Аминокислотный состав мышечной ткани сига обыкновенного Coregonus lavaretus L. Кольского полуострова

Автор: Мишанина Людмила Александровна
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Рубрика: Ихтиология
Статья в выпуске: 2 т.21, 2018 года.
Бесплатный доступ
Представлены результаты исследования аминокислотного состава белков и свободных аминокислот мышечной ткани разновозрастного сига обыкновенного Coregonus lavaretus L., обитающего в озерах Ловозеро и Умбозеро Кольского полуострова. Изучение аминокислотных спектров в отдельных тканях позволяет в определенной степени оценивать особенности белкового обмена и физиологическое состояние организма. Аминокислотный состав белков исследован методом высокоэффективной жидкостной хроматографии. Фракционный состав белков по растворимости определен спектрофотометрическим методом. Выявлены существенные различия в количественном соотношении связанных и свободных аминокислот в мышечной ткани сига обыкновенного. Установлено, что белки мышечной ткани сига оз. Ловозеро отличаются более высоким суммарным содержанием незаменимых аминокислот по сравнению с сигами оз. Умбозеро. Значительно повышается содержание незаменимой аминокислоты лейцина в белках мышечной ткани сига оз. Ловозеро, что объясняется увеличением содержания солерастворимой миофибриллярной фракции. Содержание незаменимых аминокислот валина и изолейцина существенно ниже у сига оз. Умбозеро в отличие от сига оз. Ловозеро. Возможно, недостаток этих незаменимых аминокислот приведет к нарушению метаболизма у исследуемых рыб оз. Умбозеро. Отношение основных аминокислот к кислым в мышечной ткани сига оз. Умбозеро меньше, чем у сига оз. Ловозеро. При половом созревании сига происходит уменьшение общего пула незаменимых свободных аминокислот, находящегося в обратной зависимости от содержания белковых аминокислот.
Сиг обыкновенный, свободные аминокислоты, связанные аминокислоты, незаменимые для рыб аминокислоты, озеро умбозеро, озеро ловозеро
Короткий адрес: https://sciup.org/142215141
IDR: 142215141 | УДК: 639.312:[591.133.1:577.122.3](470.21) | DOI: 10.21443/1560-9278-2018-21-2-295-302
Aminoacids composition of muscular tissue of Coregonus lavaretus L. (the Kola peninsula)
The studies of the protein aminoacid composition and free aminoacids of muscle tissue of different ages of Coregonus lavaretus L. from Lakes Umbozero and Lovozero (the Kola Peninsula) have been presented. The research of aminoacid spectra in separate tissues allows to some extent to evaluate the features of protein metabolism and the physiological state of the organism. Aminoacid composition of proteins has been investigated by high-performance liquid chromatography. Fractional composition of the proteins' solubility has been determined by spectrophotometry. Significant differences in the ratio of bound and free aminoacids of muscle tissue of Coregonus lavaretus L. have been substantiated. It is established that the proteins of the muscle tissue of whitefish from Lake Lovozero have a higher total content of essential aminoacids unlike whitefish from Lake Umbozero. The content of essential aminoacid leucine in the proteins of muscle tissue of whitefish from Lovozero increases significantly, which is associated with an increase in the content of salt-soluble myofibrillar fraction. The content of essential aminoacids valine and isoleucine is significantly lower in Coregonus lavaretus L. of Lake Umbozero compared to Coregonus lavaretus L. of Lake Lovozero. Perhaps the lack of these essential aminoacids will lead to the violation of metabolism in the studied fish from Lake Umbozero. The ratio of alkaline aminoacids to acidic ones in the muscle tissue of whitefish from Lake Umbozero is less than that of whitefish from Lake Lovozero. During whitefish puberty the overall pool of essential free aminoacids decreases, the pool is in inverse proportion to content of protein aminoacids.
Текст научной статьи Аминокислотный состав мышечной ткани сига обыкновенного Coregonus lavaretus L. Кольского полуострова
В настоящее время актуальным является исследование физиолого-биохимических механизмов адаптации рыб к экологическим условиям среды обитания, изучение функциональной роли различных классов органических веществ в адаптивных механизмах [1; 2]. Научный интерес представляет исследование ценного промыслового вида сига обыкновенного Coregonus lavaretus L., численность которого повсеместно сокращается.
Цель работы – изучение биохимического состава сига обыкновенного Coregonus lavaretus L. из озер Умбозеро и Ловозеро Кольского полуострова, а именно: исследование аминокислотного состава белков и свободных аминокислот мышечной ткани данных объектов для оценки особенностей метаболизма и развития рыб. Современные исследования аминокислотного состава рыб рода Coregonus проводились для изучения пищевой ценности [3–5].
