Анализ двигательной активности и температурная устойчивость бычка Myoxocephalus scorpius (L.)
")
Автор: Муравейко Александр Владимирович
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Статья в выпуске: 1 т.17, 2014 года.
Бесплатный доступ
Рассматриваются особенности изменений двигательной активности баренцевоморского керчака Myoxocephalus scorpius как перспективного объекта промысла. Активность определялась в лабораторных условиях при непрерывной регистрации. Анализировалось влияние резких изменений температуры на изменчивость биоритмики бычка. Оценено влияние геомагнитных возмущений на характер ритмики.
Баренцевоморский керчак, двигательная активность, температурная устойчивость, биоритмика поведения, геомагнитные возмущения
Короткий адрес: https://sciup.org/14294665
IDR: 14294665
Текст научной статьи Анализ двигательной активности и температурная устойчивость бычка Myoxocephalus scorpius (L.)
Европейский керчак Myoxocephalus scorpius широко распространен в прибрежных водах Баренцева и Белого морей. Летом керчак держится ближе к берегу, а на зиму отходит на большую глубину. Далеких миграций он не совершает и образует местные стада, которые отличаются незначительными морфологическими особенностями. Ранее считалось, что промыслового значения эта рыба не имеет ( Никольский , 1950). Обычно бычки добываются в незначительном количестве в качестве прилова при траловом промысле донных рыб. При специализированном или селективном промысле их уловы можно увеличить в несколько раз ( Торканов , 1985). Для увеличения промыслового изъятия керчака важное значение приобретают исследования суточной ритмики активности этих рыб, позволяющие более точно организовать добычу ( Кручинин и др. , 1981). В связи с отсутствием данных о биоритмах бычка-керчака было начато изучение суточных ритмов активности этих рыб. Кроме того, задачей работы являлось исследование устойчивости керчака к изменениям температуры воды.
Известны зависимости объемов вылова и поведения гидробионтов от космогеофизических факторов – вариаций магнитного поля Земли (МПЗ) и солнечной активности ( Степанюк , 2002; Чижевский , 1973; Владимирский и др. , 2004; Зимин , 2002 и др.). В силу этого, актуально изучение поведения керчака в условиях изменчивости данных факторов.
Что касается исследований двигательной активности рыб, то такие работы имеют не только прикладное значение, но важны также для фундаментальной науки, поскольку позволяют оценивать воздействие космогеофизических факторов на биосферу. Общеизвестны факты влияния на биологические объекты солнечной активности, геомагнитных бурь, лунных фаз, изменения полярности межпланетного магнитного поля и др., хотя механизмы такого влияния во многом остаются малоисследованными ( Степанюк , 2002; Чижевский , 1973; Владимирский и др. , 2004). В качестве критерия оценки воздействия этих факторов часто используется двигательная активность животных.
2. Методика проведения экспериментов
В качестве критерия биоритмики керчака использовалась его двигательная активность. Для этих целей к спинному плавнику рыбы прикреплялась тонкая резиновая нить. Другой конец нити крепили к тензодатчику, последний через мостовую схему подключали к аналоговому регистратору Н-338 4П. Запись с помощью фотодатчика преобразовывали в электрические потенциалы, которые являлись условными единицами двигательной активности. Запись двигательной активности рыб осуществляли при естественном освещении и постоянной температуре морской воды 13 °С. В опытах использовали 3-4-летних рыб. Эксперименты проводили в экранированной от электромагнитных полей камере.
На рис. 1 представлены образцы суточных записей двигательной активности бычков. Непрерывную регистрацию активности керчака проводили в течение 3 суток. В первые сутки рыба проходила период адаптации к условиям содержания в экспериментальном аквариуме. Уровень ее двигательной активности был достаточно высок в дневное и ночное время. На 2-е и последующие сутки отчетливо заметно увеличение двигательной активности в ночное и существенное снижение активности в дневное время.
