Альгофлора родников Ижевска

Автор: Госькова М.С., Науменко Н.И., Алалыкин А.С.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 4, 2025 года.
Бесплатный доступ
Приводится таксономический состав альгофлоры источников г. Ижевска и его окрестностей, где было выявлено 69 видов водорослей из 10 классов 5 отделов. В списке видов выделяется ядро родниковой альгофлоры, включающее специфичные для эукренов реофильные водоросли (Odontidium mesodon, Meridion circulare, Ulnaria ulna, Achnanthidium minutissimum, Gomphonema parvulum), а также часто встречающиеся Audouinella pygmaea, Phormidium umbilicatum, Frustulia crassinervia и Nitzschia linearis. Низкая степень видового сходства родников обусловлена большими различиями в составе редко отмечаемых видов, среди которых выделяется группа бриофильных видов, ассоциированных с произрастающими у выходов родниковых вод мхами (Odontidium mesodon, Frustulia crassinervia). В списке отмечены редкие виды Dactylococcopsis rhaphidioides f. falciformis (Cyanophyceae) и Gomphonema liyanlingae (Bacillariophyceae). Наибольшим количеством видов в списке представлен класс Bacillariophyceae – 51 вид из 27 родов, из которых наиболее богаты видами роды Nitzschia (6 видов), Pinnularia (5 видов), Cymbella и Gomphonema (по 4 вида). Также приведены сведения о некоторых физико-химических характеристиках воды родников: температуре, кислотности, общей минерализации, содержании нитратов, сульфатов, кремния и хлоридов. Обсуждается экологическая значимость родников в урбанизированном ландшафте как уникальных водных местообитаний.
Родники, подземные воды, альгофлора, Ижевск
Короткий адрес: https://sciup.org/147252641
IDR: 147252641 | УДК: 627.12[556.114+582.261/.279](470.51-25) | DOI: 10.17072/1994-9952-2025-4-338-350
Algal flora in the springs of Izhevsk
This article provides a taxonomic list of spring algal flora in the city of Izhevsk with its area, which includes 69 species of 5 divisions. In the list of algal species, there is a core assemblage of algal flora which consists of rheophilic algae typical for eucrenes (Odontidium mesodon, Meridion circulare, Ulnaria ulna, Achnanthidium minutissimum, Gomphonema parvulum) and also frequently occurring Audouinella pygmaea, Phormidium umbilicatum, Frustulia crassinervia and Nitzschia linearis Low compositional similarity in springs is a result of big differences in composition of species with low frequency of registering, among them there is a group of bryophylic algae, which are associated with mosses growing near springs (Odontidium mesodon, Frustulia crassinervia). There are also rare species Dactylococcopsis rhaphidioides f. falciformis (Cyanophyceae) and Gomphonema. liyanlingae (Bacillariophyceae) on the list. The most species of the list belong to Bacillariophyceae (51 species of 27 genera) and among them the most species are from genera Nitzschia (6 species), Pinnularia (5 species), Cymbella (4 species), Gomphonema (4 species). Also the data about some spring water characteristics, such as temperature, pH, total dissolved solids, concentration of nitrates, sulfates, silicon and chlorides are provided. Ecological role of springs in urban environment as unique water habitat is also discussed.
Текст научной статьи Альгофлора родников Ижевска
Родники являются характерной чертой ландшафта г. Ижевска (на его территории насчитывается 46 родников). Все они происходят из нижнеустьинского терригенного водоносного горизонта, залегающего под чехлом четвертичных отложений и выходящего на поверхность в нижних частях склонов рек. Водовмещающие породы горизонта – песчаники, известняки и мергели, водоупорные слои сложены глинами и алевролитами. Мощность водовмещающей толщи колеблется от 2.1 до 24 м, питание преимущественно атмосферное, водообильность очень изменчива при среднем значении дебита родников 0.5–1.0 л/с [Елькин, Рысин, 1998; Родники Ижевска, 2000]. Все рассмотренные родники по характеру выхода являются реокренами. Многие ижевские родники имеют сливы, что оставляет очень мало пригодного для развития альгофлоры пространства; часть родников заключены в постройки и сильно затемнены, что также не способствует развитию богатой альгофлоры. Отсутствие охранного статуса городских родников привело к исчезновению 16 из них за последние два десятилетия [Экология и природопользование…, 2018], в то же время большинство оставшихся родников продолжают использоваться населением в качестве источников питьевой воды при недостаточном информировании о ее качестве. Сравнение нынешнего состояния каптажей с фотографиями 1990-х гг. из монографии «Родники Ижевска» [2000] показывает деградацию большинства прилегающих к родникам территорий.