Материалы и методы
Объектом исследования служил сиг обыкновенный Coregonus lavaretus L. в возрасте 3+ (перед половым созреванием) и 5+ (половозрелый) из озер Ловозеро и Умбозеро Кольского полуострова.
В качестве материала для исследования использовали мышечную ткань разновозрастного сига обыкновенного. Вылов сига происходил в летнее время (конец июня – начало июля). Пробы для анализа были объединенными и состояли из образцов мышечной ткани, взятых у 2–4 экземпляров одинакового возраста.
Для определения аминокислотного состава отбирали аликвоту супернатанта из депротеинизированных хлорной кислотой образцов мышечной ткани (свободные аминокислоты) и навеску тканевой массы (нерастворимая часть) из тех же образцов. Последнюю подвергали избыточному гидролизу (6 н HCl, 22 часа при 110 °С в ампуле под азотом) с последующим удалением соляной кислоты на вакуумном ротационном испарителе.
Аминокислотный состав белков исследовали методом высокоэффективной жидкостной хроматографии (ВЭЖХ). Хроматографирование проводили на базе жидкостного хроматографа Shimadzu Prominence LC-20 (Япония), оснащенного флуориметрическим детектором, с предколоночной дериватизацией ортофталевым альдегидом в присутствии 2-меркаптоэтанола. Колонка Zorbax C18 – 4,6 × 150 мм (Agilent/США); подвижная фаза – 0,10 М натрий-фосфатный буфер, рН 7,5, градиент метанола от 0 до 80 %, 1 мл/мин; детектор – флуориметр, длина волны возбуждения 229 нм, эмиссии – 470 нм [6]. Калибровку колонки проводили по аминокислотному стандарту фирмы SIGMA (США) методом абсолютной калибровки.
Фракционный состав белков в образцах ацетоновых концентратов мышечной ткани определяли по растворимости в ряду вода – раствор Вебера – 0,01%-й едкий натр спектрофотометрическим методом при длине волны 550 нм после биуретовой реакции [7].
Статистическую обработку результатов измерений проводили общепринятыми методами при уровне значимости p ≤ 0,05. Гипотезу о принадлежности сравниваемых независимых выборок к одной генеральной совокупности проверяли с помощью однофакторного дисперсионного анализа.
Результаты и обсуждение
Результаты исследований мышечной ткани сига обыкновенного на содержание связанных аминокислот представлены в табл. 1.
Таблица 1. Аминокислотный состав белков мышечной ткани сига обыкновенного Coregonus lavaretus L., % к общей сумме аминокислот
Table 1. Aminoacids composition of proteins of the muscle tissue of Coregonus lavaretus L., % of total aminoacids
|
Аминокислота |
Содержание связанных аминокислот |
|||
|
3+ Ловозеро |
5+ Ловозеро |
3+ Умбозеро |
5+ Умбозеро |
|
|
Аспарагиновая кислота |
16,3 ± 0,4 |
9,1 ± 0,3 |
15,8 ± 0,4 |
9,4 ± 0,3 |
|
Глутаминовая кислота |
17,1 ± 0,4 |
13,1 ± 0,3 |
15,3 ± 0,4 |
12,8 ± 0,3 |
|
Серин |
5,2 ± 0,2 |
4,3 ± 0,2 |
8,5 ± 0,2 |
9,8 ± 0,3 |
|
Глицин |
4,0 ± 0,1 |
5,4 ± 0,2 |
6,5 ± 0,2 |
7,7 ± 0,2 |
|
Гистидин |
3,0 ± 0,1 |
2,7 ± 0,1 |
3,6 ± 0,1 |
4,3 ± 0,1 |
|
Треонин |
4,3 ± 0,1 |
4,0 ± 0,1 |
5,0 ± 0,2 |
5,6 ± 0,2 |
|
Аргинин |
6,2 ± 0,2 |
5,2 ± 0,2 |
4,9 ± 0,2 |
5,7 ± 0,2 |
|
Аланин |
3,7 ± 0,2 |
5,6 ± 0,2 |
7,8 ± 0,2 |
9,5 ± 0,2 |
|
Таурин |
0,2 ± 0,1 |
0,6 ± 0,1 |
0,3 ± 0,1 |
0,5 ± 0,1 |
|
Тирозин |
3,8 ± 0,1 |
3,2 ± 0,1 |
4,8 ± 0,1 |
5,8 ± 0,1 |
|
Валин |
5,4 ± 0,2 |
5,2 ± 0,1 |
3,1 ± 0,1 |
3,3 ± 0,1 |
|
Метионин |
3,1 ± 0,1 |
3,3 ± 0,1 |
2,7 ± 0,1 |
4,0 ± 0,1 |
|
Изолейцин |
3,9 ± 0,1 |
4,2 ± 0,1 |
1,5 ± 0,1 |
2,6 ± 0,1 |
|
Фенилаланин |
3,5 ± 0,1 |
4,7 ± 0,1 |
3,2 ± 0,1 |
4,5 ± 0,1 |
|
Лейцин |
5,0 ± 0,2 |
15,4 ± 0,3 |
7,4 ± 0,2 |
6,3 ± 0,2 |
|
Лизин |
15,3 ± 0,4 |
14,0 ± 0,3 |
9,6 ± 0,3 |
8,2 ± 0,2 |
В исследованных пробах изучаемых объектов в количественном отношении доминируют аспарагиновая и глутаминовая кислоты, серин, аланин, лейцин и лизин.
