Альтернативный путь дыхания в растениях: регуляция и функции

Автор: Гармаш Елена Владимировна
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Биологические науки
Статья в выпуске: 3, 2010 года.
Бесплатный доступ
Представлены и обсуждены данные по функционированию электрон-транс- портной цепи (ЭТЦ) растительных митохондрий. Сформулированы ключевые вопросы изучения энергетически малоэффективного альтернативного (циа- нидрезистентного) пути дыхания (АП) в поддержании дыхательного метабо- лизма углерода, окислительно-восстановительного баланса в ЭТЦ, контроле накопления активных форм кислорода в норме и при стрессе. На основе соб- ственных экспериментальных данных показано, что дыхание и соотношение дыхательных путей в растениях варьирует в зависимости от объекта, его функционального состояния, силы и продолжительности действия факторов среды. Сделан вывод о том, что вовлечение АП расширяет адаптивные воз- можности растительного организма, способствует поддержанию роста и защи- те от стресса. Обсуждены механизмы регуляции дыхания и возможное уча- стие АП в процессах апоптоза и поддержании взаимосвязи фотосинтеза и ды- хания. Определены перспективы развития дальнейших исследований.
Растительные митохондрии, дыхательная цепь, альтернативное дыхание, альтернативная оксидаза, регуляция, рост, стресс
Короткий адрес: https://sciup.org/14992398
IDR: 14992398 | УДК: 581.12:576.311.347
The alternative respiration pathway in plants: regulaion and finctions
Information on functioning of plant mitochondria electron-transport chain is presented and discussed. Crucial problems of studying a wasteful alternative (cyanide-resistant) respiratory pathways (AP) for maintenance a respiratory carbon metabolism, redox balance during mitochondrial electron transport, and alleviating formation of reactive oxygen species under normal growth conditions and stress are formulated. Our experimental data show that respiration and respiratory pathways ratio depends on plant material, its functional state, intensity and period of environmental factors impact. It is concluded that AP engagement expands adaptive possibilities of plant, contributes to growth maintenance and protection against stress. Mechanisms of a possible AP participation in apoptosis and in regulation of photosynthesis and respiration relationship are discussed. The outlook for the further investigation of alternative respiration is shown.
Текст научной статьи Альтернативный путь дыхания в растениях: регуляция и функции
Дыхание – универсальный процесс и фундаментальная основа жизни любой живой клетки. Ежесуточно растения окисляют в дыхании от 30 до 70% ассимилированного при фотосинтезе углерода. Интермедиаты и энергия, извлекаемая в процессе дыхания, необходимы для синтеза de novo и поддержания целостности и функциональной активности клеточных структур, специализированных тканей и органов растительного организма. Дыхание – это показатель функциональной стойкости растений и равноправный фотосинтезу элемент про- дукционного процесса. Дыхание выступает как механизм, ускоряющий эволюцию. Известно, что окисление ДНК образуемыми в процессе дыхания активными формами кислорода представляет собой важнейший инструмент природного мутагенеза [1]. Поэтому проблема дыхания с научных позиций привлекает исследователей уже около двух столетий.
В лаборатории экологической физиологии растений Института биологии Коми НЦ УрО РАН исследования дыхания растений ведутся более 30 лет. Сформулированы и разработаны фундаментальные вопросы физиологии дыхания растений [2]. В настоящее время в рамках исследования механизмов регуляции дыхания растений уделяется особое внимание изучению дыхательных путей в растительной клетке в норме и при стрессе.
Дыхание – это поэтапный компартментиро-ванный процесс. Первые этапы дыхания протекают в цитоплазме, а основной продукт гликолиза – пируват – диффундирует в специализированные дыхательные органеллы – митохондрии, где подвергается окислительному декарбоксилированию в цикле трикарбоновых кислот (ЦТК). В реакциях цикла образуются соединения с высоким восстановительным потенциалом – пиридиннуклеотиды НАДН и ФАДН 2 . На заключительном этапе восстановительные эквиваленты окисляются с образованием энергии в ЭТЦ, локализованной во внутренней мембране митохондрий.
