehrenb.) Heinzerl.
P. placentula f. rostrata (A. Mayer) Bukht.
Rhoicosphenia abbreviata (C. Agardh) Lange-Bert.
Порядок Achnanthales
Achnanthes sp.
-
A. taeniata Grunow
Achnanthidium exiguum (Grunow) Czarn.
-
A. minutissimum var. affinis (Grunow) Bukht.
-
A. pyrenaicum (Kütz.) H. Kobayasi
-
A. exilis (Kütz.) Bukht.
Cocconeis pediculus Ehrenb.
-
C. placentula Ehrenb.
-
C. scutellum Ehrenb.
Karayevia clevei (Grunow) Round et Bukht.
Planothidium hauckianum (Grunow) Round et Bukht.
P. hauckianum var. rostratum (Schulz) Bukht.
P. lanceolatum (Bréb.) Round et Bukht.
P. rostratum (Østrup) Lange-Bert.
Rossithidium nodosum (Cleve) Aboal
Порядок Naviculales
Brachysira exilis (Kütz.) Round et D.G. Mann Caloneis silicula (Ehrenb.) Cleve
-
C. sp.
-
C. undulata (W. Greg.) Krammer
Cavinula cocconeiformis (W. Greg.) D.G. Mann et Stickle
-
C. lacustris (W. Greg.) D.G. Mann et Stickle
Cosmioneis pusilla (W. Sm.) D.G. Mann et Stickle Craticula ambigua (Ehrenb.) D.G. Mann
-
C. cuspidata (Kutz.) D.G. Mann
-
C. halophila (W. Grun.) D.G. Mann
-
C. halophila var. subcapitata (Østrup) Czarn.
Diploneis smithii (Bréb.) Cleve
-
D. sp.
Fallacia forcipata (Grev.) Stickle et D.G. Mann
Gyrosigma acuminatum (Kütz.) Rabenh.
-
G. acuminatum var. gallicum Grunow
-
G. attenuatum (Kütz.) Rabenh.
-
G. distortum (W. Sm.) Cleve
-
G. kuetzingii (Grunow) Cleve
-
G. macrum (W. Sm.) Cleve
-
G. scalproides (Rabenh.) Cleve
-
G. sp.
-
G. spenceri (W. Sm.) Cleve
Hippodonta hungarica (Grunow) Lange-Bert., Met-zeltin et A. Witkowski
-
H. capitata (Ehrenb.) Lange-Bert., Metzeltin et
-
A. Witkowski
Luticola ventricosa (Kütz.) D.G. Mann
Navicula amphibola Cleve
N. cari Ehrenb.
N. cincta (Ehrenb.) Ralfs
N. cryptocephala Kütz.
N. gregaria Donkin
N. integra (W. Sm.) Ralfs
N. kefvingensis (Ehrenb.) Kütz.
N. menisculus Schum.
N. oblonga Kütz.
N. peregrina (Ehrenb.) Kütz.
N. platystoma Ehrenb.
N. radiosa Kütz.
N. rhynchocephala Kütz.
N. salinarum Grunow
N. sp.
N. tripunctata (O. Müll.) Bory
N. viridula Kütz.
Neidium affine (Ehrenb.) Pfizer
N. affine var. amphirhynchus (Ehrenb.) Cleve
N. dubium (Ehenb.) Cleve
Parlibellus crucicula (W. Sm.) A. Witkowski, Lange-
Bert. et Metzeltin
Pinnularia distinguenda Cleve
P. divergens W. Sm.
P. gibba var. parva (Grunow) Freng.
P. interrupta W. Sm.
P. major (Kütz.) Rabenh.
P. microstauron (Ehrenb.) Cleve
P. sp.
P. subcapitata W. Greg.
Proschkinia longirostris (Hust.) D.G. Mann
Sellaphora bacillum (Ehrenb.) D.G. Mann
S. pupula (Kütz.) D.G. Mann
S. pupula var. elliptica (Hust.) Bukht.
S. pupula var. mutata (Krasske) Bukht.
Stauroneis anceps Ehrenb.