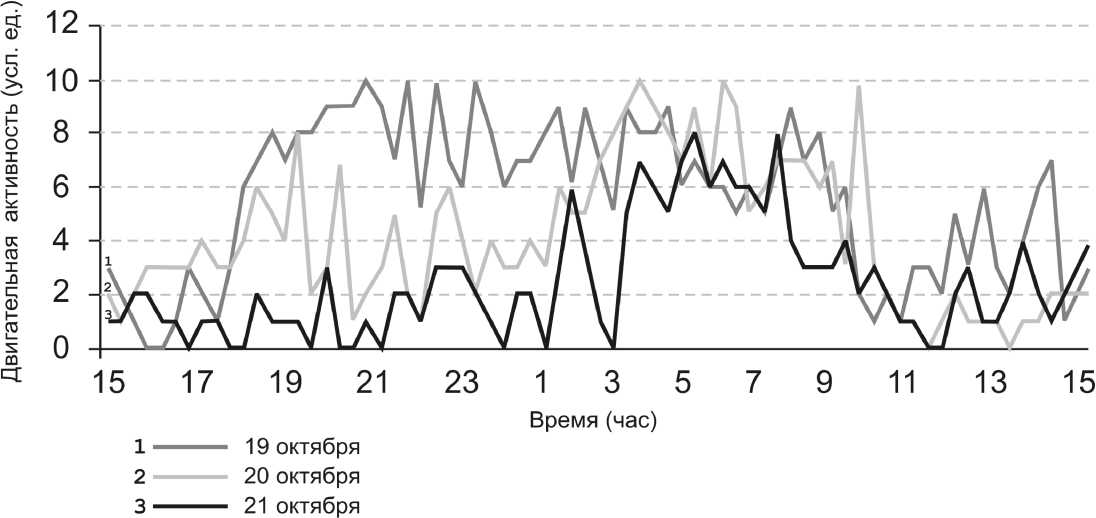
Рис. 1. Суточная динамика двигательной активности бычка-керчака 19 октября (кривая 1), 20 октября (кривая 2) и 21 октября 2007 г. (кривая 3)
Однако это не означает, что бычок-керчак является ночным хищником. Он является хищником-засадчиком, прекрасно маскируется под окружающий фон и, будучи неважным пловцом, в дневное время атакует только близко проплывающую добычу. В ночное время бычок активно передвигается по дну и пер еходит на питание бентосными животными ( Никольский , 1950).
На следующем этапе работ выявляли устойчивость рыбы к резкому повышению и снижению температур окружающей водной среды. Для этих целей использовали две емкости: аквариум с нагретой до 13 °С водой и аквариум с морской водой, охлажденной до температуры +2 °С. Животных быстро пересаживали из одной емкости в другую и наблюдали за их выживаемостью. На протяжении всего эксперимента регистрировали двигательную активность. Охлажденные рыбы в течение длительного времени (около 25 мин) оставались малоподвижными. После того, как животные начали двигаться, их помещали в теплую ванну, где они сохраняли высокий ур овень двигательной активности (рис. 2). При помещении животных в ванну с отрицательной темпер атурой воды до –2 °С, они погибали в течение 5 минут.