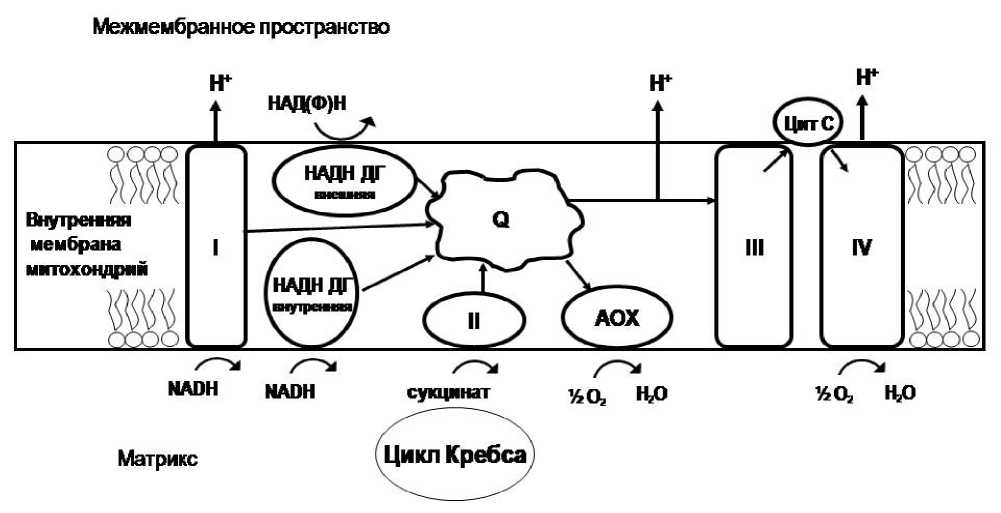
Основная ЭТЦ состоит из четырех трансмембранных белковых мультиферментных комплексов, двух небольших по молекулярной массе лабильных компонентов – убихинона и цитохрома с, выполняющих функцию переносчиков электронов между комплексами, и АТФ-синтазы. Перенос пары электронов по основному цитохромному пути (ЦП) от НАДН к кислороду с последующим синтезом трех молекул АТФ является универсальным для животных и растений.
Дыхательная цепь растений сложнее и пластичнее в функциональном плане, чем животных (рис. 1). Главной уникальной особенностью митохондрий высших растений, а также многих водорослей, мхов, папоротников и некоторых грибов является способность переноса электронов с пула убихинона на О 2 через альтернативную оксидазу [3]. Этот путь устойчив к действию цианида, поэтому назван цианидустойчивым (альтернативным).
Альтернативная оксидаза (АОХ) – убихинол-кислород-оксидоредуктаза – это низкомолекулярный белок (32 кДа), представляет собой один полипептид. АОХ выделена и идентифицирована более 20 лет назад [4]. Синтез белка кодируется ядерны- ми генами – Аох [5]. В мембране АОХ существует как мономер и димер. Активация фермента связана с восстановлением дисульфидной связи в сульфгидрильную [6].
Электронный транспорт по АП не связан с двумя участками генерации мембранного потенциала (комплексы III и IV). Лишь при окислении эндогенного НАДН градиент протонов может устанавливаться в комплексе I. Поэтому АП запасает мало энергии, большая часть рассеивается в виде тепла. Другими словами, альтернативный путь транспорта электронов снижает энергетическую эффективность дыхания, как минимум, на 60%. Тогда зачем растениям нужен этот путь?
Есть мнение, что альтернативное дыхание – это возникший в процессе эволюции растений способ поддержания дыхания в присутствии цианида. Многие растения (около 2500 видов) содержат цианогенные гликозиды, распад которых сопровождается образованием синильной кислоты и цианидов. Биологическая роль этих соединений может быть защитной, направленной против фитопатогенов или травоядных животных. Однако это не объясняет, почему АОХ есть у всех растений и какую роль она выполняет в клетке.
Хорошо известно, что АП участвует в термогенезе тканей цветка некоторых ароидных. Нагревание термогенных тканей облегчает испарение соединений, привлекающих насекомых-опылителей. Благодаря этому феномену, описанному еще Ламарком в XVIII в., в 30-х гг. XX в. было открыто циа-нидрезистентное дыхание (Okunuki, 1932, 1939; Van Herk, 1937, цит. по [7]). С тех пор минул почти век, но вопрос о механизмах регуляции и физиологической роли АП в нетермогенных тканях до сих пор остается открытым [3, 8, 9, 10].