S. anceps var. gracilis (Ehrenb.) Brun
S. legumen (Ehrenb.) Kütz.
-
S. smithii Grunow
Порядок Thalassiophysales
Amphora libyca Ehrenb.
-
A. ovalis Kütz.
A. sp.
Halamphora coffeiformis (C. Agardh) Levkov
H. perpusilla (Grunow) Q.-M. You et Kociolek
H. veneta (Kütz.) Levkov
Порядок Bacillariales
Cylindrotheca closterium (Ehrenb.) Reimann et J.C. Lewin
Hantzschia amphioxys (Ehrenb.) Grunow
H. amphioxys var. major Grunow
-
H. virgata (Roper) Grunow
Nitzschia capitellata Hust.
N. clausii Hantzsch
N. communis Rabenh.
N. dubia W. Sm.
N. gracilis Hantzsch
N. hantzschiana Rabenh.
N. heufleriana Grunow
N. palea (Kütz.) W. Sm.
N. recta Hantzsch
N. sigmoidea (Nitzsch) W. Sm.
N. sp.
N. spectabilis (Ehrenb.) Ralfs
N. sublinearis Hust.
N. thermalis Kutz.
N. thermalis var . minor Hilse
N. tryblionella Hantzsch
N. vermicularis (Kütz.) Hantzsch
Tryblionella acuminata W. Sm.
T. angustata W. Sm.
T. apiculata W. Greg.
T. hungarica (Grunow) D.G. Mann
ПорядокRhopalodiales
Rhopalodia gibba (Ehrenb.) O. Müll.
R. gibberula (Ehrenb.) O. Müll.
Порядок Surirellales
Cymatopleura elliptica (Bréb.) W. Sm.
-
C. solea (Bréb.) W. Sm.
-
C. solea var. apiculata (W. Sm.) Ralfs
-
C. solea var. subconstricta O. Müll.
-
C. solea var. vulgaris F. Meister
Entomoneis alata Ehrenb.
Surirella angusta Kütz.
-
S. gracilis Grunow
S. linearis W. Sm.
S. linearis var. constricta (Ehrenb.) Grunow
S. ovalis Bréb.
S. ovata Kütz.
S. ovata var. pseudopinnata (W. Sm.) Hust.
S. ovata var. salina (W. Sm.) Hust.
S. robusta Ehrenb.
S. sp.
S. tenera W. Greg.
Общее количество всех видов и внутривидовых таксонов изученного района сравнительно велико и для альгофлоры региона – 300. Это почти равно приведённому числу видов в Камском водохранилище [Беляева, 2013], больше, чем в Воткинском водохранилище [Третьякова, Головачева, Батова, 1988]. Если рассматривать каждую реку в отдельности, то количество видов и внутривидовых таксонов водорослей и цианопрокариот велико для пресных рек такой длины [Гончаров, 1994], но меньше, чем в малых реках большей протяжённости [Фролова, 2004]. В р. Яйве число таксонов внутриродового ранга меньше, чем в р. Волиме. Число таксонов ниже родового ранга в р. Лёнве сравнимо для рек такой длины, а в р. Чёрной внут-риродовых таксонов больше, чем можно ожидать для таких рек [Гончаров, 1994]. По сравнению с числом внутриродовых таксонов высокоминерализованных рек Приэльтонья значения этого показателя в изученных нами реках также низки [Буркова, 2011; Горохова, Зинченко, 2014].
Господствующее положение отдела Bacillario-phyta по видовому богатству в альгофлоре изученных рек (72–95% общего видового богатства на участке) типично для альгофлоры водотоков умеренных широт и, по-видимому, обусловлено большой экологической пластичностью данного таксона. Меньшую представленность по данному показателю получили отделы Chlorophyta (до 19%) и Cyanoprocaryota (до 14%). Другие отделы водорослей (Chrysophyta, Dinophyta, Euglenophyta и Strep-tophyta) представлены небольшим числом таксонов внутриродового ранга на отдельных участках (табл. 3). Ведущие семейства по видовому богатству из диатомей ( Fragilariaceae , Bacillariaceae , Cymbellaceae ) и Scenedesmaceae из зелёных водорослей, вероятно, получают преимущество за счёт высокой толерантности к условиям среды. Роль эвгленовых водорослей в акватории исследованного района невелика, что нетипично для малых рек [Охапкин, 1997].
