Альтерационные, компенсаторно-приспособительные и адаптационно-пластические структурные изменения нервов конечности при дистракционном остеосинтезе и нейротензии

Автор: Борисова И.В.
Журнал: Гений ортопедии @geniy-ortopedii
Рубрика: Конкурсные работы молодых ученых
Статья в выпуске: 3, 2006 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/142120934
IDR: 142120934
The alterative, compensatory-and-adaptive, adaptative-and-plastic structural changes of limb nerves for distraction osteosynthesis and neurotension
Текст статьи Альтерационные, компенсаторно-приспособительные и адаптационно-пластические структурные изменения нервов конечности при дистракционном остеосинтезе и нейротензии
Анализ литературы свидетельствует, что в последние десятилетия проводятся интенсивные исследования структурных изменений нервов конечностей при скелетном вытяжении, дистракционном остеосинтезе, нейротензии экспандером Радована [1-4, 10-14, 16]. Однако комплексные морфологические исследования, включающие применение световой и электронной микроскопии, а также сочетание феноменологических описаний с применением популяционно-статистического подхода для оценки изменений нервных волокон, в доступной литературе отсутствуют. Цель исследования - комплексная оценка структурных изменений проводниковой части нервов конечностей при дистракционном остеосинтезе и нейротензии по методу Илизарова.
МАТЕРИАЛЫ И МЕТОДЫ
Эксперименты проведены на 17 взрослых беспородных собаках. В первой серии опытов (n=14) изучали структурную реорганизацию седалищного нерва при дистракционном остеосинтезе бедренного сегмента тазовой конечности (рис. 1-1) модифицированным аппаратом Илизарова, защищённым свидетельством на полезную модель [8]. Во второй серии (n=3) исследовали изменения нервов при тракции мягких тканей в поперечном направлении нейротензии по методу Илизарова (рис. 1-2): операции включали имплантацию опорного элемента тракционного устройства (по а.с. № 1693744) в ложе седалищного нерва. В первой серии животных выводили из опыта в конце дистракции (удлинение бедра на 14-17,5 %), в различные сроки фиксации и после снятия аппарата, во второй серии - после удлинения седалищного нерва на 25-30 % и окончания периода фиксации. Содержание животных, оперативные вмешательства и эвтаназию осуществляли согласно приказу МЗ СССР № 755 1977 г.
Для исследования брали участки седалищного нерва оперированной и контралатеральной конечностей. Часть материала погружали в жидкий азот и в криостате разлагали на поперечные срезы, в которых кальций-кобальтовым методом выявляли активность АТФ-азы по Padycula-Herman [6], продольные криостатные срезы им-прегнировали серебром по методике А.К. Коломийцева с соавт. (1981) [5] и по Рассказовой. Основную часть материала помещали в глутаральдегидную смесь, постфиксировали в тетраоксиде осмия и заливали в эпоксидные блоки. Полутонкие (толщиной 0,5-1,0 мкм) срезы большой площади [7] окрашивали толуидиновым синим или метиленовым синим-основным фуксином по Уикли. Ультратонкие поперечные срезы контрастировали уранил-ацетатом и цитратом свинца, изучали в просвечивающем электронном микроскопе JEM-100B («Jeol», Япония) при ускоряющем напряжении 40 kV. Изображения для последующего анализа фотографировали на пластинки. Ультраструктурное исследование проведено совместно с д.м.н. М.М. Щудло.