В монографии, обобщающей данные о характеристиках и охране хозяйственно-питьевых вод Удмуртской АССР [Наумов, 1978], родниковые воды Ижевска по химическому составу характеризовались преимущественно как гидрокарбонатные с умеренной минерализацией.
Исследования воды большинства городских родников и прилегающих к ним почв по некоторым химическим параметрам периодически проводились с 1990-х гг. [Родники Ижевска, 2000; Исаев и др., 2004; Рылова и др., 2006; Рылова и др., 2008; Владыкина и др., 2016; Экология и природопользование …, 2018; Раянова, 2021]; в 2004–2020 гг. изучалась бриофлора некоторых родников Ижевска [Рубцова, 2024]. Однако населяющая родники альгофлора в перечисленных публикациях не рассматривалась. Целью нашей работы было выявление таксономического состава водорослевой флоры ряда ижевских родников и дополнение сведений о физико-химических характеристиках их вод.
Материалы и методы
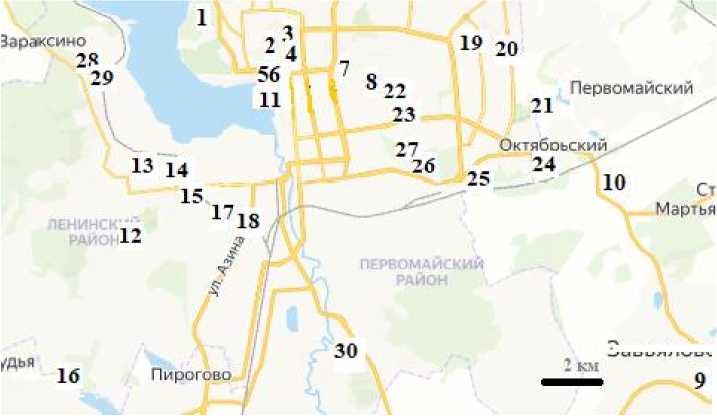
Сбор проб воды и альгологического материала проводился с 22 по 29 августа 2023 г. в следующих точках (номера в скобках соответствуют номерам родников, указанным в монографии «Родники Ижевска» [2000]) (рис. 1): 1. Важнин ключ (№ 50); 2. Подборенский 1 (№ 59); 3. Подборенский 2 (№ 55); 4. Подборенский 3 (№ 48); 5. Подборенский 4 (№ 46); 6. Подборенский 5 (№ 47); 7. Родник имени Д. А. Пригова (№ 40); 8. Козий родник (№ 41); 9. Пугачёвский родник (село Завьялово); 10. Артезианская скважина в деревне Кабанихе; 11. Родник на набережной (№ 51); 12. Родник в Парке им. Е.Ф. Драгунова; 13. Ключ 1; 14. Ключ 2; 15. Родник № 13 (№ 13); 16. Артезианская скважина в микрорайоне Шунды (ниже улицы Учхозной); 17. Родник № 11 (№ 11); 18. Казанский родник (№ 10); 19. Чемошурский 1 (№ 19); 20. Чемошурский 2 (№ 23); 21. Родник в Ярушкинском парке; 22. Карлутский 1 (№ 42); 23. Карлутский 2, 3 (№ 33, № 34); 24. Родник в 3-м микрорайоне (северо-западнее СНТ «Пенсионер»); 25. Карлутский 4 (№
28); 26. Карлутский 5 (№ 29); 27. Карлутский 6 (№ 30); 28. Малиновский 1 (№ 3); 29. Малиновский 2 (№ 5); 30. Родник в микрорайоне Медведево (№ 25).