Таблица 3
Распределение видового богатства отделов водорослей и цианопрокариот в альгофлоре изученных рек (в %), число внутриродовых таксонов ( N ) и родовой коэффициент ( Kр )
|
Река |
Bacillario-phyta |
Chloro-phyta |
Chrysо-phyta |
Cyano-procaryota |
Dino-phyta |
Eugleno-phyta |
Strepto-phyta |
N |
Kр |
|
Яйва |
72 |
19 |
– |
6 |
– |
2 |
1 |
169 |
2.28 |
|
Волим |
82 |
6 |
1 |
6 |
1 |
3 |
1 |
221 |
2.95 |
|
Чёрная |
84 |
– |
– |
14 |
– |
– |
2 |
44 |
1.47 |
|
Лёнва |
95 |
– |
– |
– |
– |
5 |
– |
21 |
1.40 |
Из 300 найденных видов особый интерес пред- ставляют зелёная водоросль Enteromorpha intesti-
nalis – индикатор загрязнения хлоридами [Водоросли, 1989] и диатомея Actinocyclus normanii – каспийский вселенец, уже встречающийся в массе в Камском водохранилище [Генкал, Беляева, 2011; Беляева, 2013]. Этот вид единично отмечен в р. Яйве в зоне подпора, близ д. Володин Камень (участок «Яйва 12»).
Сравнение видового состава исследованных рек
Самое высокое значение родового коэффициента зафиксировано для альгофлоры р. Волима – 2.95, что говорит об относительно благоприятных условиях формирования разнообразия альгофлоры. А именно, на этой реке есть участки, не подверженные значительной антропогенной нагрузке; участки, сильно загрязняемые отходами калийного производства, а также – с зоогенным воздействием (бобровые запруды).
Альгофлора р. Яйвы характеризовалась меньшей величиной индекса Кр – 2.28, что, вероятно, связано с однотипностью биотопов реки в её нижнем течении, отсутствием мелководий и слабым развитием пойменной системы.
Величина индекса Кр для альгофлоры р. Чёрной была низка – 1.47, а наименьшее значение наблюдалось для таковой р. Лёнвы – 1.40. Низкие величины родового коэффициента свидетельствуют о неблагоприятных условиях формирования альгофлоры в этих реках, что, на наш взгляд, связано с сильным засолением этих водотоков.
При рассмотрении участков по отдельности максимальное значение родового коэффициента ( Kр = 2.15) наблюдалось на участке р. Волима ниже д. Сибири («Волим 3»), минимальное значение ( Kр = 1.23) – на обоих участках р. Лёнвы («Лёнва 14» и «Лёнва 15»).
Максимальный уровень видового сходства между альгофлорами разных рек, оцениваемый по индексу Чекановского-Серенсена, был равен 0.50 для Яйвы и Волима. Минимальная видовая общность ( K ч-с = 0.14) была отмечена для Лёнвы и Волима. Принимая за границу флористической общности величину 0.5 [Вайнштейн, 1976], можно заключить, что альгофло-ры изученных рек являются разобщёнными. Дендрограмма, построенная на основании коэффициента Че-кановского-Серенсена, показала сильную разобщённость альгофлор р. Лёнвы по отношению к таковой р. Яйвы и её притоков – Волима и Чёрной. Альгоф-лоры р. Яйвы и её притока первого порядка – Волима, показывают самое большое видовое сходство этих рек, находящееся на границе общности ( K ч-с = 0.50). Отличия между альгофлорами р. Яйвы и её притоком 2-го порядка – р. Чёрной – уже достаточно велики (рис. 2).
