An initial investigation of Otocarpus virgatus seeds germination an endemic, rare and threatened plant of Algeria

Author: Djebbouri Mohammed, Terras Mohamed, Zouidi Mohamed
Journal: Журнал стресс-физиологии и биохимии @jspb
Article in issue: 1 т.18, 2022.
Free access
Otocarpus virgatus Durieu is an endemic plant, very rare to a very narrow distribution around the region of Saïda (Algeria) critically endangered, it has indehiscent fruits. The objective of this work is to study the influence of its fruit pericarp on the germination of seeds and effects of different concentrations of gibberellic acid (GA3) on the germination rate of isolated seeds or seeds in indehiscent siliques. The results of this study indicate that O. virgatus seeds have shallow physiological dormancy and pericarp significantly reduced germination, inhibition was not due to lack of water absorption by seeds but to chemical inhibitors and mechanical resistance .Exclusively in isolated seeds all concentrations of GA3 significantly increased the germination percentage and reduced the average germination time and no significant difference in the final germination percentages for the inside of the indehiscent siliques deals with the GA3. Our data provided useful information on germination protocols for the ex situ propagation of this endemic, rare and threatened species, being the first report on germination behaviour of seeds of this taxon.
Otocarpus virgatus durieu, germination, seeds, pericarp, threatened species, conservation
Short address: https://sciup.org/143178332
IDR: 143178332
Text of the scientific article An initial investigation of Otocarpus virgatus seeds germination an endemic, rare and threatened plant of Algeria
More than three-quarters (77.9%) of the strict endemic taxa of Algeria where sub-endemic plants are more or less rare in Algeria, the endemic ones more or less common represent less than a quarter of the total (Véla and Benhouhou, 2007). Rare plants have a high conservation value, either for heritage reasons or for a probability of extinction (Pimm et al. , 1988; Gaston, 1991).
The genus Otocarpus is an endemic annual plant, very rare with a narrow distribution around the Saida region (Algeria) represented by a single species (Quézel and Santa, 1961-1962), it does form a singular taxonomic essence as confirmed by phylogenetic studies of the Brassiceae tribe of which it is a member (Warwick and Black, 1997; Warwick and Sauder, 2005). This genus is represented by a single species (Quézel and Santa, 1961-1962).
Several authors have reported this plant in the clay fields and pastures of the low semi-arid mountains and highlands of Saida and even in the nearby Tiaret region of Saida (Trabut, 1887; Maire, 1965; Miara et al. , 2014; Djebbouri and Terras, 2019). This species has been assessed as critically endangered, based on IUCN criteria (Walter and Gillett, 1998) .
Ex situ conservation of these species is considered an appropriate means to reduce the risk of extinction of threatened and endangered species (Rajjou and Debeaujon, 2008; Kirmizi et al. , 2011) and in these conservation programs seed germination studies are essential tools (Ortega-Baes and Rojas-Aréchiga, 2007). In addition and according to our knowledge no information available in the literature on the germinative behaviour of seeds of this endemic narrow species.
At the O. virgatus the Fruit is an indehiscent silique, does not open at the time of maturity, and forms the unit of dispersal and germination to ensure the regeneration of the plant (Quézel and Santa, 1961-1962; Miara et al. , 2014).
The objective of this work is to study the influence of its fruit pericarp on seed germination and the effects of different concentrations of gibberellic acid (GA3) on seed and silique germination (Fig 02), to provide useful information on the germination protocols of these siliques for the ex situ multiplication of this endemic, rare and threatened species.
MATERIALS AND METHODS
Influence of pericarp on water permeability
To study the water permeability of the pericarps, three replicates of 20 intact siliques and 20 seeds of Virgatus were weighed before and after a period of soaking in distilled water for 48 hours (siliques and seeds soaked in water have been dried with filter paper and weighed until the mass is constant, i.e., maximum absorption).
The percent mass increase (% Wr) was calculated as follows:
Wr % = [(Wi - Wd) / Wd]*100
Wi: mass of soaked silks or seeds; Wd: mass of air dried silks or seeds (Lu et al. 2015).
To examine the effect of pericarp on water absorption by the seeds, the seeds soaked inside in their pericarp were removed and weighed, their percentage increase in mass was compared with isolated seeds immersed in water.
Effect of pericarp on germination
To test the influence of pericarp on germination, several tests were carried out as follows: i) a control test with siliques fruit that has been placed in petri dishes to verify the mechanical effect of pericarp on germination. ii) A test by isolated seeds separated from the silica was placed in Petri dishes to assess the embryonic dormancy of the seeds. iii) for the last test isolated seeds placed next to each other at about 1 cm are used to test the chemical effect of the pericarps on the germination of the seeds.