Завьялове
Рис. 1. Карта-схема точек пробоотбора (номера точек отбора проб соответствуют приведенным в тексте)
[Scheme of sample sites (numbers of sites coincide with those in the text)]
Всего были собраны и проанализированы пробы из 28 родников и 2 артезианских скважин. Определяемые физико-химические параметры включали температуру воды, кислотность, общую минерализацию, содержание нитратов, сульфатов, кремния и хлоридов.
Показатель общей минерализации воды (total dissolved solids, TDS) измерялся карманным солемером T.D.S Meter 3 Hold, pH воды – карманным pH-метром (Water test, Китай). Температуру (T) воды измеряли ртутным термометром с минимальным делением в 1°C . Химический анализ проб воды осуществлялся фотометрическими методами (при помощи спектрофотометра «UNICO 2100») для нитратов (ПНД Ф 14.1:2:4.4-95) и кремния (ПНД Ф 14.1:2:4.215-06), турбидиметрическим методом для сульфатов (ПНД Ф 14.1:2.159-2000), аргентометрическим методом для хлоридов (ПНД Ф 14.1:2:3.96-97).
Перифитон отбирали с прилегающих к выходам родников субстратов (песка, камней, бетонных, металлических или деревянных водостоков). Анализ видового состава осуществлялся с помощью светового микроскопа Zeiss Axiostar plus (Carl Zeiss, Германия) и частично электронно-сканирующего микроскопа InspectS50 по принятым определителям [Виноградова и др., 1980; Царенко, 1990; Komarek, Anagnostidis, 2005; Куликовский и др., 2016] и электронной базе Diatoms of North America [2024]. Для приготовления препаратов диатомовых водорослей были использованы перекись водорода (37%) и среда Naphrax. Номенклатура таксонов приведена в соответствии с электронной базой данных AlgaeBase ]2023]. Степень сходства флор выражалась индексом Жаккара.
Результаты и их обсуждение
Территориально в списке исследованных родников можно выделить пять групп (в скобках указаны номера точек пробоотбора):
-
1) подборенские (2–6): родники этой группы, принадлежащие бассейну р. Подборенка, расположены компактно и сходны по составу воды и альгофлоры; t 9.5–11.0°C, pH 7.0–7.3, TDS 490–600, NO 3 40–57 мг/дм3, SO 4 61–72 мг/дм3, Si 10–12 мг/дм3, Cl 78–92 мг/дм3; здесь отмечены максимальные значения содержания сульфатов в воде;
-
2) карлутские (7–8, 22–23, 25–27): родники этой группы, принадлежащие бассейну р. Карлутка, расположены на протяженной территории и более разнообразны по составу воды, чем родники первой группы; t 7.0–12.0°C, pH 6.9–7.4, TDS 360–640, NO 3 50–95 мг/дм3, SO 4 28–58 мг/дм3, Si 8.0–11.5 мг/дм3, Cl 37–106 мг/дм3;
-
3) чемошурские (19–21): родники этой группы принадлежат бассейну р. Чемошурка; t 7.0–10.5°C, pH 7.0–7.2, TDS 300–490, NO 3 18–25 мг/дм3, SO 4 30–51 мг/дм3, Si 12.0–13.5 мг/дм3, Cl 53–101 мг/дм3;
-
4) правобережные (15, 17–18): группа родников к югу от Ижевского водохранилища; t 8.5–10.0°C, pH 6.9–7.0, TDS 500–570, NO 3 52–105 мг/дм3, SO 4 48–68 мг/дм3, Si 14.5–19.0 мг/дм3, Cl 67–102 мг/дм3 (очень высокие концентрации нитратов (до 177 мг/дм3) отмечались для родников этой группы еще со второй половины 1990-х гг. [Родники Ижевска, 2000]);
-
5) Малиновские (28–29): родники, принадлежащие бассейну р. Малиновка; t 7.0–7.5°C, pH 7.1–7.2, TDS 260–360, NO 3 36–37 мг/дм3, SO 4 19–25 мг/дм3, Si 13.5–14.0 мг/дм3, Cl 17–46 мг/дм3.