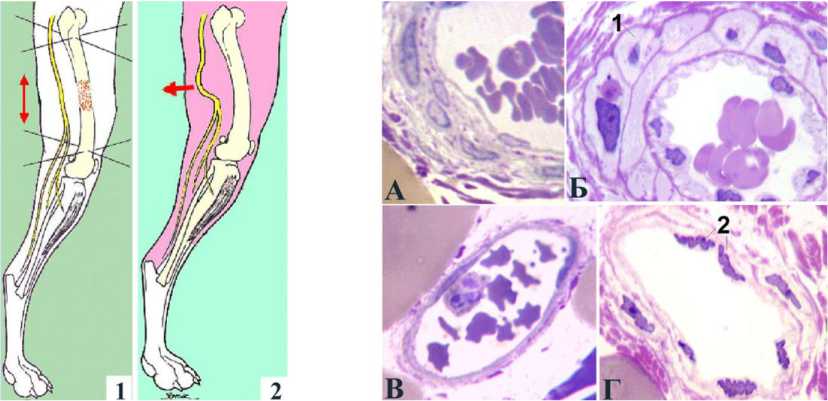
Рис. 2. Эпиневральные сосуды удлинённого седалищного нерва собаки при дистракционном остеосинтезе ( А, В ) и тракции в поперечном направлении ( Б, Г ). А, Б артерии, В, Г вены. Поперечные полутонкие срезы, окраска по Уикли, увеличение (УВ) 1250 . 1 продольноориентированная ГМК, 2 клетки с гофрированными ядрами
Рис. 1. Схема экспериментов: 1 удлинение седалищного нерва собаки при дистракционном остеосинтезе. 2 удлинение нерва при тракции мягких тканей в поперечном направлении
Микроскопические исследования полутон-ких срезов дополняли компьютерной морфометрией. Для оцифровки изображений использовали лупу и большой исследовательский микроскоп фирмы “Opton” (Германия), совмещённый с аппаратно-программным комплексом “ДиаМорф” (Москва). Формировали выборку из 30-46 полей зрения при инструментальном уве- личении 1250х. Подсчитывали численную плотность мякотных волокон, оценивали их распределение по диаметру. Для каждого волокна рассчитывали число G как отношение диаметра осевого цилиндра к диаметру волокна [15]. Используя аппроксимационные средства Microsoft Excel-97, строили диаграммы зависимости числа G от диаметра мякотных волокон.
РЕЗУЛЬТАТЫ
В первой серии опытов при исследовании криостатных срезов визуально определяется увеличение количества функционирующих микрососудов в эпиневрии и эндоневрии по сравнению с контралатеральным и интактным нервом. Петли эпиневрального сосудистого русла удлинённого нерва в отличие от интактного и контралатерального приобретают вытянутую форму. Сосуды артериального и артериолярного типа имеют широко раскрытые просветы. При микроскопии полутонких срезов в них определяется сохранность внутренней эластической мембраны, гипертрофия мышечной оболочки с преимущественно косоциркулярной ориентацией гладкомышечных клеток (ГМК) (рис. 2, А). Количество волокнистых и клеточных элементов в адвентиции увеличено.
В опытах с нейротензией наряду с продольной переориентацией петель эпиневрального сосудистого русла отмечается ещё более выраженное, чем в первой серии, экспансивное ремоделирование. Эпиневральные артерии имеют типичное для растущих и функционирующих в условиях гипердинамического кровотока сосудов строение: в медии увеличиваются размеры и количество ГМК, нередки миоциты, ориентированные продольно (рис. 2, Б). В стенках вен об- наруживаются гофрированные клеточные ядра, что свидетельствует о повышении тонуса (рис. 2, Г в сравнении с 2, В).
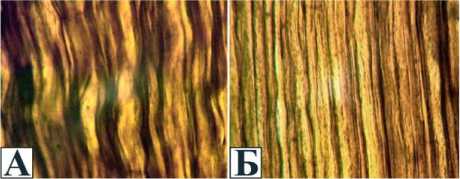
Рис. 3. Спрямление и истончение филаментознотубулярных тяжей при дистракционном остеосинтезе: А контралатеральный нерв; Б удлинённый нерв. Продольные криостатные срезы. Импрегнация серебром. УВ 500
Изменения нервных волокон при дистракционном остеосинтезе и поперечной тракции во многом сходны. В процессе дозированного растяжения сохраняется непрерывность их осевых цилиндров и шванновских оболочек (рис. 3 и 4) при некотором спрямлении и истончении филаментозно-тубулярных тяжей. В отдельных мякотных волокнах можно отметить признаки деформационного и гипоксического стресса: удлинение узлов Ранвье в результате паранодаль-ной демиелинизации и расширение насечек Шмидта-Лантермана (рис. 5).