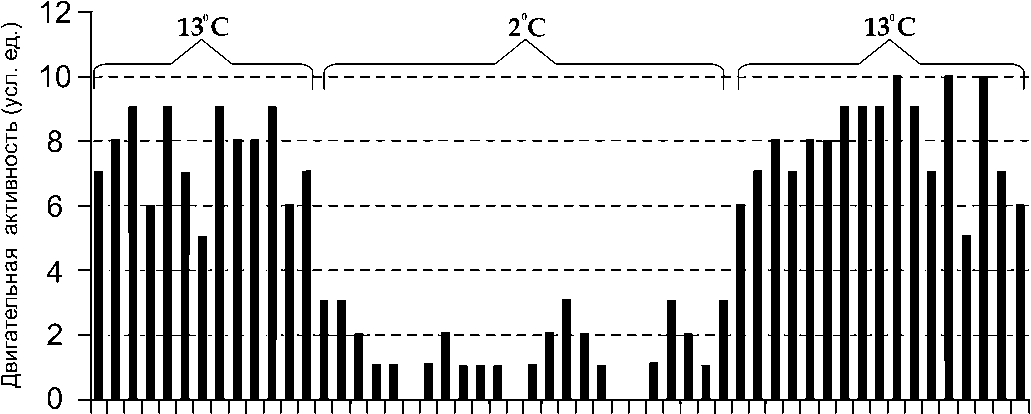
Рис . 2. Влияние резкого снижения и повышения температур воды на двигательную активность бычка-керчака. Шаг гистограммы по оси х – 1 мин
3. Методики обработки данных при выявлении биоритмики
Для обработки полученных данных использовались стандартные пакеты MESOSAUR, HERST и Excel.
Для начала ряды двигательной активности были профильтрованы полосно-пропускающим фильтром. Это необходимо для выделения из данных полезного сигнала при условии, что известен частотный диапазон сигнала. Полосно-пропускающий фильтр характеризуется двумя частотами и выделяет в качестве сигнала все составляющие с частотами в заданной полосе.
По отфильтрованным данным рассчитывались периодограммы, функции спектральной плотности (автоспектры), кросс-спектры, функция когерентности и фаза. Для определения степени детерминированности поведения рыб в период эксперимента производились вычисления фрактальной размерности процесса ( Федер , 1991). Размерность связана с так называемым показателем Херста H и определяется как разность (2 – H ). Показатель Херста описывается эмпирическим соотношением:
H = log (τ/2) ( R / S ), (1)
где H – показатель Херста; τ – временное "окно", ч.; R – размах, т.е. разность максимального и минимального значений за рассматриваемый интервал времени; S – стандартное отклонение.
Для процесса независимых испытаний H = 0,5. При H > 0,5 временной ряд имеет персистентное (поддерживающееся) поведение, т.е. процесс является детерминированным. При H < 0,5 – поведение хаотично.
4. Обсуждение результатов
Полученные результаты свидетельствуют о высокой устойчивости бычка-керчака к резким перепадам температуры окружающей водной среды с 13 до 2 °С, в этих опытах ни одно из 7-ми животных не погибло. Однако рыбы не выдерживают отрицательных температур.
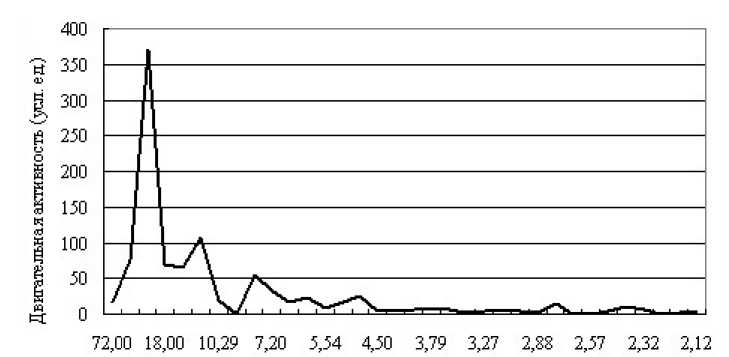
Периодограмма отфильтрованного ряда активности керчака приведена на рис. 3. Отчетливо выделяются следующие периоды активности: 24, 12, 8, 6 часов.
Для выяснения вопроса, являются ли движения рыбы хаотичными или закономерными, был рассчитан Н -показатель (показатель Херста) (рис. 4). При расчете задавался шаг 1 и окно 40 значений. Как следует из этих данных, движения рыбы в течение всего эксперимента однозначно являлись закономерными ( Н -показатель выше 0,5) и определялись внутренними физиологическими потребностями.