Содержание в белках фенилаланина, аспарагиновой и глутаминовой кислот у одновозрастного сига из разных водоемов не отличается.
При переходе сига в половозрелое состояние в белках мышечной ткани значительно уменьшается содержание заменимых для рыб аминокислот – аспарагиновой и глутаминовой кислот.
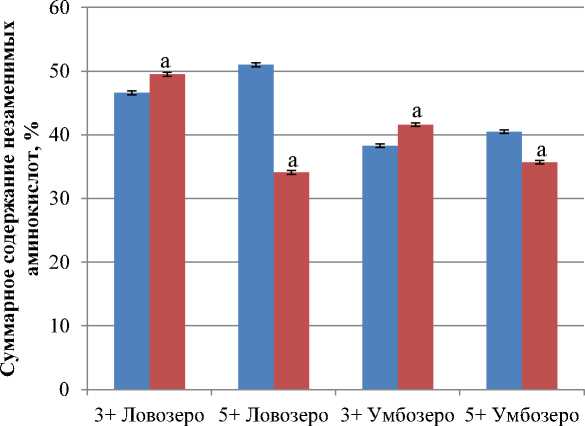
Белки мышечной ткани сига из исследуемых водоемов отличаются по общему содержанию незаменимых для рыб аминокислот. Их сумма у четырехлеток и шестилеток из оз. Ловозеро составляет 46,6 и 51 % от общей суммы связанных аминокислот соответственно, общее содержание незаменимых аминокислот у рыб из оз. Умбозеро более низкое – 38,3 и 40,5 % соответственно (рис. 1).
-
■ незаменимые связанные АК
-
■ незаменимые свободные АК
Рис. 1. Суммарное содержание незаменимых аминокислот в мышечной ткани сига обыкновенного Coregonus lavaretus L., % к общей сумме аминокислот; а – достоверность различий по сравнению со связанными аминокислотами, p ≤ 0,05
Fig. 1. The total content of essential aminoacids in the muscle tissue of Coregonus lavaretus L., % of total aminoacids; a – reliability of differences compared to related aminoacids, p ≤ 0,05
У сига оз. Ловозеро с возрастом происходит незначительное уменьшение аргинина, лизина и валина, а у сига оз. Умбозеро – лейцина. Значительно повышается содержание незаменимой аминокислоты лейцина в мышечной ткани сига оз. Ловозеро (до 15,4 % от общего содержания связанных аминокислот). Именно лейцин обладает особо выраженной способностью стимулировать биосинтез белка, клеточный метаболизм и рост клетки [8]. После проведения фракционирования белков по растворимости выявлено, что для половозрелого сига (5+) оз. Ловозеро характерно высокое содержание солерастворимой фракции в отличие от сига этого же возраста оз. Умбозеро (табл. 2). Повышение содержания лейцина в белке мышечной ткани сига оз. Ловозеро можно объяснить увеличением концентрации солерастворимой миофибриллярной фракции, основным белком которой является миозин, в большом количестве содержащий данную аминокислоту.