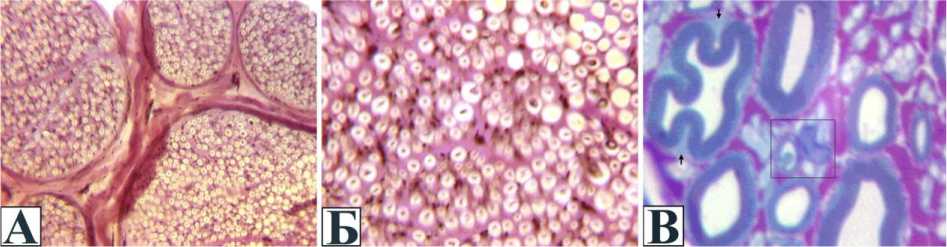
Рис. 4. Нервные волокна в конце периода дистракции, серия 1: А и Б - поперечный криостатный срез, окраска по Ван Гизо-ну; А УВ 200х; Б УВ 500 ; В - поперечный полутонкий срез (окраска по Уикли, УВ 1250 ). В рамке новообразую-щееся мякотное нервное волокно в ассоциации с незрелым аксо-глиальным комплексом
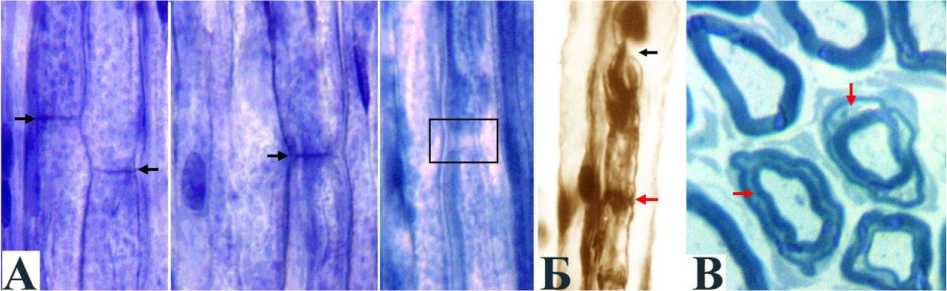
Рис. 5. Признаки деформационного стресса нервных волокон в конце дистракции: А продольный криостатный срез, окраска гематоксилином Вейгерта, УВ 200 ; Б импрегнация по Рассказовой, УВ 200 ; В полутонкий срез, окраска толуидиновым синим, УВ 1250 . Чёрные стрелки нормальные перехваты Ранвье, в рамке "удлинённый" перехват Ранвье. Красные стрелки расширенные насечки Шмидта Лантермана
Фигуры аксональной дегенерации и демиелинизации единичны. Наряду с этим регулярно встречаются незрелые аксо-глиальные комплексы и новообразованные мякотные волокна малого диаметра с тонкой миелиновой оболочкой в ассоциации со зрелыми безмякотными волокнами. В некоторых пучках они рассеяны по эн-доневрию хаотично, в других образуют скопления в субпериневральной зоне и вблизи эндо-невральных септ. Состояние аксоскелета разрежение или конденсация нейрофиламентов и микротрубочек в зрелых волокнах, их аккордное расположение в новообразующихся аксо-глиальных комплексах отражает разнообразные изменения скорости аксонного транспорта.
При нейротензии спектр деструктивных изменений более широк, чем при дистракционном остеосинтезе: встречались картины валлеров-ской, аксональной дегенерации, демиелинизации и томакулярные изменения, хотя частота фигур деструкции от 3 до 5 %. Регенерационные кластеры, признаки арборизации мякотных волокон, новообразованные миелинизирующиеся волокна в составе незрелых униаксональных и полиаксональных аксо-глиальных комплексов обнаруживались нечасто.
По данным морфометрического анализа, численная плотность мякотных волокон в первой серии опытов к концу дистракции и особенно в ранний период фиксации увеличивается по сравнению с контралатеральным и особенно интактным нервом. Распределение мякотных волокон по диаметру остаётся бимодальным (рис. 6). Зависимость аксо-миелинового отношения от диаметра нервных волокон (рис. 7) выявляет две основные тенденции перестройки мякотных проводников. Повышение этого параметра во фракциях малого калибра доказывает присутствие новообразованных волокон с тонкой миелиновой оболочкой. Снижение его во фракциях крупного калибра отражает уменьшение диаметра и площади аксонов, связанное с нарушениями аксоплазматического тока.