Период,ч
Рис. 3. Периодограмма отфильтрованного ряда суточной активности керчака
Также можно отметить, что вариации коэффициента Херста являются квазипериодическими. В период эксперимента выделены три минимума детерминированности (уменьшение Н -показателя до 0,60,65) и, соответственно три максимума со значением Н -показателя примерно 0,9. Все это, несомненно, определяется ритмикой. Минимумы наблюдаются в послеполуденное время.
При поиске связи двигательной активности с геомагнитными возмущениями МПЗ были произведены аналогичные расчеты.
На рис. 5 изображены периодограммы активности бычка и индукции МПЗ. Наблюдаются следующие периоды индукции: 18; 12; 7,2; 6 часов. У рядов активности и индукции существуют совпадающие периоды, такие как 12 и 6 часов.
Более подробно связь между рядами описывают функции кросс-спектра и когерентности, результаты расчета которых сведены в таблицу. Наглядные графики распределения кросс-спектра и функции когерентности представлены на рис. 6.
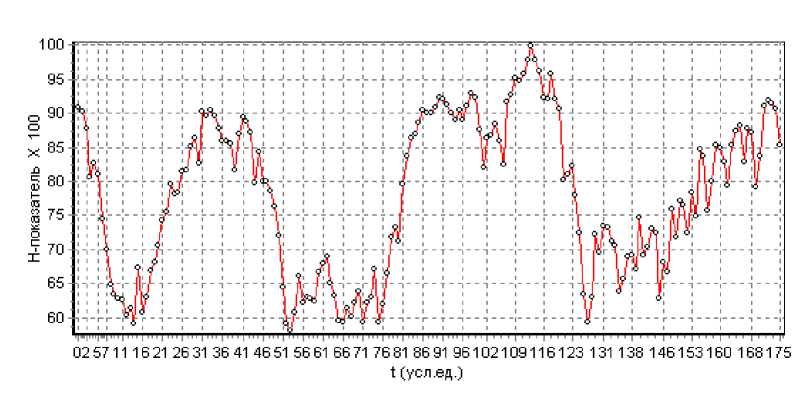
Рис. 4. Показатель Херста ( Н -показатель), рассчитанный по двигательной активности бычка-керчака
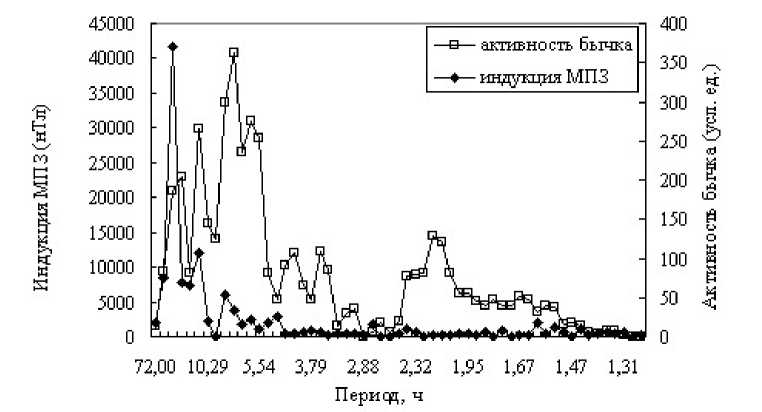
Рис. 5. Периодограммы активности бычка-керчака и индукции МПЗ
Таблица. Сравнение когерентных величин полученных данных
|
Периоды активности, ч. |
Периоды индукции МПЗ, ч. |
Периоды в кроссспектре, ч. |
Функция когерентности, усл.ед. |
Сдвиг по фазе между активностью и индукцией, ч. |
|
24,00 |
18,00 |
28,80 |
0,74 |
12,23 |
|
12,00 |
12,00 |
11,00 |
0,51 |
0,78 |
|
8,00 |
7,20 |
8,00 |
0,75 |
3,08 |
|
6,00 |
6,00 |
6,00 |
0,96 |
0,31 |
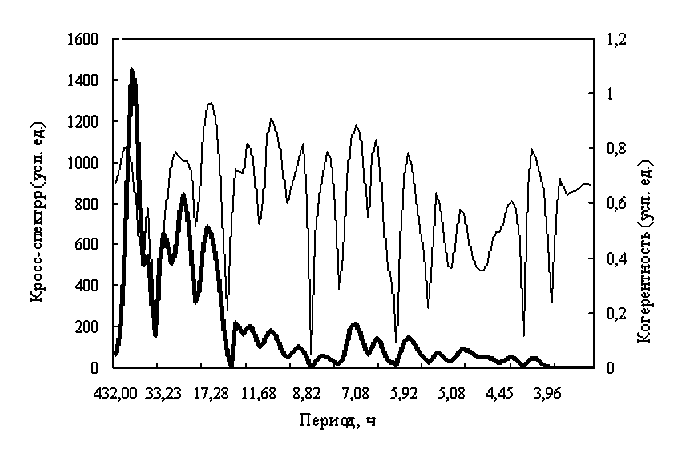
Рис. 6. Кросс-спектр и функция когерентности между рядами активности бычка и индукцией МПЗ. Жирная линия – кросс-спектр, тонкая – функция когерентности
Сдвиги по фазе между активностью и индукцией МПЗ рассчитаны в часах по формуле:
т ( 180 -H 1