Прочие родники расположены одиночно и удаленно от этих групп. В целом значения pH воды находятся в диапазоне 6.9–9.0, TDS – 130 (точка 9) – 640 (точка 25 ), концентрации NO 3 в диапазоне <1 (точки 10 и 16 ) – 105 мг/дм3 (точка 18 ) (ПДК нитратов (по СанПиН 1.2.3685-21) превышена в 16-ти точках, наиболее значительно в воде родников карлутской и правобережной групп), SO 4 в диапазоне 9 (точка 9 ) – 72 мг/дм3 (точка 4 ), Si в диапазоне 4.5 (точка 10 ) – 19 мг/дм3 (точка 15 ), Cl в диапазоне <1 (точка 16 ) – 147 мг/дм3 (точка 11 ).
Сравнение с литературными данными по родникам других городских территорий [Назаров, 2002; Буймова, 2006; Зуева, Китаев, 2010; Кузнецова, 2016; Орлов, 2016; Лукашевич, Чернышова, 2018; Тизян, Скугорева, 2020; Пасечник и др., 2022; Романова, Большаник, 2022; Соболева, 2023] показывает, что физические параметры и концентрации макроэлементов в воде ижевских родников в целом не выходят за характерные для этого типа водных объектов пределы, хотя в воде родников северо-западной части города (точки 12, 13, 15 ) заметно высокое содержание кремния. Вода артезианских скважин (точки 10 и 16 ) отличается минимальным содержанием нитратов и кремния и низким содержанием хлоридов.
Список видов водорослей исследованных родников
Обозначения обилия и встречаемости вида (символ слева: + – единичные клетки, r – мало, f – в значительном количестве, m – в массе; символ справа: P – Подборенская группа родников, K – Карлутская группа, B – Правобережная группа, C – Чемошурская группа, M – Малиновская группа, + – более, чем в двух группах).
Cyanobacteriophyta
Cyanophyceae
Chroococcales
Chroococcaceae
Chroococcus sp. r+
Dactylococcopsis rhaphidioides f. falciformis Printz (рис. 3c) fP
Microcystaceae
Merismopedia sp. rK
Oscillatoriales
Microcoleaceae
Phormidiochaete balearica (Bornet et Flahault ex Forti) Komárek fPB
Oscillatoriaceae
Phormidium umbilicatum Gomont f+
Coleofasciculales
Coleofasciculaceae
Symploca sp. rB
Heterokontophyta
Bacillariophyceae
Achnanthales
Achnanthaceae
Achnanthidium minutissimum (Kützing) Czarnecki (рис. 2q) m+
Ach. sp. (рис. 3b) f+
Cocconeidaceae
Cocconeis euglypta Ehrenberg (рис. 2ad) r+
-
C. pediculus Ehrenberg fK
Achnanthidiaceae
Planothidium frequentissimum (Lange-Bertalot) Lange-Bertalot (рис. 2s) fPK
P. granum (Hohn and Hellerman) Lange-Bertalot (рис. 2k) rP
P.lanceolatum (Brébisson ex Kützing) Lange-Bertalot (рис. 2u) fPK
Psammothidium chlidanos (Hohn and Hellerman) Lange-Bertalot (рис. 2m) fCP
P. subatomoides (Hustedt) Bukhtiyarova et Round (рис. 2r) fP
Bacillariales
Bacillariaceae