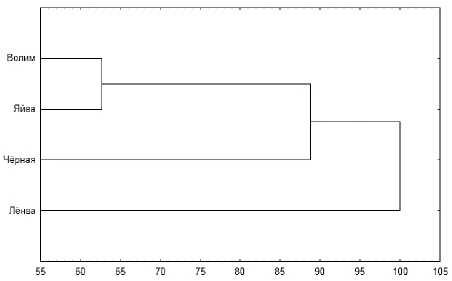
Рис. 2. Дендрограмма сходства альгоценозов изученных рек, основанная на коэффициенте Чекановского-Серенсена
Проведено сравнение видового состава водорослей и цианопрокариот на отдельных участках изученных рек. Так, максимальное сходство ( K ч-с = 0.67) зарегистрировано для участков р. Лёнвы («Лёнва 14» и «Лёнва 15»). Минимальное значение индекса видового сходства ( K ч-с = 0.12) было отмечено между нижележащим участком р. Лёнвы («Лёнва 15») и участком р. Волима ниже дер. Сибирь («Волим 3»). Тем не менее, на построенной дендрограмме отчётливо видно подразделение участков на несколько групп (рис. 3).
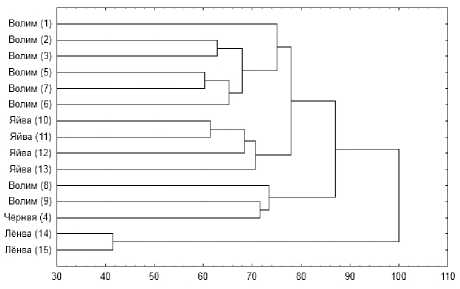
Рис. 3. Дендрограмма сходства альгоценозов отдельных участков изученных рек, основанн-ная на коэффициенте Чекановского-Серенсена
Первый кластер составляют участки р. Лёнвы («Лёнва 14» и «Лёнва 15»), альгофлора которых значительно отличается от остальных (рис. 3), что также можно наблюдать и на предыдущей денро-грамме (рис. 2). Это можно объяснить высокой солёностью реки, как следствие – минимальным среди изученных рек видовым богатством (по 16 внутриродовых таксонов), а также отдельной гидрографической сетью, не связанной с сетями других изученных рек и впадающей непосредственно в Камское водохранилище.
Второй кластер – нижние участки р. Волима («Волим 8», «Волим 9») и р. Чёрной («Чёрная 4»). Данные участки рек характеризуются экстремальными значениями солёности, электропроводности и содержания растворённых в воде веществ, а их альгоф- лора – относительно низким числом внутриродовых таксонов – 35–44 (рис. 3).
В третий кластер объединяются остальные участки р. Волима («Волим 1–3 и 5–7») (рис. 3). Кластер, в свою очередь, подразделяется на группы следующим образом. В отдельную группу выделяется участок «Волим 1» – верховья реки с самой низкой температурой и минерализацией вод, аль-гофлора этого участка отличается максимальным числом видов (109). Отдельную группу составляют участки выше и ниже дер. Сибирь («Волим 2» и «Волим 3», соответственно), где электропроводность и общая минерализация вод несколько повышаются, а видовое богатство альгофлоры, наоборот, снижается (63 и 84 таксона, соответственно). В условиях повышения солёности и электропроводности вод р. Волима ниже устья р. Чёрной («Волим 5», «Волим 6» и «Волим 7») происходит дальнейшее снижение числа таксонов (49–64) (рис. 3).
Четвёртый кластер образуют участки р. Яйвы, воды которых характеризуются минимальными значениями электропроводности, солёности и содержания растворённых веществ при максимальных среди исследованных рек температуре и окислительно-восстановительном потенциале (см. табл. 1). Альгофлора этих участков относительно богата – 73–89 видов. В составе кластера в одну группу объединяются участки р. Яйвы у с. Романово («Яйва 10») и д. Белая Пашня («Яйва 11»). Эти участки реки расположены выше устья р. Волима, кроме того, здесь отсутствует влияние подпора Камского водохранилища. Вторую группу формируют участки реки у д. Володин Камень («Яйва 12») и пос. Яйвинский Рейд («Яйва 13»), то есть часть реки, расположенная ниже устья р. Волима, и где, кроме поступления высокоминерализован- ных вод, наблюдается влияние подпора Камского водохранилища.