Таблица 2. Содержание белковых фракций в мышечной ткани сига обыкновенного Coregonus lavaretus L., % к общей сумме белков
Table 2. The content of protein fractions in the muscle tissue of Coregonus lavaretus L., % of total protein
|
Мышечная ткань |
Белковые фракции, % |
||
|
Водорастворимая |
Солерастворимая |
Щелочерастворимая |
|
|
3+ Ловозеро |
32,4 ± 3,6 |
55,3 ± 3,2 |
12,3 ± 1,3 |
|
5+ Ловозеро |
26,0 ± 2,8 |
64,2 ± 3,8 |
9,8 ± 1,4 |
|
3+ Умбозеро |
33,4 ± 3,3 |
53,7 ± 5,2 |
12,9 ± 1,2 |
|
5+ Умбозеро |
38,2 ± 3,7 |
46,9 ± 4,3 |
14,9 ± 1,3 |
Содержание таких незаменимых для рыб аминокислот, как валин и изолейцин, существенно ниже у сига оз. Умбозеро по сравнению с сигом оз. Ловозеро.
Снижение содержания изолейцина приводит к замедлению процессов роста и развития. При недостаточном поступлении валина отмечаются расстройства координации движений. Недостаточное содержание валина и изолейцина способствует изменению осморегуляции у рыб [8]. Можно предположить, что недостаток этих незаменимых аминокислот приведет к нарушению метаболизма у исследуемых рыб оз. Умбозеро.
Потребность рыб в любой из незаменимых аминокислот зависит от уровня других аминокислот в рационе. Недостаток аминокислот приводит к увеличению затрат корма на единицу прироста. При недостатке незаменимых аминокислот вступает в силу "закон минимума", согласно которому дефицит лишь одной незаменимой аминокислоты ограничивает эффективность использования всего рациона, что приводит к снижению роста, повышенному жирообразованию, и в конечном итоге увеличиваются затраты белка на единицу прироста рыб.
Результаты исследований мышечной ткани сига обыкновенного на содержание свободных аминокислот представлены в табл. 3.
Таблица 3. Свободные аминокислоты мышечной ткани сига обыкновенного Coregonus lavaretus L., % к общей сумме аминокислот
Table 3. Free aminoacids of the muscle tissue of Coregonus lavaretus L., % of total aminoacids
|
Аминокислота |
Содержание свободных аминокислот |
|||
|
3+ Ловозеро |
5+ Ловозеро |
3+ Умбозеро |
5+ Умбозеро |
|
|
Аспарагиновая кислота |
7,1 ± 0,2 |
7,8 ± 0,2 |
9,3 ± 0,3 |
7,8 ± 0,2 |
|
Глутаминовая кислота |
8,1 ± 0,2 |
8,6 ± 0,2 |
7,5 ± 0,2 |
6,9 ± 0,2 |
|
Серин |
3,3 ± 0,1 |
8,1 ± 0,2 |
5,4 ± 0,1 |
7,4 ± 0,2 |
|
Глицин |
7,9 ± 0,2 |
12,1 ± 0,3 |
8,1 ± 0,2 |
11,3 ± 0,3 |
|
Гистидин |
10,3 ± 0,3 |
11,9 ± 0,3 |
6,5 ± 0,2 |
8,2 ± 0,2 |
|
Треонин |
6,5 ± 0,2 |
7,3 ± 0,2 |
6,1 ± 0,2 |
8,4 ± 0,2 |
|
Аргинин |
2,9 ± 0,1 |
2,0 ± 0,1 |
4,2 ± 0,1 |
3,1 ± 0,1 |
|
Аланин |
11,3 ± 0,3 |
14,6 ± 0,3 |
9,0 ± 0,3 |
12,7 ± 0,3 |
|
Таурин |
0,2 ± 0,1 |
0,3 ± 0,1 |
0,3 ± 0,1 |
0,5 ± 0,1 |
|
Тирозин |
8,9 ± 0,2 |
12,1 ± 0,3 |
12,6 ± 0,3 |
14,6 ± 0,3 |
|
Валин |
2,1 ± 0,1 |
0,9 ± 0,1 |
3,2 ± 0,1 |
1,4 ± 0,1 |
|
Метионин |
3,7 ± 0,1 |
2,3 ± 0,1 |
6,2 ± 0,2 |
3,1 ± 0,1 |
|
Изолейцин |
2,5 ± 0,1 |
0,9 ± 0,1 |
4,2 ± 0,1 |
2,5 ± 0,1 |
|
Фенилаланин |
2,9 ± 0,1 |
2,5 ± 0,1 |
3,8 ± 0,1 |
3,5 ± 0,1 |
|
Лейцин |
3,3 ± 0,2 |
1,6 ± 0,1 |
3,2 ± 0,2 |
2,1 ± 0,1 |
|
Лизин |
19,0 ± 0,4 |
7,0 ± 0,2 |
10,4 ± 0,4 |
6,5 ± 0,2 |
У сигов из исследуемых водоемов имеются отличия в содержании свободных аминокислот, что может быть связано с разной кормовой базой изучаемых объектов.