Denticula tenuis Kützing (рис. 2t) fK
Nitzschia clausii Hantzsch f+
Nitzschia dissipata (Kützing) Rabenhorst rP
N. fonticola (Grunow) Grunow (рис. 2aa) rB
N. linearis Smith. fPK
N. palea (Kützing) Smith (рис. 2z) f+
N. sigmoidea (Nitzsch) Smith r+
Tryblionella salinarum (Grunow) Pelletan fP
Cymbellales
Anomoeoneidaceae
Anomoeoneis sphaerophora Pfitzer r+
Cymbellaceae
Cymbella hantzschiana Krammer (рис. 2a) mP
-
C. hustedtii Krasske (рис. 2b) mK
-
C. neogena (Grunow) Krammer r+
-
C. vulgata Krammer (рис. 2c) fP
Witkowskia abiskoensis (Hustedt) Kulikovskiy, Glushchenko, Mironov et Kociolek (рис. 2p) rPBK Gomphonemataceae
Gomphonema acuminatum Ehrenberg (рис. 2h) f+
-
G. capitatum Ehrenberg (рис. 2g) r+
-
G. liyanlingae Metzeltin et Lange-Bertalot (рис. 2i). rP
Gomphonema parvulum (Kützing) Kützing sensu lato (рис. 2f). Как показано в исследовании D.T. Rose [2008], этот таксон должен по крайней мере частично включать G. gracile , т. к. инициальные клетки G. parvulum морфологически полностью соответствуют второму виду. Кроме того, некоторые выделенные в отдельные виды формы, такие как G. exilissimum , могут все же являться экоморфами (вид описывается как приуроченный к олиготрофным условиям с низкой минерализацией, в то время как G. parvulum описывается как эврибионт). В наших образцах морфологические признаки отнесенных к G. parvulum клеток значительно варьировали, что может быть скорее выражением возрастной структуры популяции, чем соседства различных форм. m+
Rhoicospheniaceae
Rhoicosphenia abbreviata (Agardh) Lange-Bertalot f+
Eunotiales
Eunotiaceae
Eunotia bilunaris (Ehrenberg) Schaarschmidt (рис. 2ab) fP
Fragilariales
Fragilariaceae
Fragilaria radians (Kützing) Williams et Round fP
-
F. vaucheriae (Kützing) Petersen (рис. 2x) fPK
Odontidium mesodon (Ehrenberg) Kützing (рис. 2ac, рис. 3a) m+
Staurosiraceae
Staurosira tabellaria (Smith) Leuduger-Fortmorel (рис. 2n) fP
Licmophorales
Ulnariaceae
Ulnaria ulna (Nitzsch) Compère m+ Naviculales
Naviculaceae
Caloneis silicula (Ehrenberg) Cleve fK
Navicula cf. vaneei rK
Diploneidaceae
Diploneis boldtiana Cleve (рис. 2j) rPK
Amphipleuraceae
Frustulia crassinervia (Brébisson ex W.Smith) Lange-Bertalot et Krammer fK
Neidiaceae
Neidiomorpha binodis (Ehrenberg) M. Cantonati, Lange-Bertalot et N. Angeli (рис. 2e) +K Pinnulariaceae
Pinnularia canadodivergens Kulikovskiy, Lange-Bertalot et Metzeltin rKC
P. isselana Krammer +C
P. neomajor Krammer r+
P. ovata Krammer +K
P. subrostrata (Cleve) Cleve (рис. 2o) rK
Stauroneidaceae
Stauroneis phoenicenteron (Nitzsch) Ehrenberg rKB