Эколого-географическая характеристика альгофлоры
Сравнивая видовой состав водорослей изученных рек в целом, можно заключить, что в альгоф-лоре водотоков, испытывающих засоление на всём своём протяжении, доля галофильных форм возрастает, доля индифферентных и галофобных – снижается. Так, в р. Чёрной и Лёнве вклад в аль-гофлору галофильных форм (32–36%) выше, чем в р. Волиме и Яйве (18–23%). Доля индифферентных форм во флоре р. Чёрной и Лёнвы, наоборот, ниже (48–53%), чем в таковой рек Волима и Яйвы (60–70%).
Сильное засоление рек (до 51.31‰) обусловливает высокий процент индифферентов по отношению к солёности (45–69%) и галофилов (18–40%) на участках рек в зоне влияния отходов калийного производства (табл. 4). При этом явной зависимости между относительным видовым богатством индикаторов галобности и степенью засоления на конкретных станциях наблюдения не выявлено. Например, на вышележащем участке р. Лёнвы («Лёнва 14») не обнаружено галофобных видов при уровне солёности около 10‰, а в р. Чёрной (участок «Чёрная 4») доля галофобных форм в аль-гофлоре составляет 3% при значительно б о льшей солёности (около 50‰). Также обращает на себя внимание необычное распределение индикаторов галобности (преобладание галофилов – 30–40% и высокая доля галофобов – 5–15%) в альгофлоре залива р. Волима (участки «Волим 8» и «Волим 9»), где отмечается стратификация вод по солёности.
Таблица 4
Распределение индикаторных видов водорослей и цианей в альгофлоре изученных рек (в %) по галобности (Гал.), приуроченности к месту обитания (Эко.), географическому распространению (Гео.), отношению к реакции среды (рН) и сапробности (Сап.)
|
Река |
Гал. |
Эко. |
Гео. |
рН |
Сап. |
|||||||||||||
|
ind |
hl |
mh |
hb |
B |
P |
P-B |
К |
Бор. |
А-А |
alf |
ind |
alb |
acf |
α |
β |
o |
x |
|
|
Яйва |
70 |
18 |
6 |
6 |
72 |
17 |
11 |
62 |
28 |
10 |
46 |
38 |
10 |
6 |
12 |
53 |
27 |
8 |
|
Волим |
60 |
23 |
10 |
7 |
82 |
9 |
9 |
63 |
32 |
5 |
49 |
34 |
9 |
8 |
14 |
56 |
24 |
6 |
|
Чёрная |
48 |
36 |
10 |
3 |
56 |
26 |
18 |
76 |
17 |
7 |
43 |
37 |
7 |
11 |
16 |
60 |
20 |
4 |
|
Лёнва |
53 |
32 |
10 |
5 |
65 |
17 |
18 |
88 |
12 |
– |
70 |
18 |
12 |
– |
26 |
62 |
13 |
– |
Примечание: ind – индифференты, hl – галофилы, mh – мезогалобы, hb – галофобы, B – бентосные формы, P – планктонные формы, P-B – планкто-бентосные формы, К – космополитный ареал, Бор . – бореальный ареал, А-А – арктоальпийский ареал, alf – алкалифилы, alb – алкалибионты, acf – ацидофилы, α – α -мезосапробы, β – β -мезосапробы, о – олигосапробы, х – ксеносапробы.
Вероятно, представленность индикаторов в аль-гофлоре исследованных рек определяется не только собственно концентрацией солей в воде, но и рядом других параметров, которые трудно учесть. В частности, развитием пойменной системы на конкретном участке водотока и наличием осадков, повышающих вероятность попадания галофобных видов в основное русло, а также характером течения – турбулентное или ламинарное.
В составе альгофлоры отмечены собственно планктонные формы, виды, ведущие прикреплённый образ жизни, и планкто-бентосные формы (табл. 4).
Преобладание бентосных форм (47–85%) в толще воды говорит о формировании фитопланктона рек путём смывания водными массами водорослей и цианей с донного и растительного субстратов. Фитопланктон рек, формирующийся таким образом, получил название потамофитопланктона [Грезе, 1957; Пономарёва, 2014]. Максимальная доля бентосных форм (85%) была зафиксирована на участке р. Волима ниже д. Сибири («Волим 3»).