Значительный вклад в развитие представлений об активации различных оксидаз и смены дыхательных путей в зависимости от интенсивности и
надндг
НАДН
АОХ
Цикл Кребса
Матрикс
NADH
NADH сукцинат
%О, Н,0
Межмембранное пространство
Внутренняя мембрана митохондрий
% О, НгО
н* А НАД(Ф)Н
Рис. 1. Цепь переноса электронов во внутренней мембране растительных митохондрий. Объяснения см. в тексте.
направленности процессов метаболизма, а также биологического состояния растения, внесла О.А. Семихатова. Она впервые обратила внимание на то, что в листьях разного возраста функционируют разные дыхательные системы [11]. Основную роль в дыхании молодых листьев играет цитохромокси-даза, а в зрелых – цианидустойчивая система, природа которой в то время была неизвестна [11].
Объяснение роли АП в нетермогенных тканях предложено в 1982 г. голландским ученым H. Lambers [12]. Согласно сформулированной им гипотезе «сверхпотока» («energy overflow») при насыщении цитохромного пути АП работает как «выхлопной клапан», через который «сжигается» избыток углеводов, который клетка не в состоянии эффективно использовать. Однако позже Lambers и его коллеги установили, что цитохромный и альтернативный пути дыхания конкурируют за электроны [13]. Поэтому гипотеза «сверхпотока» несколько трансформировалась. Считается, что АД участвует в поддержании баланса между углеводным метаболизмом и скоростью электронного транспорта, а именно в снижении высокого соотношения АТФ/АДФ за счет быстрого окисления НАДН без образования АТФ. При этом цикл Кребса продолжает функционировать. Такая ситуация может быть целесообразна для растений в период активного роста, когда они больше нуждаются в метаболитах для синтезов de novo , чем в АТФ. Нами показано, что скорость дыхания и подавление дыхания в присутствии специфического ингибитора АОХ были выше в корнях и листьях быстро растущих растений ячменя, чем у растений с низкой скоростью роста (рис. 2).
ем АОХ, в присутствии ингибиторов ЭТЦ [14]. Показано, что AOX участвует в стабилизации пула убихинонов (Q r /Q t ) in vivo [10]. Убедительным также является тот факт, что H 2 O 2 – вторичный мессенджер в сигнальном каскаде, приводящем к экспрессии гена Aox 1 [3].
Стимуляцию АП наблюдали при действии неблагоприятных факторов среды, которые могут вызвать окислительный стресс в клетке. Показано, что активность АП и количество белка АОХ возрастали при засухе, внедрении патогена [15], дефиците элементов минерального питания [16-18], низких положительных температурах [19-21]. В других работах не выявлено активации АОХ в ответ на действие факторов, в частности, низкой температуры [8, 21, 22]. Проанализировав литературу, мы предположили, что степень вовлечения АП и его физиологическая роль в ответ на изменение факторов зависят, в первую очередь, от силы и продолжительности действия фактора, устойчивости организма и его функционального состояния.
Нами проведены эксперименты с холодостойкими сортами ячменя ( Hordeum distichum L., с. Андрей, Новичок), длительно адаптированными к двум температурным режимам выращивания – 13/8ºС и 21/17ºС (день/ночь). Эти температуры ограничивают зону температурного оптимума роста данных сортов [23]. При пониженной температуре дыхание органов растений не проявляло чувствительности к ингибитору АОХ (рис. 3). Следовательно, можно полагать, что дыхание осуществлялось в основном по ЦП, без заметного участия АП. По-видимому, отсутствие чувствительности к ингибитору АОХ у холодостойкого сорта ячменя, выращи-
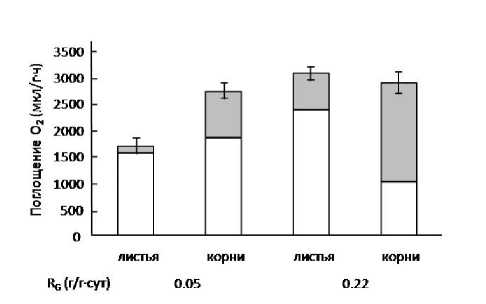
Рис. 2. Соотношение дыхательных путей в медленно- и быстрорастущих растениях ячменя ( Hordeum distichum L., с. Андрей). R G – относительная скорость роста, – SHAM-чувствительное (альтернативное) дыхание.