S. smithii Grunow (рис. 2d) rK
Rhabdonematales
Tabellariaceae
Meridion circulare (Greville) Agardh (рис. 2y) m+
Surirellales
Surirellaceae
Surirella brebissonii Krammer et Lange-Bertalot fPK
S. lacrimula English (рис. 2w) rP
S. sp. (рис. 2v) +K
Thalassiophysales
Catenulaceae
Amphora pediculus (Kützing) Grunow (рис. 2l) f+
Coscinodiscophyceae
Melosirales
Melosiraceae
Melosira varians Agardh f+
Xanthophyceae
Tribonematales
Tribonemataceae
Tribonema sp. +K
Rhodophyta
Florideophyceae
Acrochaetiales
Audouinellaceae
Audouinella pygmaea (Kützing) Weber Bosse (рис. 3d) mPB
Chlorophyta
Chlorophyceae
Chaetophorales
Fritschiellaceae
Stigeoclonium sp. fK
Trebouxiophyceae
Microthamniales
Microthamniaceae
Microthamnion strictissimum Rabenhorst fB
Klebsormidiophyceae
Klebsormidiales
Klebsormidiaceae
Klebsormidium fluitans (Gay) Lokhorst fK
Ulvophyceae
Ulotrichales
Ulotrichaceae
Ulothrix sp . fK
Cladophorales
Cladophoraceae
Cladophora sp. f+
Charophyta
Zygnematophyceae
Desmidiales
Desmidiaceae
Cosmarium sp. rK
Closteriaceae
Closterium lunula Ehrenberg et Hemprich ex Ralfs fM
Spirogyrales
Spirogyraceae
Spirogyra sp. rKM
Zygnematales
Zygnemataceae
Mougeotia sp. rK
Всего в ходе исследования нами было выявлено 69 видов водорослей из 40 родов, 37 семейств и 9 классов; из них к Heterokontophyta принадлежат 53 вида (Bacillariophyceae – 51 вид из 27 родов, 21 семейства и 10 порядков, Coscinodiscophyceae – 1 вид, Xanthophyceae – 1 вид), Chlorophyta – 5 видов из 4 семейств и 4 классов, Charophyta – 4 вида из 3 семейств и 3 порядков, Rhodophyta – 1 вид, Cyanobacteria – 6 видов из 5 семейств и 3 порядков. Наибольшее видовое разнообразие имели порядки Naviculales (12
видов) и Cymbellales (11 видов), на уровне семейства – Bacillariaceae (8 видов), среди родовых таксонов – Nitzschia (6 видов), Pinnularia (5 видов), Cymbella и Gomphonema (по 4 вида).
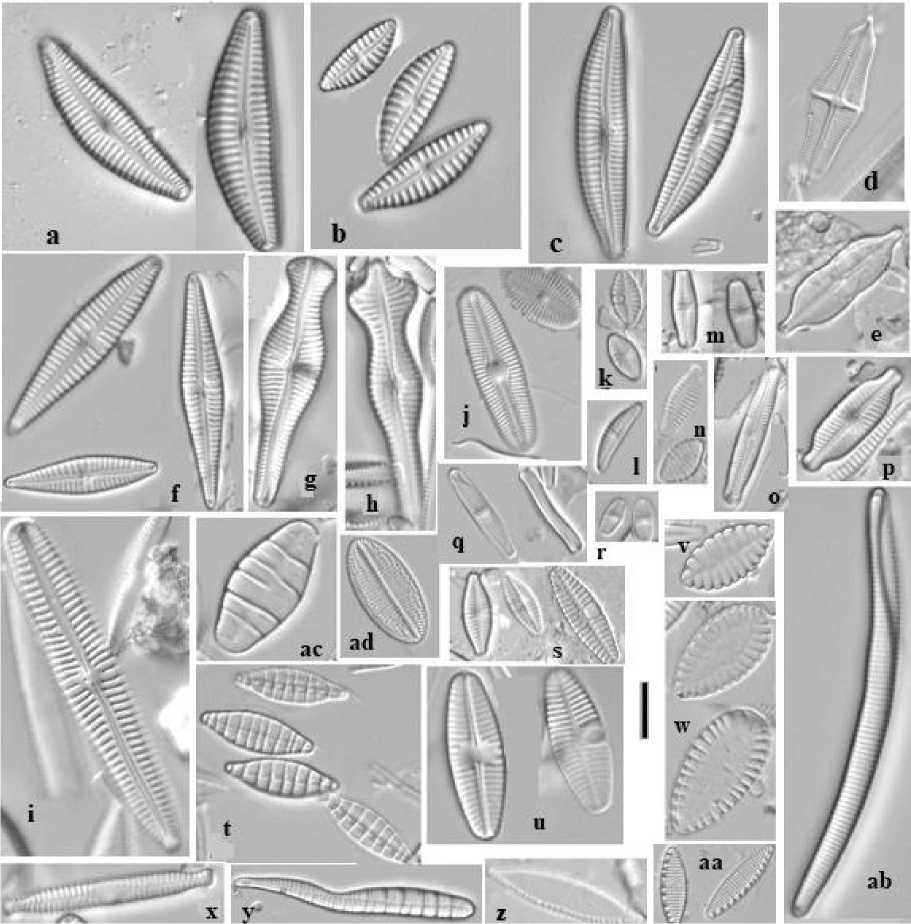
Рис. 2 . Диатомовые водоросли родников Ижевска [Diatoms of Izhevsk springs]:
a – Cymbella hantzschiana ; b – C. hustedtii ; c – C. vulgata ; d – Stauroneis smithii ; e – Neidiomorpha binodis ; f – Gomphonema parvulum ; g – G. capitatum ; h – G. acuminatum ; i – G. liyanlingae ; j – Diploneis boldtiana ; k – Planothidium granum ; l – Amphora pediculus ; m – Psammothidium chlidanos ; n – Staurosira tabellaria ; o – Pinnu-laria subrostrata ; p – Witkowskia abiskoensis ; q – Achnanthidium minutissimum ; r – Psammothidium subatomoides ; s – Planothidium frequentissimum ; t – Denticula tenuis ; u – Planothidium lanceolatum ; v – Surirella sp.; w – S. lac-rimula ; x – Fragilaria vaucheriae ; y – Meridion circulare (тератоформа); z – Nitzschia palea ; aa – Nitzschia fonti-cola ; ab – Eunotia bilunaris ; ac – Odontidium mesodon ; ad – Cocconeis euglypta . Масштабная линейка: 10 мкм [Scale bar: 10 mcm]