Action of gibberellic acid (GA3) on germination
To test the effect of the plant hormone gibberellic acid on germination two tests were carried out: the first one allowed the use of the complete fruit immersed in the hormone for 48 hours at 4 different PPM concentrations (0, 250, 500, 750). The twelfth is to use isolated seeds immersed in GA 3 for 48 hours at the same previous concentrations.
The germination parameters measured
Two parameters were calculated at 15 days of germination initiation (germination was stabilized on day 9):
The germination percentage (GP) is the best indicator to identify the effects of the pericarp and the different concentrations of gibberellin acid on the physiological germination of seeds. The germination percentage (GP) is calculated as follows:
GP = (number of seeds sprouted / total number of seeds) 100.
Average germination time (MTG) according to the formula given by (Ellis and Roberts 1981) .
MGT = ∑ n × D/∑ n
GP = (number of germinated seeds / total number of seeds) × 100.; Σn: number of seeds that germinated during the germination test. This means that the lower the value, the faster the germination (Melendo and Giménez, 2019).
Statistical analysis
All data were analyzed for normality and homogeneity of variances in advance, with mean values for each experiment corresponding to the average of three replicates of 20 seeds/siliques ± standard error (ES).
Independent sample t-tests were used to compare the means of data on the effect of pericarp on water absorption.
A one-way ANOVA was used to analyze the averages of the pericarp effect data and GA 3 on germination, to compare the averages the Duncan test was used and the level of significance was set at 0.05.
RESULTS
Effect of pericarp on water absorption
The statistical treatment result showed significant differences (P<0,01) for the mass of siliques before and after immersion in water, with a percentage increase of 118 % ± 9%.
On the other hand, no significant difference (P>0.05) was recorded for the percentage increase in soaked seeds without pericarps and soaked seeds inside pericarps, the increase in seed mass was respectively 41.6% ± 0.03%, 40.2% ± 0.03%.
Effect of Pericarp on Germination
The results of Table 1 show that the comparison of germination rates shows the presence of a significant difference (P<0,05) between fruits, isolated seeds and seeds isolated and incubated in the presence of pericarps (Fig 03).
Elimination of pericarp increased germination from 5% to 38.33%, and the presence of pericarps significantly decreased the rate of germination.
GA 3 effect on germination
Silique
No significant difference in final germination percentages for seeds inside Silique indehiscents immersed in GA 3 for 48 hours at 4 concentrations (ppm) 0 250 500 750. (All > 0.05). (Table 02).
Isolated seeds
Compared to the controls, the percentage of germination was significantly increased by GA 3 , depending on its concentration (Fig. 1). The best germination rates were recorded in GA 3 treatment at 750 ppm followed by 500 and 250 ppm. All GA 3 concentrations decreased similarly the average germination time, compared to the control treatment.
Table 1. Effect of Pericarp on Germination
|
tests |
germination rate % |
TMG |
|
Intact fruit(siliques) |
5.00 A |
- |
|
Incubated seeds in the presence of detached pericarpes |
20.00 B |
2.83 |
|
Isolated seeds |
38.33 C |
4.03 |
|
P Value |
0.02 |
0.13 |
In each column, values with different letters are significantly different according to a Duncan test (˛ = 0.05), P value from an analysis of variance.
Table 2. Final germination percentages for seeds inside indehiscent siliques.
|
tests |
germination rate % |
TMG |
|
Silique (control) |
5.00 |
- |
|
Silique submerged in a concentration of 250 ppm of GA 3 |
8.33 |
- |
|
Silique submerged in a concentration of 500 ppm of GA 3 |
8.33 |
- |
|
Silique submerged in a concentration of 750 ppm of GA 3 |
16.67 |
2.24 |
|
P Value |
0.29 |
- |
In each column, values with different letters are significantly different according to a Duncan test (˛ = 0.05), P value from an analysis of variance.
Figure 1. A- silique. B- seeds .

Figure 2. Germination of seeds in their indehiscent siliques (pericarp).