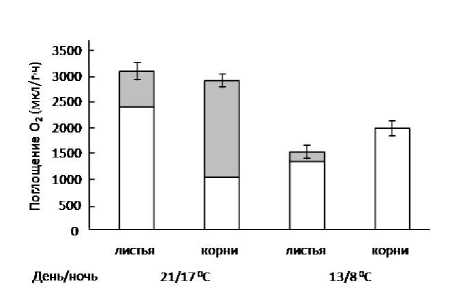
Рис. 3. Влияние температурного режима выращивания (день/ночь) на соотношение дыхательных путей в растениях ячменя ( Horedeum distichum L., с. Андрей). Обозначения как на рис. 2.
Другая функция АП связана с защитой клетки от окислительного стресса. Митохондрии являются местом образования активных форм кислорода (АФК), в частности супероксидрадикала и H2O2, чему способствуют торможение электронного транспорта и «перевосстановление» цепи. Активация АОХ приводит к снижению степени восстановлен-ности пула убихинонов за счет быстрого сброса электронов на кислород с образованием воды [14]. Прямым доказательством этой функции АП явились данные по накоплению АФК в тканях трансгенных растений табака с пониженным содержани- ваемого при более низкой (но в пределах температурного оптимума роста) температуре, обусловлено необходимостью сохранения максимальной энергетической эффективности дыхания для поддержания процессов жизнедеятельности. Это согласуется с современными представлениями о том, что регуляция энергетического метаболизма клетки направлена на избежание вовлечения АП [24].
В другом эксперименте изучали влияние кадмия на соотношение дыхательных путей в органах ячменя (Hordeum distichum L., сорт Новичок) при этих же температурных режимах [25]. Показано, что корни обработанных кадмием растений проявляли чувствительность к ингибитору АОХ – SHAM (рис. 4). Увеличение доли АП происходило на фоне повышения концентрации малонового диальдегида – продукта пероксидации липидов – свидетельствующем об окислительном стрессе. Известно, что АОХ препятствует генерации АФК в ЭТЦ митохондрий за счет быстрого сброса электронов на кислород и снижения степени восстановленно-сти пула убихинонов [3, 10, 19]. Значительное повышение доли АП (40% от общего дыхания) при низкой концентрации Cd (30 мкмоль/л) в корнях растений в условиях температурного режима 21/17º С, когда скорость общего дыхания еще достаточно высока, по-видимому, обусловлено необходимостью поддержания интенсивной работы ЦТК. Органические кислоты ЦТК способны связывать ТМ с последующей изоляцией в вакуоли [26]. Этот механизм мог активироваться и при сильном кадмиевом стрессе (100 мкмоль/л) в условиях более низкой температуры. Таким образом, активация АП является одним из механизмов поддержания гомеостаза подверженных стрессу кадмием клеток корней.
дыхание, снижая уровень АФК и препятствуя активации программируемой смерти клетки [27, 28]. Поэтому АОХ называют антиапоптическим белком (antiapoptotic protein) [27, 29].
В зеленых тканях растений фотосинтез и дыхание протекают в одной клетке. Однако долгое время эти процессы изучались автономно, и вопрос об их взаимодействии в фотосинтезирующей клетке остается до сих пор наименее изученным. Согласно современным данным на свету между хлоропластами и митохондриями идет активный обмен метаболитами. Дыхательные ферменты, в том числе альтернативные дегидрогеназы и АОХ в митохондриальной ЭТЦ, участвуют в окислении восстановительных эквивалентов, поступающих из цитозоля или хлоропластов в митохондрии при их близком контакте [30]. Это один из способов защиты клетки от фотоокисления.
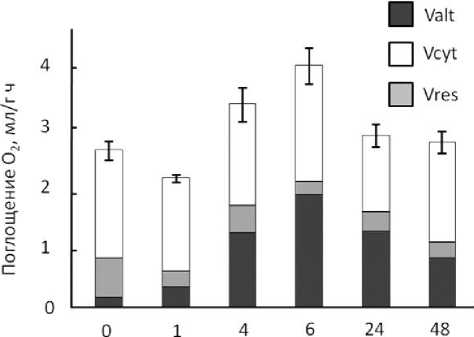
Нами исследовано вовлечение АП в дыхание листа пшеницы в процессе его зеленения [31]. Зеленение – процесс образования хлорофилла. В этиолированных проростках дыхание осуществлялось, главным образом, по цитохромному пути (рис. 5).