Комплекс наиболее часто встречающихся видов водорослей – Odontidium mesodon , Meridion circulare , Ulnaria ulna , Achnanthidium minutissimum , Gomphonema parvulum (два первых указанных вида выявлены нами вместе в 13 точках, все перечисленные – в 4 точках). Также часто встречаются Phormidium umbilica-tum (в 12 точках), Audouinella pygmaea , Frustulia crassinervia (в 9 точках) и Nitzschia linearis (в 8 точках), однако большинство выявленных видов обнаружены только в одной или двух точках. Эта особенность отмечалась у родниковых флор других территорий [Cantonati et al., 2012a , 2012b]. Наибольшее разнообразие видового состава было зафиксировано в пробах из точек 6 «Подборенский 5» и 23 «Карлутский 2, 3» (по 15 видов), наименьшее – в точках 25 «Карлутский 4» и 16 «Артезианская скважина в микрорайоне Шунды» (по 4 вида), не считая точки 8 «Козий родник», где водорослевые обрастания полностью отсут- 344
ствовали по причине проведения строительных работ. Сравнительный анализ видового состава водорослей групп родников и отдельных родников показал невысокую степень сходства: значение коэффициента Жаккара не превышает 0.5. Низкая степень сходства связана с однократно встреченными видами, однако по представленности шести вышеуказанных часто встречающихся видов группы родников полностью сходны, а отдельные (не включенные в группы) родники отличаются отсутствием одного или двух из этих видов. Из наиболее часто встречающихся видов O. mesodon, M. circulare и A. pygmaea являются типичными криореофилами; A. minutissimum , U. ulna и G. parvulum обладают более широким экологическим спектром: трофические характеристики этих видов находятся в диапазоне от стенобионтных оли-готрофов до мезотрофов.
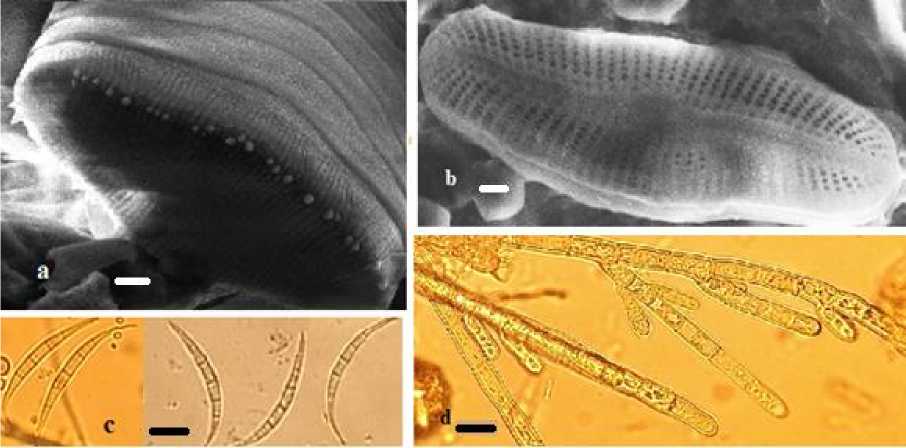
Рис. 3 . Водоросли родников Ижевска [Algae of Izhevsk springs]:
a – Odontidium mesodon, СЭМ; b – Achnanthidium sp., СЭМ; c – Dactylococcopsis rhaphidioides, СМ; d – Audoui-nella pygmaea, СМ. Масштабная линейка: a, b – 1 мкм; c, d – 10 мкм [Scale bar: a, b – 1 mcm; c, d – 10 mcm]
Значительные различия физических характеристик родников (освещенность, характер прилегающего грунта, дебит), влияющие на развитие альгофлоры, различное число родников в группе (от 2 до 7) и степень территориальной компактности определяют степень видового сходства их альгофлор.