В опытах с нейротензией изменения морфометрических параметров нервных волокон имеют такую же направленность, но в количественном отношении они менее выражены.
Совокупность изменений количественных параметров в сочетании с феноменологическими находками свидетельствует о том, что в удлиняемом нерве появляется большое количество новообразованных волокон и происходит перераспределение аксоплазматического тока между зрелыми и новообразующимися волокнами. По-видимому, новообразованные волокна являются отростками тех же нейронов или их дополнительными ветвлениями.
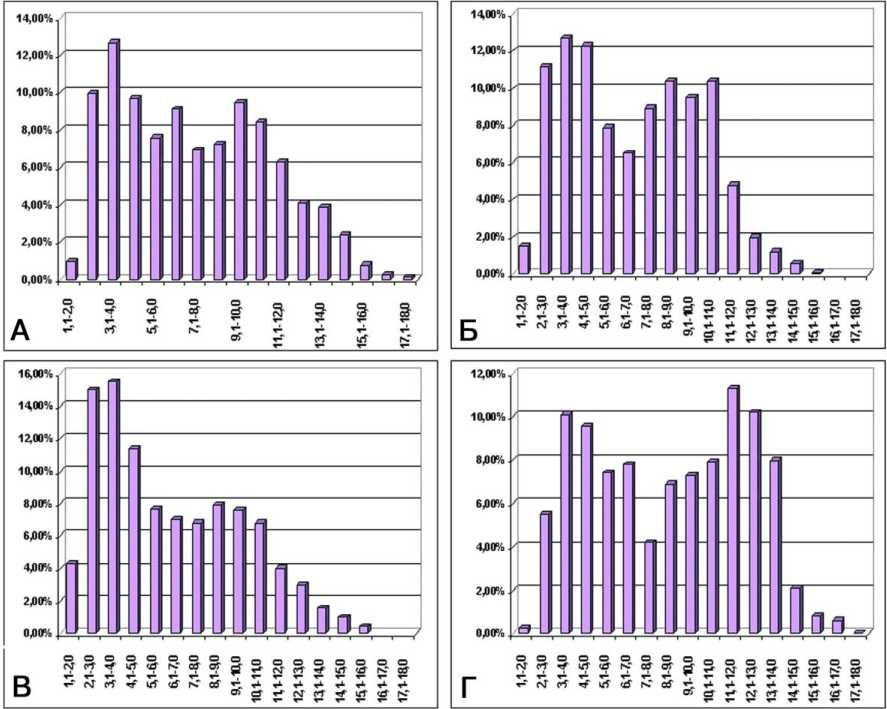
Рис. 6. Усреднённые гистограммы распределения мякотных волокон по спектру калибров в интактном нерве и опытах с дистракцией: A - по данным от интактных собак; Б - в конце дистракции; В - через 30 дней фиксации; Г - после снятия аппарата
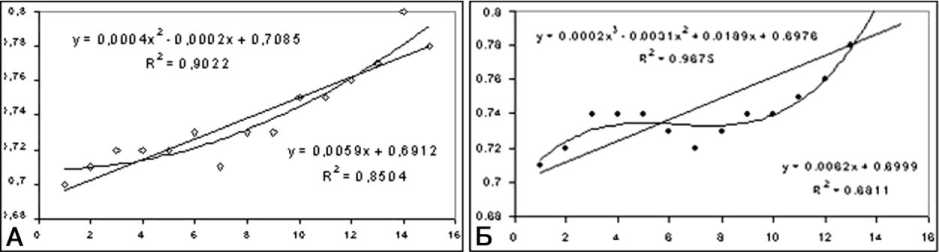
Рис. 7. графики зависимости числа G от диаметра мякотных нервных волокон: А - интактный нерв; Б - удлинённый нерв в конце периода дистракции. Ось абсцисс - диаметр мякотных волокон в микрометрах, ось ординат - значение числа G
В зрелых волокнах выражена базофилия и гипертрофия цитоплазмы шванновских клеток, ультраструктурно в ней определяется повышение содержания свободных рибосом, что свидетельствуют об усилении пластического метаболизма. Внутриклеточная гипертрофия шванновских клеток обеспечивает удлинение сегментов миелина мякотных волокон, о чём свидетельствует снижение доли ядросодержащих профилей мякотных волокон в поперечных срезах и снижение численности перехватов Ранвье в продольных срезах удлинённых нервов. В эндоневрии удлиняемого нерва можно встретить свободные (не связанные с аксонами) шванновские клетки с признаками подвижности цитолеммы, что малохарактерно для интактного нерва. По-видимому, они участвуют в формировании и элиминации вставочных сегментов миелина, репарирующих участки паранодальной демиелинизации, которые обнаруживаются в удлинённом нерве на этапе дистракции и в ранние сроки фиксации.