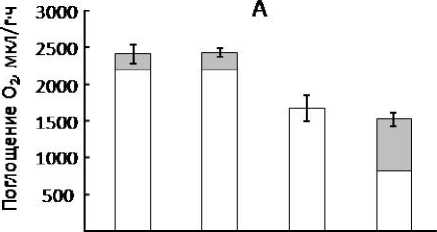
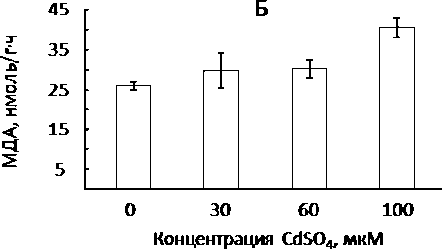
Рис. 4. Влияние кадмия на соотношение дыхательных путей (А) и активность ПОЛ (Б) в корнях ячменя ( Hordeum distichum L., с. Новичок) при пониженной температуре выращивания (день/ночь – 13/8 °С). Обозначения как на рис. 2.
Необходимо также назвать еще одну возможную причину усиления дыхания по АП в варианте c высокой концентрацией кадмия и пониженной температурой. На фоне остановки роста и частичной гибели растений вовлечение АОХ могло быть вызвано процессом вынужденной гибели клеток. Согласно современным данным митохондрии – главные участники апоптоза, сигнальными молекулами которого являются АФК [27]. Повышение АФК в клетке ведет к формированию специальных пор в мембране митохондрий и выходу цит с. Возможно, что на начальном этапе, когда цитохромный путь ограничен, АОХ поддерживает митохондриальное
Время на свету, ч
Рис. 5. Дыхание листьев пшеницы ( Triticum aesti-vum L., с. Иргина) в процессе зеленения.
– V alt , – Vcyt, – Vres – активность альтернативного, цитохромного и остаточного дыхания, соответственно.
Свет индуцировал вовлечение АП, доля которого в течение суток после начала освещения возрастала до 50% от общего дыхания. В этот же период зеленение сопровождалось увеличением пространственной зависимости расположения митохондрий в зоне нахождения хлоропластов, что указывает на наличие обмена метаболитами между органеллами [31]. Эксперимент показывает, что митохондрии играют важную роль в формировании фотосинтетического аппарата, а АП может участвовать в поддержании взаимосвязи фотосинтеза и дыхания.
Таким образом, энергетически малоэффективный АП является специфическим механизмом, поддерживающим клеточный гомеостаз и функционирование растений в различных условиях среды. Вовлечение АП расширяет адаптивные возможности растительного организма, способствует поддержанию роста и защите от стресса. Убедительными являются данные об участии АОХ в термогенезе специализированных тканей цветка ароидных, поддержании окислительно-восстановительного баланса в ЭТЦ митохондрий и препятствии формирования АФК, особенно в условиях стресса. В настоящее время наиболее сильно развиваются исследования о связи альтернативного дыхания с ростом, процессом апоптоза, генерацией тепла в «нетермогенных» тканях растений при гипотермии, защитой от фотоокисления и др. Спорными остаются вопросы регуляции активности и вовлечения АОХ in vivo. Мы обратили внимание на то, что дыхание и соотношение дыхательных путей в растениях варьируют в зависимости от объекта, его функционального состояния, продолжительности и силы действия внешних факторов, устойчивости к ним растительного организма. Идея многофункциональности АП нашла подтверждение в наших экспериментах.
Необходимо отметить, что исследование альтернативного пути дыхания вышло из рамок только фундаментальной проблемы. Показатели дыхания и соотношения дыхательных путей могут быть использованы для выявления реакции растений в ответ на изменение условий среды (глобальное потепление, увеличение СО 2 в атмосфере). Показано усиление активности АП в листьях Arabi-dopsis thaliana , селектированных для роста в условиях повышенной концентрации СО 2 [32]. В экспериментах, направленных на борьбу с митохондриальными болезнями человека, показано, что ген АОХ может быть успешно введен в человеческие клетки почки и фибробласты. Мутантные клетки проявляли толерантность к дефициту цитохромок-сидазы [33].