Родники являются уникальными по постоянству среды водными местообитаниями, где встречаются редкие виды и специфические сообщества водорослей [Cantonati et al., 2012b; Delgado et al., 2013]. Сравнение с данными по некоторым альпийским и балканским родникам [Levkov et al., 2005; Fránková et al, 2009; Cantonati et al., 2012b; Kamberović at al., 2019] показывает, что несколько родов и видов можно считать типичными представителями европейской родниковой флоры. Так, O. mesodon является обитателем богатых кремнием и кальцием холодных реокренов и вместе с M. circulare составляет характерное сообщество эукренов (выходов родниковых вод) [Cantonati et al., 2012b], которое и наблюдалось нами в большинстве исследованных проб. A. minutissimum и другие виды рода Achnanthidium часто доминируют в полноводных алкалинных водотоках на карбонатном субстрате; P. frequentissimum, G. parvulum (f. exil-issimum ) и D. tenuis также являются преимущественно родниковыми, гигропетрофильными видами [ibid]. Виды рода Audouinella представляют собой важный компонент флоры в затененных родниках с повышенным содержанием нитратов. F. crassinervia и виды рода Eunotia считаются связанными с хорошо освещенными низкоалкалинными и низкоминерализованными родниками [ibid], однако в наших пробах бриофильный вид F. crassinervia не проявлял связи с этими условиями. Среди самых характерных для родников родов, которые были представлены и в наших пробах, выделяются Achnanthidium , Planothidium, Psammothidium, Gomphonema , Cymbella , Meridion , Odontidium , Audouinella (при этом большая доля биомассы в исследованных нами родниках принадлежит Ulnaria , которая является скорее «речным» родом [Peeters et al., 2017]); все они – типичные представители перифитона.
Заключение
Таким образом, в альгофлоре исследованных родников можно выделить совокупность типичных видов, составляющих ее ядро. Большинство выявленных видов встречались только в одном или нескольких родниках. Среди них были как характерные представители родниковых флор, чувствительные к характеру субстрата и составу воды, так и эвритопные виды.
Поскольку скорость течения и субстрат играют важнейшую роль в формировании перифитонного сообщества водорослей в числе абиотических условий среды, состояние естественных выходов подземных вод оказывается не менее значимым фактором в формировании видового состава альгофлоры, чем состав воды и температура. Сохранение родников в качестве уникальных водных местообитаний и городских достопримечательностей возможно только при организации охранных мероприятий [Родники Ижевска, 2000; Гагарина, 2012; Гагарина, Юнусова, 2015; Экология и природопользование…, 2018], однако имеющиеся предложения, включающие регулярную чистку каптажных камер и контроль за состоянием родников и благоустройством их территорий, представляются нам направленными на сохранение родников только как дополнительных источников питьевого водоснабжения, а не родниковых экосистем, поскольку эти меры недостаточно способствуют сохранению остатков естественной родниковой альгофлоры. Тем не менее, придание некоторым родникам статуса особо охраняемых природных территорий местного значения, вероятно, позволило бы сохранить эти экосистемы в качестве природных местообитаний.