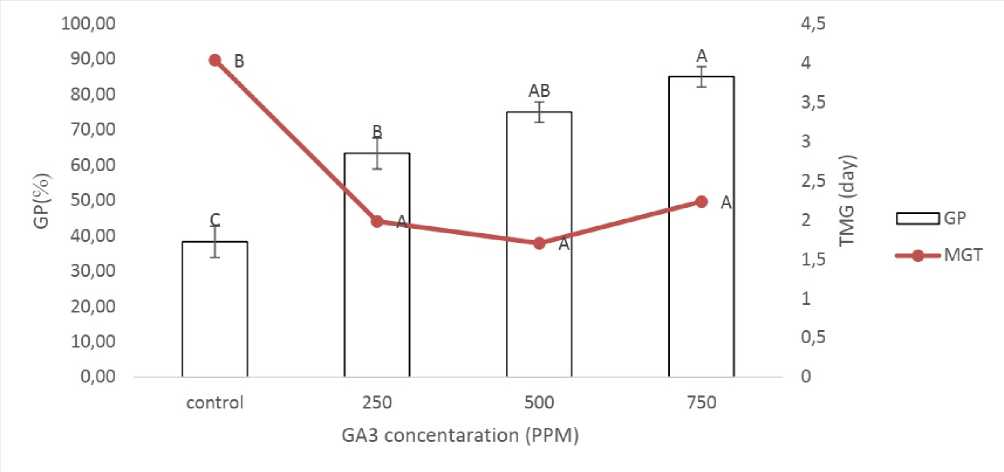
Figure 3. Effect of GA 3 on the germination of isolated seeds
DISCUSSION
In many species the pericarp is indehiscent, it does not open spontaneously, at the time of maturity to ensure the regeneration of the plant, and the fruit forms the unit of dispersion and germination. Among these species, Otocarpys virgatus which is a strict endemic species of the HautsPlateaux of western Algeria, very rare and unknown species (Cousens et al. , 2009; Lu et al. , 2010; Ohadi et al. , 2011; Miara et al. , 2014; Eskandari et al. , 2017).
Comparison of the mass of the siliques before and after soaking in water shows a significant difference, in addition, no significant difference was recorded between the percentage increase in the mass of seeds soaked in water without pericarps and seeds immersed inside pericarps, therefore the inhibitory effect of the pericarp is not due to their impermeability. This confirms that neither the pericarp nor the seed shell prevented the passage of water through the seed to the embryo and the inhibition of germination. Similar results were found in two other annual species of Brassicaceae : Raphanus raphanistrum (Cousens et al. , 2009) and Diptychocarpus strictus (Lu et al. , 2010).
The study of the effect of the indehiscent pericarp of Otocarpys virgatus on germination indicates that the rate of germination is decreased as long as the seeds are inside the fruits but the removal of the pericarp to significantly increase the percentages of germination, this hypothesis that pericarp acts by providing both mechanical resistance and chemical inhibitors was confirmed in other Brassicaceae species (Lu et al., 2015; Eskandari et al., 2017).
The use of GA 3 in our experiment significantly increases the germination rate and significantly decreases the average germination time in isolated seeds, these germination-enhancing effects of GA 3 have been reported in many species according to several authors(Honarmand et al. 2016; Eskandari et al. , 2017; Lu et al. , 2017), in our experience the dormancy of the isolated seeds was broken by GA 3 , these results show that the seeds of Otocarpys virgatus have a shallow physiological dormancy (Baskin and Baskin, 2004).
Statistically, the results show that the seeds treated with GA3 that are inside the indehiscent siliques (siliques immerged in GA3 at different concentrations do not affect the percentage of germination). The pericarp inhibited the germination of non-dormant seeds, that is, even after the seeds have lost their physiological dormancy, they cannot break the wall of the fruit and germinate, the same results in others Brassicaceae species have been confirmed that the pericarp inhibits the germination of non-dormant seeds inside indehiscent fruits, regardless of the treatment that ended dormancy (Mamut et al., 2014; Lu et al., 2015; Lu et al., 2017), because the fruits of some species of Brassicaceae are hard and did not open easily which delays or inhibits germination even after lifting physiological dormancy, and fruit walls could prevent germination by mechanical restriction or chemical inhibition (Cheam 1986; Cousens et al., 2009; Mamut et al., 2014).
CONCLUSION
The results of this study show that the pericarp had a significant effect on the germination of seeds of O. virgatus through mechanical and chemical inhibition thus preventing early germination. Removal of the pericarp and treatment of the kernels with GA 3 significantly improved germination rates. This information made it possible to provide certain details concerning the germination of the seeds of Otocarpus virgatus , an original and endemic plant of western Algeria, long forgotten, and whose evolutionary, biogeographic and heritage importance is undeniable.
CONFLICTS OF INTEREST
The authors declare that they have no potential conflicts of interest.
References An initial investigation of Otocarpus virgatus seeds germination an endemic, rare and threatened plant of Algeria
- Baskin J.M. and Baskin C.C. (2004) A classification system for seed dormancy. Seed Science Research, 14:1-16.
- Cheam A. (1986) Seed production and seed dormancy in wild radish (Raphanus raphanistrum L.) and some possibilities for improving control, Weed Research. 26:405-414.
- Cousens R.D., Young K.R. and Tadayyon A. (2009) The role of the persistent fruit wall in seed water regulation in Raphanus raphanistrum (Brassicaceae), Annals of botany. 105:101-108.