Планкто-бентосные и планктонные формы имеют меньший вклад в альгофлоре рек – 8–32% и 2–32%, соответственно. Их максимальная роль установлена в низовьях р. Волима, где наблюдается подпор от водотока-приёмника – р. Яйвы («Волим 8» и «Волим 9»). Здесь также был отмечен наименьший вклад бентосных форм. Минимальный процент планктонных форм имеет среднее течение р. Волима («Волим 6»), а планкто-бентосных – участок р. Яйвы у пос. Яйвин-ский Рейд («Яйва 13»).
При сравнении альгофлоры изученных водотоков в р. Волиме сформировалась наибольшая доля бентосных форм (82%) и наименьшая – планктонных и планкто-бентосных форм (по 9%) по отношению к другим изученным рекам. В р. Чёрной, наоборот, виден минимальный вклад в альгофлору бентосных форм (56%) и максимальный – планктонных и планк-то-бентосных форм по отношению к другим рекам (26 и 18%, соответственно). Альгофлоры р. Яйвы и Лёнвы занимают промежуточное положение по соотношению таксонов, принадлежащих к различным экологическим группам.
По географическому распространению во всех изученных реках наибольшую роль играют виды-космополиты (54–86%). Далее (11–37%) располагаются бореальные виды, доля аркто-альпийских видов невелика (0–10%). Наблюдается небольшая тенденция увеличения доли видов-космополитов во флоре участков, подверженных засолению. По сравнению с Камским водохранилищем [Беляева, 2013] доля бореальных и арктоальпийских видов в альгофлоре исследованных нами рек относительно высока.
В связи с нейтральными и нейтрально-щелочными условиями среды (рН = 7.16-8.18) в альгофлоре рек преобладают алкалифилы (33–77%) и индиффе-ренты (15–50%). В целом, альгофлоры р. Яйвы, Во-лима и Чёрной характеризуются сходным составом индикаторных таксонов по отношению к pH. В р. Лёнве, образующей собственную речную сеть, состав индикаторных таксонов водорослей и цианей отличается: здесь не отмечены ацидофилы, а доля алкали-филов возрастает за счёт снижения числа индифферентных форм (табл. 4).
По сапробности во всех водотоках многочисленны β -мезосапробные виды, в меньшем количестве в р. Волиме, Чёрной и Яйве – олигосапробы, а в р. Лёнве – α -мезосапробы.
Анализ состава индикаторных по сапробности представителей альгофлоры рек на отдельных участках не обнаруживает явных трендов. Сточные воды со шлакоотвалов калийного производства несут с собой неорганические вещества, поэтому изменения соотношений форм-индикаторов органического загрязнения не происходит. Так, максимальная совокупная доля олиго- и ксеносапробов (38–41%) выявлена на участках р. Волима выше устья р. Чёрной, выше моста на трассе «Романово-Володин Камень» и в заливе р. Волима («Волим 3», «Волим 6» и «Волим 9»).
Заключение
Общее видовое богатство водорослей и цианопрокариот участков рек, находящихся под влиянием стока со шлакоотвалов калийного производства и прилегающих к ним фоновых участков, достаточно велико – 300 видов и разновидностей, входящих в 106 родов, 59 семейств, 33 порядка и 7 отделов. Основную роль в видовом разнообразии играют диатомеи (73%). Наибольшее распространение получают бесшовные диатомовые из семейства Fragilariaceae : F. capucina , D. vulgare и U. ulna .
В эколого-географическом отношении альгоф-лора изученного района представлена преимущественно индифферентами и галофилами по отношению к галобности, индифферентами и алкали-филами по отношению к рН. В связи с особенностью формирования планктона малых рек, большинство отмеченных видов являются бентосными формами. По географическому распространению преобладают космополиты. По отношению к са-пробности представленные таксоны главным образом – β -мезосаробы, на втором месте почти во всех реках – олигосапробы, а в р. Лёнве – α -мезосапробы.
По показателю общности Чекановского-Серенсена альгоценозы, зарегистрированные на отдельных участках рек, делятся на несколько групп, согласованных также со степенью солёности вод этих участков.
В целом, на участках рек, испытывающих антропогенное засоление, снижается видовое богатство водорослей и цианей, величины родового коэффициента, и увеличивается доля галофилов.