В заключение следует сказать, что вопросы регуляции активности и вовлечения альтернативного дыхания находятся в настоящее время в центре особенного внимания экспериментальных биологов, особенно за рубежом (Physiologia plantrum, Vol. 129, 2007; Vol. 137, 2009). Интенсивное получение сведений о функционировании этого уникального пути в растениях, с одной стороны, приближает нас к разгадке вопроса о причинах активации альтернативной оксидазы in vitro , in vivo , in situ , с другой – ставит перед исследователями много новых задач. Зная законы регуляции соотношения дыхательных путей, можно контролировать (на молекулярногенетическом уровне) жизнедеятельность не только, как оказалось, растений, но и человека. Надеемся, что мы внесем свою лепту в поток этой важной информации.
Автор признательна своему научному руководителю профессору Т.К. Головко, вдохновившей на изучение этой проблемы, а также за ценные советы при обсуждении работы.
Список литературы Альтернативный путь дыхания в растениях: регуляция и функции
- Скулачев В.П. Эволюция, митохондрии и кислород//Соросовский образовательный журнал, 1999. № 9. С. 1-7.
- Головко Т.К. Дыхание растений (физиологические аспекты). С-Пб.: Наука, 1999. 204 с.
- Vanlerberghe G.C., McIntosh L. Alternative oxidase: from gene to function//Annu. Rev. Plant Physiol. Plant Mol. Biol., 1997. Vol. 48. P. 703-734.
- Elthon T.E., McIntosh L. Characterization and solubilization of the alternative oxidase of Sauromantum guttatum mitochondria//Plant Physiol., 1986. Vol. 82. P. 1-6.
- Whelan J., Millar A.H., Day D.A. The alternative oxidase is encoded in a multigene family in soybean//Planta, 1996. Vol. 198. P. 197-201.
- Umbach A.L., Siedow J.N. Covalent and noncovalent dimers of the cyanide-resistant alternative oxidase protein in higher plant mitochondria and their relationship to enzime activity//Plant Physiol., 1993. Vol. 103. P. 845-854.
- Meeuse B.J.D. Thermogenic respiration in aroids//Annu. Rev. Plant Physiol., 1975. Vol. 26. P. 117-126.
- Головко Т.К., Пыстина Н.В. Альтернативный путь дыхания в листьях Rhodiola rosea L. и Ajuga reptans L.: возможная физиологическая роль//Физиология растений, 2001. Т.48. С. 846-853.
- Шугаев А.Г. Альтернативная CN-резистентная оксидаза митохондрий растений: структурная организация, механизмы регуляции активности, возможная физиологическая роль//Физиология растений, 1999. Т. 46. С. 307-320.
- Millenaar F.F., Lambers H. The аlternative oxidase: in vivo regulation and function//Plant Biol., 2003. Vol. 5. P. 2-15.
- Семихатова О.А. Смена дыхательных систем. Л.: Изд-во «Наука», 1969. 128 с.
- Lambers H. Cyanide-resistant respiration: a non-phosphorylating electron transport pathway acting as an energy overflow//Physiol. Plant., 1982. Vol. 55. P. 478-485.
- The cyanide-resistant oxidase: to inhibit or not inhibit, that is the question/D.A.Day, K.Krab, H.Lambers, A.L.Moore, J.N.Siedow, A.M.Wagner, J.T.Wiskich//Plant Physiol., 1996. Vol. 110. P.1-2.
- Maxwell D.P., Wang Y., McIntosh L. The alternative oxidase lowers mitochondrial reactive oxygen production in plant cells//Proc. Nat. Acad. Sci USA., 1999. Vol. 96. P. 8271-8276.
- Maxwell D.P., Nickels R., McIntosh L. Evidence of mitochondrial involvement in the transduction of signals required for the induction of genes associated with pathogen attack and senescence//Plant J., 2002. Vol. 29. P. 269-279.
- Рахманкулова З.Ф., Федяев В.В., Подашевка О.А., Усманов И.Ю. Альтернативные пути дыхания и вторичный метаболизм у растений с разными типами адаптивных «стратегий» при дефиците элементов минерального питания//Физиология растений, 2003. Т. 50. С. 231-237.
- Energy metabolism of Plantago lanceolata as dependent on the supply of mineral nutrients/H.Lambers, F.Posthumus, I.Stulen, L.Lanting, S.van de Dijk, R.Hofstra//Physiol. Plant., 1981. Vol. 51. P. 85-92.
- The role of alternative oxidase in modulating carbon use efficiency and growth during macronutrient stress in tobacco cells/S.M.Sieger, B.K.Kristensen, C.A.Robson, S.Amirsadeghi, E.W.Eng, A.Abdel-Mesih, I.M.Moller, G.C.Vanlerberghe//J. Exp. Bot., 2005. Vol. 56. P. 1499-1515.