- Djebbouri M. and Terras M. (2019) Floristic diversity with particular reference to endemic, rare or endangered flora in forest formations of Saida (Algeria). International Journal of Environmental Studies, 76:990-1003.
- Ellis R. and Roberts E. (1981) The quantification of ageing and survival in orthodox seeds. Seed Science and Technology (Netherlands), 9: 373409.
- Eskandari B.S., Ghaderian S.M., Ghasemi R. and Schat H. (2017) Optimization of seed germination in an Iranian serpentine endemic, Fortuynia garcinii. Flora. 231:38-42.
- Gaston K.J. (1991) How large is a species' geographic range? Oikos, 434-438.
- Honarmand S.J., Nosratti I., Nazari K. and Heidari H. (2016) Factors affecting the seed germination and seedling emergence of m uskweed (M yagrum perfoliatum). Weed biology and management, 16:186-193.
- Kirmizi S., GULERYUZ G. and ARSLAN H. (2011) Germination responses to GA3 and short-time chilling of three endemic species: Tripleurospermum pichleri, Cirsium leucopsis and Senecio olympicus (Asteraceae). Plant Species Biology, 26:51-57.
- Lu J., Tan D., Baskin C. and Baskin J. (2017) Delayed dehiscence of the pericarp: role in germination and retention of viability of seeds of two cold desert annual Brassicaceae species. Plant Biology, 19:1422.
- Lu J., Tan D., Baskin J.M. and Baskin C.C. (2010) Fruit and seed heteromorphism in the cold desert annual ephemeral Diptychocarpus strictus (Brassicaceae) and possible adaptive significance. Annals of botany, 105:999-1014.
- Lu JJ, Zhou Y.M., Tan D.Y., Baskin C.C. and Baskin J.M. (2015) Seed dormancy in six cold desert Brassicaceae species with indehiscent fruits. Seed Science Research, 25:276-285.
- Maire R. (1965) Flore de l'Afrique du Nord: Dicotyledonae: rhoeadales: papaveraceae, sf. Fumarioidea pp; capparidaceae, cruciferae pp. Lechevalier, Paris, France.
- Mamut J., Tan D.Y., Baskin C.C. and Baskin J.M. (2014) Role of trichomes and pericarp in the seed biology of the desert annual Lachnoloma lehmannii (Brassicaceae). Ecological research, 29:33-44.
- Melendo M. and Giménez E. (2019) Seed germination responses to salinity and temperature in Limonium supinum (Plumbaginaceae), an endemic halophyte from Iberian Peninsula. Plant Biosystems-An International Journal Dealing with all Aspects of Plant Biology, 153:257-263.
- Miara M.D., Hammou M.A., Hadjadj-Aoul S. and Rebbas K. (2014) Redécouverte d'Otocarpus virgatus Durieu (Brassicaceae) dans la région de Tiaret (nord-ouest de l'Algérie). Bull. Soc. linn. Provence, 65:31-31.
- Ohadi S., Mashhadi H.R. and Tavakol-Afshari R. (2011) Effects of storage and burial on germination responses of encapsulated and naked seeds of turnipweed (Rapistrum rugosum) to light. Weed science, 59:483-488.
- Ortega-Baes P. and Rojas-Aréchiga M. (2007) Seed germination of Trichocereus terscheckii (Cactaceae): light, temperature and gibberellic acid effects. Journal of arid environments, 69:169-176.
- Pimm S.L., Jones H.L. and Diamond J. (1988) On the risk of extinction. The American Naturalist, 132:757-785.
- Quézel P. and Santa S. (1961-1962) Nouvelle flore de l'Algérie et des régions désertiques. Centre nat edition, Paris.
- Rajjou L. and Debeaujon I. (2008) Seed longevity: survival and maintenance of high germination ability of dry seeds. Comptes Rendus Biologies, 331:796-805.
- Trabut L. (1887) D'Oran a Mécheria: notes botaniques & catalogue des plantes remarquables, Jourdan, Alger.
- Véla E, Benhouhou S(2007) Évaluation d'un nouveau point chaud de biodiversité végétale dans le Bassin méditerranéen (Afrique du Nord). Comptes Rendus Biologies, 330:589-605.
- Walter K.S. and Gillett H.J. (1998) 1997 IUCN Red List of threatened plants, IUCN, Gland.
- Warwick S. and Black L. (1997) Phylogenetic implications of chloroplast DNA restriction site variation in subtribes Raphaninae and Cakilinae (Brassicaceae, tribe Brassiceae). Canadian Journal of Botany, 75:960-973.
- Warwick S.I. and Sauder C.A. (2005) Phylogeny of tribe Brassiceae (Brassicaceae) based on chloroplast restriction site polymorphisms and nuclear ribosomal internal transcribed spacer and chloroplast trn L intron sequences. Canadian Journal of Botany, 83:467-483