Умбозеро является глубоким и слабо прогреваемым озером, в нем встречаются только малотычинковые сиги (с числом жаберных тычинок от 16 до 30). В более теплом неглубоком водоеме (озеро Ловозеро) обитают как малотычинковые, так и многотычинковые сиги (с числом тычинок от 30 до 60). В связи с этим у сигов имеются различия в питании. Сиги с большим количеством жаберных тычинок обладают повышенной способностью к фильтрации планктона жаберным аппаратом и следовательно, в питании стараются избегать крупных форм бентоса и мелкой рыбы. Малотычинковые сиги предпочитают питаться бентосом [9–12].
Глубоководность, преобладание каменистых берегов, бедность растительности, низкая температура воды, минерализация и содержание биогенных элементов в оз. Умбозеро обусловили невысокие количественные показатели планктонных и бентосных сообществ. Данный водоем характеризуется как ультраолиготрофный. Зоопланктон отнесен к бедному в кормовом отношении коловраточно-копеподитному типу и представлен веслоногими ракообразными, ветвистоусыми рачками, коловратками и двухстворчатыми моллюсками. Средняя биомасса зоопланктона 0,3–0,5 мг/л. Бентос представлен 33 таксонами. Средняя биомасса бентофауны – 0,85 г/м 2 , не менее 50 % приходится на хирономид [13].
В отличие от Умбозера Ловозеро – это олиготрофный водоем. В зоопланктоне оз. Ловозеро доминируют коловратки и ветвистоусые рачки; его средняя биомасса составляет 0,8 мг/л. Для Ловозера характерно сравнительно большое разнообразие беспозвоночных организмов. Широко представлены личинки ручейников, моллюски. По численности, биомассе и видовому составу в бентосе доминируют личинки хирономид. Средняя биомасса бентоса – 2,10 г/м 2 , максимальный показатель – 12,36 г/м 2 [13].
Анализ результатов показал, что из свободных аминокислот мышечной ткани сига обыкновенного из озер Умбозеро и Ловозеро превалируют лизин, аланин, гистидин, тирозин и глицин.
Обнаружены различия в количественном отношении содержания свободных аминокислот в изучаемых объектах. Свободные аминокислоты мышечной ткани рыб пластичны и претерпевают изменения в период полового созревания [8].
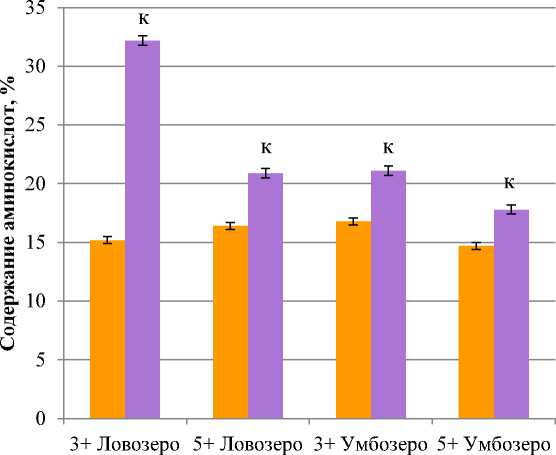
Отношение основных аминокислот к кислым в мышечной ткани сига оз. Умбозеро меньше (для четырехлеток – 1,3, для шестилеток – 1,2), чем у сига оз. Ловозеро (2,1 и 1,3 соответственно) (рис. 2). Такие различия можно объяснить разным характером питания рыбы.
Повышение интенсивности синтеза белка в мышцах сопровождается значительным уменьшением количества свободных аминокислот. Выявлено, что с возрастом суммарное содержание незаменимых свободных аминокислот от общего пула свободных аминокислот заметно снижается у сига оз. Ловозеро: у четырехлеток и шестилеток данного водоема оно составляет 49,5 и 34,1 % соответственно; у сигов оз. Умбозеро – 41,6 и 35,7 % (рис. 1). При половом созревании сига происходит уменьшение общего пула незаменимых свободных аминокислот за счет лизина, лейцина, изолейцина, фенилаланина, валина и аргинина, особенно снижается количество лизина. Именно эта аминокислота активно участвует в обмене веществ. При развитии организма лизин интенсивно расходуется на увеличение массы тела. У взрослых рыб, в отличие от растущих, потребность в этой аминокислоте значительно снижена.