Признаки адаптационного роста зрелых волокон во второй серии опытов выражены ещё сильнее, чем в первой. Усиление базофилии цитоплазмы миелинобразующих шванновских клеток и их гипертрофия обнаруживалась в па-ранодальных и интернодальных срезах мякотных нервных волокон и особенно на уровне перикарионов (рис. 8). Даже на свето-оптическом уровне отчётливо выявлялись осевые цилиндры безмякотных волокон на фоне резко базофильной цитоплазмы шванновских клеток.
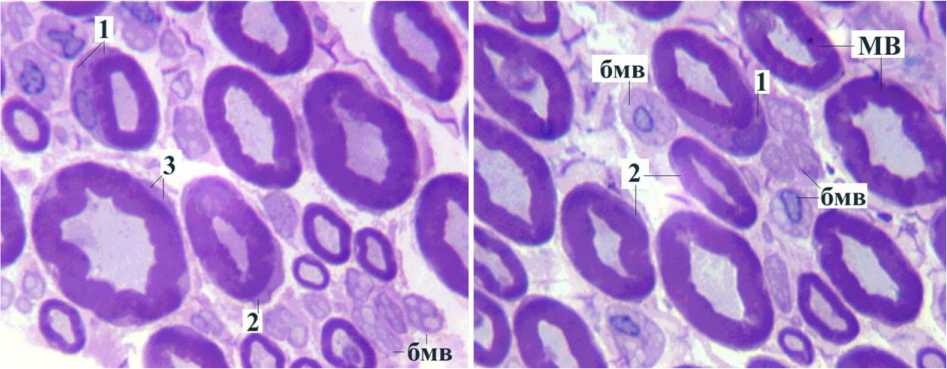
Рис. 8. Нервные волокна удлинённого седалищного нерва при тракции в поперечном направлении. Поперечный полутон-кий срез, окраска по Уикли. УВ 1250 . МВ мякотные волокна, бмв безмякотные волокна, 1 гипертрофия цитоплазмы миелинобразующих шванновских клеток в области перикариона, 2 на уровне интернода, 3 на уровне паранода
ЗАКЛЮЧЕНИЕ
По данным Г.А. Илизарова с соавт. (1982), "в оболочках нервов удлиняемой конечности взрослых животных происходит рекапитуляция целого ряда морфологических признаков, характерных для тканей, развивающихся в пре- и постнатальном периодах онтогенеза", в том числе свойственное растущим органам интенсивное и массовое новообразование собственных нервных элементов эпипериневрия. Результаты нашего исследования расширяют представления о морфогенетических процессах проводниковой части удлиняемого нерва в частности позволяют уточнить качественные и количественные критерии сходства структуры удлиняемого нерва с нервами растущих особей:
-
1. Повышенное отношение численности без-мякотных волокон к численности мякотных и общее повышение численности проводников по сравнению со среднестатистическими показателями нервов зрелых интактных особей.
-
2. Наличие большого количества недифференцированных аксо-глиальных комплексов и новообразованных волокон с тонкой миелиновой оболочкой.
-
3. Признаки повышенной биосинтетической и энергетической активности в цитоплазме шванновских клеток, образующих неврилемму зрелых нервных волокон.
-
4. Разнообразие изменений расположения элементов аксоскелета и органелл, указывающих на замедление и интенсификацию аксонального транспорта.
-
5. Признаки миграции шванновских клеток и ремоделирования сегментов миелина.
Кроме того, полученные результаты раскрывают морфологические эквиваленты обратимых нарушений функции удлиняемого нерва и обосновывают использование эффекта Илизарова для возмещения травматических дефектов нервных стволов.