где T - период, ч.; ф - фаза (берется по модулю); Ат - сдвиг по времени, ч.
Самая большая связь между рядами активности баренцевоморского бычка и индукцией наблюдается при периоде 6 часов, функция когерентности при этом равна 0,96 и наблюдается самое маленькое отставание фаз, по сравнению с другими периодичностями, составляющее 0,31 часа.
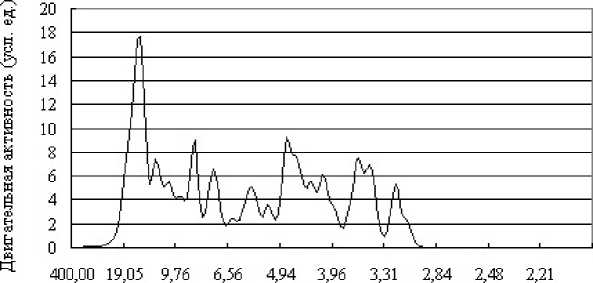
Кроме этого, для определения степени влияния МПЗ на двигательную активность керчака был построен спектр ошибок ряда активности, изображенный на рис. 7.
Период, ч
Рис. 7. Спектр остатков ряда активности бычка
Высокие значения ошибок приходятся на следующие периоды: 5; 2,7; 1,6; 1,2 ч. Ни один из вышеперечисленных периодов максимальных остатков не совпадает с периодами взаимосвязи рядов активности и индукции. Из этого следует, что корреляция индукции МПЗ и двигательной активности животного не является предопределяющим для жизнедеятельности фактором и скорее служит источником информации для "настройки" внутренней биоритмики керчака, а также получения прогностической информации о происходящих в окружающем пространстве процессах.
5. Выводы
Анализ экспериментально полученных данных позволяет уверенно утверждать, что двигательная активность бычков не является хаотичной, а жестко детерминирована. Биоритмика активности керчаков по данным наблюдений содержит суточные, полусуточные и четвертьсуточные периоды вариаций. Сравнение характеристик биоритмики поведения бычков с характеристиками геомагнитных возмущений показало высокую когерентность на указанных периодах. Интереснее всего оказался совпадающий период в 6 часов. При этом периоде наблюдается наибольшая связь между рядами, функция когерентности 0,96, и самое маленькое отставание по фазе – около получаса.
Выявленная цикличность, при которой наблюдается наибольшая активность керчака, позволяет выбрать оптимальное для промысла время 6, 8, 12, 24 часов.