■ кислые АК
■ основные АК
Рис. 2. Содержание кислых и основных аминокислот в мышечной ткани сига обыкновенного Coregonus lavaretus L., % к общей сумме аминокислот;
к – достоверность различий по сравнению с кислыми аминокислотами, p ≤ 0,05
Fig. 2. The сontent of acidic and alkaline aminoacids in the muscle tissue of Coregonus lavaretus L., % of total aminoacids; к – reliability of differences compared to acidic aminoacids, p ≤ 0,05
Заключение
-
1. Белки мышечной ткани сига оз. Умбозеро в отличие от сига оз. Ловозеро отличаются пониженным содержанием незаменимых для рыб аминокислот изолейцина и валина. Вероятно, недостаток этих незаменимых аминокислот приведет к нарушению метаболизма у исследуемых рыб оз. Умбозеро. Значительно повышается содержание незаменимой аминокислоты лейцина в мышечной ткани сига оз. Ловозеро, что можно объяснить увеличением концентрации солерастворимой миофибриллярной фракции.
-
2. Отношение свободных основных аминокислот к кислым в мышечной ткани сига оз. Умбозеро меньше, чем у сига оз. Ловозеро, что, возможно, объясняется разным характером питания рыбы. При половом созревании сига происходит уменьшение общего пула незаменимых свободных аминокислот, особенно это заметно у сига оз. Ловозеро. Снижение количества свободных аминокислот сопровождается повышением интенсивности синтеза белка в мышцах в связи с ростом рыбы.
Список литературы Аминокислотный состав мышечной ткани сига обыкновенного Coregonus lavaretus L. Кольского полуострова
- Хочачка П., Сомеро Д. Биохимическая адаптация. М.: Мир, 1988. 568 с.
- Мишанина (Похольченко) Л. А. Тканевые белки и липиды у дикой и заводской молоди атлантического лосося Salmo salar L. Кольского полуострова: дис. … канд. биол. наук. Петрозаводск, 2011. 120 с.
- Гнедов А. А., Кайзер А. А. Качественные показатели сига сибирского (Coregonus lavaretus pidschian gmelin) низовий бассейна реки Енисей//Вестник Красноярского государственного аграрного университета. 2009. № 9 (36).- С. 162-166.
- Двинин Ю. Ф., Двинин М. Ю. Химический состав и биохимические свойства тканей и органов некоторых пресноводных рыб Мурманской области/Гос. ком. Рос. Федерации по рыболовству. Поляр. науч.-исслед. ин-т морского рыб. хоз-ва (ПИНРО). Мурманск: ПИНРО, 2002. 44 с.
- Абрамов А. Ф., Слепцова Т. В., Ефимова А. А. Пищевая и биологическая ценность чира Coregonus nasus (Pallas) индигирской популяции в Республике Саха (Якутия)//Наука и образование. 2016. № 1. C. 91-95.
- Brent L., Frederick W. A method for quantitative amino acid analysis using precolumn o-Phthalaldehyde derivatization and high performance liquid chromatography//Journal of Chromatographic Science. 1991. V. 19 (5). P. 259-265.
- Антипова Л. В., Глотова И. А., Рогов И. А. Методы исследования мяса и мясных продуктов. М.: Колос, 2001. C. 69-71.
- Джабаров М. И. Аминокислотный состав тканей различных видов рыб в онтогенезе и при изменении экологических условий. М.: ВНИРО, 2006. 213 с.
- Решетников Ю. С. Экология и систематика сиговых рыб. М.: Наука, 1980. 300 с.
- Решетников Ю. С. Современные проблемы изучения сиговых рыб//Вопросы ихтиологии. 1995. Т. 35, № 2. С. 156-174.
- Решетников Ю. С., Лукин А. А. Сиговые рыбы//Биоресурсы Онежского озера. Петрозаводск: КарНЦ РАН, 2008. С. 121-137.
- Журавлев В. Б., Ломакин С. Л., Решетников Ю. С. Морфоэкологическая характеристика обыкновенного сига Coregonus lavaretus (L.) озера Сорулукель в Республике Алтай//Экология. 2014. № 5. С. 376-384.
- Ежегодник состояния экосистем поверхностных вод России (по гидробиологическим показателям) за 2014 год/под науч. ред. В. М. Хромова. М.: Институт глобального климата и экологии Федеральной службы России по гидрометеорологии и мониторингу окружающей среды и РАН, 2015. 168 с.