- Fiorani F., Umbach A.L., Siedow J.N. The Alternative oxidase of plant mitochondria is involved in the acclimation of shoot growth at low temperature. A study of Arabidopsis AOX1a transgenic plants//Plant Physiol., 2005. Vol. 139. P. 179-180.
- The distribution of electron tran-sport between the main cytochrome and alternative pathways in plant mitochondria during short-term cold stress and cold hardening/O.I.Grabelnych, O.N.Sumina, S.P.Funderat, T.P.Pobezhimova, V.K.Voinikov, A.V. Kolesnichenko//J. Thermal Biol., 2004. Vol. 29. P. 165-175.
- The electron partitioning between the cytochrome and altenative respiratory pathways during chilling recovery in two cultivars of maize differing in chilling sensitivity/M.Ribas-Carbo, R.Aroca, M.A.Gonzаlez-Meler, J.J.Irigoyen, M.Sanchez-Diaz//Plant Physiol., 2000. Vol. 122. P. 199-204.
- Иванова Т.И., Кирпичникова О.В., Шерстнева О.А., Юдина О.С. Годичный цикл дыхания листьев вечнозеленых растений//Физиология растений, 1998. Т.45. С. 906-913.
- Головко Т.К., Родина Н.А., Куренкова С.В.,Табаленкова Г.Н. Ячмень на Севере (селекционно-генетические и физиолого-биохимические основы продуктивности). Екатеринбург: УрО РАН, 2004. 186 с.
- Priault P., Vidal G., De Paepe R., Ribas-Carbo M. Leaf age-related changes in respiratory pathways are dependent on complex I activity in Nicotiana sylvestris//Physiol. Plant., 2007. Vol. 129. P. 152-162.
- Гармаш Е.В., Головко Т.К. Влияние кадмия на рост и дыхание ячменя при двух температурных режимах выращивания//Физиология растений, 2009. Т. 56. С. 382-387.
- Lösch R., Köhl K.I. Plant respiration under the influence of heavy metals//Heavy metal stress in plants. From molecules to ecosystem/Eds Prasad M.N.V., Hagemeyer J. Berlin: Springer-Verlag, 1999. P. 139-156.
- Plant mitochondrial pathway leading to programmed cell death/A.Vianello, M.Zancani, C.Peresson, E.Petrussa, V.Casolo, J.Krajnakova, S.Patui, E.Braidot, F.Macri//Physiol. Plant., 2007. Vol. 129. P. 242-252.
- Vanlerberghe G.C., Robson C.A., Yip J.Y.H. Induction of mitochondrial alternative oxidase in response to a cell signal pathway downregulating the cytochrome pathway prevents programmed cell death//Plant Physiol., 2002. Vol. 129. P. 1829-1842.
- Marechal E., Baldan B. Dual role of plant mitochondria in promoting PCD or cell survival//Trends Plant Sci., 2002. Vol. 7. P. 525-526.
- Noguchi K., Yoshida K. Interaction between photosynthesis and respiration in illuminated leaves//Mitochondrion, 2008. Vol.8. P. 87-99.
- Atkin O.K., Macherel D. The crucial role of plant mitochondria in orchestrating drought tolerance//Annals of Botany, 2009. Vol.103. P. 581-597.
- Engagement of alternative respiration in spring wheat seedlings during greening/E.V.Garmash, J.Kozlova, S.Plyusnina, R.Malyshev, Y.Yatsco, T.Golovko//Acta Biologica Cracoviensia, 2009. Vol.51. Suppl. 2. Abstracts of the 4th Conference of Polish Society of Experimental Plant Biology (Cracow, Poland, 21-25 September 2009). P. 43.
- Plastic and adaptive responses of plant respiration to changes in atmospheric CO2 concentration/M.A.Gonzalez-Meler, E.Blanc-Betes, C.E.Flower, J.K.Ward, N.Gomez-Casanovas//Physiol. Plant., 2009. Vol. 137. P. 473-484.
- The alternative oxidase, a tool for compensating cytochrome c oxidase deficiency in human cells/E.P.Dassa, E.Dufour, S.Goncalves, H.T.Jacobs, P.Rustin//Physiol. Plant., 2009. Vol. 137. P. 427-434.