Автор выражает искреннюю благодарность к.б.н., доценту Пермского государственного национального исследовательского университета М.А. Бакланову за предоставление гидрохимических данных изученных рек; сотрудникам Пермского отделения «ГосНИОРХ»: м.н.с. В.С. Котельниковой, рыбоводу С.П. Огородову и, в особенности, зам. директора, к.б.н. И.В. Поздееву за организацию экспедиций и помощь в сборе материала.
Список литературы Альгофлора рек Пермского края в условиях антропогенного засоления отходами калийного производства
- Баринова С.С., Медведева Л.А., Анисимова О.В. Бодоросли-индикаторы в оценке качества окружающей среды. М.: БННИИПрироды, 2000. 150 с
- Беляева П.Г. Фитоперифитон предгорной реки Сылва (бассейн Камы)//Ботанический журнал. 2004. Т. 89. № 3. С. 435-449
- Беляева П.Г. Особенности развития цианобактерий планктона Камского и Боткинского водохранилищ//Бестник Уральской медицинской академической науки. 2011. № 4/1. С. 22
- Беляева П.Г. Бидовой состав и структура фитопланктона Камского водохранилища//Бестник Пермского университета. Серия Биология. 2013. Бып. 3. С. 4-12
- Беляева П.Г., Поздеев И.В. Донные сообщества р. Чусовая (бассейн Камы)//Бестник Пермского университета. 2005. Был. 6. Биология. С. 103-108
- Буркова Т.Н. Характеристика фитопланктона высокоминерализованной реки Хара//Известия Пензенского гос. педагог. ун-та им. Б.Г. Белинского. 2011. № 25. С. 493-496
- Вайнштейн Б.А. Об оценке сходства между биоценозами//Биология, морфология и систематика водных организмов. Л.: Наука, 1976. 156-164 с
- Водоросли: справочник/С.П. Бассер, Н.Б. Кондратьева, Н.П. Масюк и др. Киев: Наук. думка, 1989. 608 с
- Генкал С.И., Беляева П.Г. Диатомовые водоросли (Centrophyceae) Камского водохранилища (Россия)//Альгология. 2011. T. 21, № 3. С. 312-324
- Генкал С.И., Охапкин А.Г. Диатомовые водоросли (класс Centrophyceae) в фитопланктоне Камских водохранилищ//Поволжский экологический журнал. 2010. № 3. С. 254-262
- Гончаров А.В. Фитопланктон малых рек Московского региона: автореф. дис.... канд. биол. наук. М., 1994. 18 с
- Горохова О.Г., Зинченко Т.Д. Фитопланктон высокоминерализованных рек Приэльтонья//Известия Самарского научного центра РАН. 2014. Т. 16, № 5-5. С. 1715-1721
- Грезе В.Н. Кормовые ресурсы рыб реки Енисей и их использование//Известия БНИИОРХ. 1957. Т. 41. С. 1-236 с
- Китаев С.П. Основы лимнологии для гидробиологов и ихтиологов. Петрозаводск, 2007. 395 с
- Комлев А.М., Черных E.A. Реки Пермской области. Пермь: Кн. изд-во, 1984. 214 с
- Мартыненко Н.А. Флора десмидиевых водорослей Урала на основе изучения коллекционных штаммов//Проблемы систематики и географии водных растений: материалы междунар. науч. конф. Борок, 2015. С. 57
- Определитель пресноводных водорослей СССР. М.: Сов. наука, 1951. Бып. 4. 620 с
- Определитель пресноводных водорослей СССР. М., 1953. Бып. 2. 651 с
- Определитель пресноводных водорослей СССР. М., 1954. Бып. 3. 188 с
- Определитель пресноводных водорослей СССР. М., 1954. Бып. 6. 212 с
- Определитель пресноводных водорослей СССР. М., 1955. Бып. 7. 281 с
- Определитель пресноводных водорослей СССР. М., 1959. Бып. 8. 230 с
- Определитель пресноводных водорослей СССР. Л.: Наука, 1980. Бып. 13. 248 с
- Определитель пресноводных водорослей СССР. Л., 1982. Бып. 11 (2). 624 с
- Определитель пресноводных водорослей СССР. Л., 1986. Бып. 10 (1). 360 с
- Охапкин А.Г. Структура и сукцессия фитопланктона при зарегулировании речного стока (на примере р. Болги и ее притоков): автореф. дис.... д-ра биол. наук. СПб., 1997. 50 с
- Пономарёва Ю.А. Структура и динамика потамо-фитопланктона реки Енисей в нижнем бьефе Красноярской ГЭС: дис.... канд. биол. наук. Красноярск, 2014. 152 с
- Селиванова Е.А., Немцева Н.В. Экологическо-трофическая характеристика микробиоценоза гипергалинного водоема на примере Соль-Илецкого озера Развал (Оренбургская область)//Проблемы региональной экологии. 2011. № 5. С. 79-86
- Тарасова Н.Г. Диатомовые водоросли Камских водохранилищ в 2005 г//IX школа диатомологов России и стран СНГ: морфология, систематика, онтогенез, экология и биогеография диатомовых водорослей: тез. докл. Борок, 2005. С. 64-65
- Таусон А.О. Бодные ресурсы Молотовской области. Молотов: ОГИЗ, 1947. 321 с
- Третьякова С.А., Головачева С.И., Батова Е.М. Фитопланктон//Биология Боткинского водохранилища. Иркутск, 1988. С. 26-36
- Формирование загрязнения поверхностных вод в районе разработки месторождения калийных солей/Н.Г. Максимович и др.//Современные проблемы водохранилищ и их водосборов: тр. V Междунар. науч.-практ. конф. Пермь, 2015. Т. 2. С. 120-123
- Фролова Г.И. Фитопланктон малых рек Ярославской области: автореф. дис.... канд. биол. наук. М., 2004. 25 с
- Царенко П.М. Краткий определитель хлорококковых водорослей Украинской ССР. Киев: Наук. думка, 1990. 208 с
- Царенко П.М. Номенклатурно-таксономические изменения в системе «зеленых» водорослей//Альгология. 2005. T. 15, № 4. С. 459-467
- Царенко П.М. Рекомендации по унификации цитирования фамилий авторов таксонов водорослей//Альгология. 2010. № 20. С. 86-121
- Diatom Monographs. Gantner, 2003. Vol. 4. 656 p
- Diatoms of Europe: Navicula sensu stricto, 10 genera separated from Navicula sensu lato, Frustulia Vol. 2./H. Lange-Bertalot. Ruggell: ARG Gant-ner Verlag KG, 2001. 526 p.
- Diatoms of Europe. Diatoms of the European inland waters and comparable habitats. Vol. 4. Cym-bopleura, Delicata, Navicymbula, Gomphocym-bellopsis, Afrocymbella/H. Lange-Bertalot. ARG Gantner Verlag KG, 2003. 530 p
- Diatoms of Europe. Diatoms of the European inland waters and comparable habitats. Vol. 5. Amphora sensu lato/H. Lange-Bertalot. Ruggell: ARG Gantner Verlag KG, 2009. 916 p
- Diversity and ecology of algae from the Nahal Qishon river, northern Israel/S.S. Barinova, O.V. Anis-simova, E. Nevo et al.//Plant biosystems. 2004. № 138 (3). P. 245-259
- Round F.E., Crawford R.M., Mann D.G. The Diatoms. Biology and morphology of the genera. Cambridge: Cambridge University Press, 1990. 747 p.
- Round F.E., Bukhtiyarova L. Four new genera based on Achnanthes (Achnanthidium) together with a re-definition of Achnanthidium//Diatom Research. 1996. Vol. 11 (2). P. 345-361
- Sorensen T. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content//Kongelige Danske Videnskaber-nes Selskab. Biol. krifter. 1948. Vol. 4. P. 1-34
- Taxonomic classification of cyanoprokaryotes (cyano-bacterial genera) 2014, using a polyphasic approach/J. Komarek, J. Kastovsky, J. Mares//Preslia. 2014. Vol. 86, № 4. P. 295-335
- Ventosa A., Arahal D.R. Microbial life in the Dead Sea//Enig-matic Microorganisms and Life in Extreme Enviroments. Dordrecht; Boston; London: Kluwer Academic Publishers, 1999. P. 357-368
