Анализ календарного возраста рамет грушанки круглолистной (Pyrola rotundifolia L.) при интерпретации онтогенетической структуры вида
 при интерпретации онтогенетической структуры вида")
Автор: Дубровная Светлана Алексеевна, Хуснетдинова Ландыш Завдетовна, Мавлюдова Ляйля Усмановна, Галимова Алина Раисовна, Бимкмухаметова Залия Шамилевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 2-3 т.19, 2017 года.
Бесплатный доступ
Была изучена онтогенетическая структура ценопопуляций Pyrola rotundifolia. Показано, что высокая доля растений виргинильного онтогенетического состояния в структуре ценопопуляции была обусловлена как замедленным развитием растений, так и интенсивными процессами вегетативного размножения, в ходе которого отмечалось систематическое пополнение ценопопуляции молодыми раметами. Календарный возраст растений можно использовать для корректной интерпретации онтогенетической структуры ценопопуляции. Увеличение календарного возраста, сопровождающееся последовательным переходом растений в более взрослое онтогенетическое состояние, может свидетельствовать о нормальном развитии растений. Длительное пребывание рамет в виргинильном онтогенетическом состоянии, соответствие календарного возраста рамет виргинильного и средневозрастного генеративного онтогенетических состояний может быть связано с затруднениями перехода растений к цветению.
Онтогенетическая структура, ценопопуляция, календарный возраст, рамета
Короткий адрес: https://sciup.org/148205160
IDR: 148205160 | УДК: 574.3;
Analysis of the calendar age of (Pyrola rotundifolia L.) ramets at interpretation of ontogenetic structure species
The ontogenetic structure of pricing of Pyrola rotundifolia had been studied. It is shown that a high proportion of plants of the virginal ontogenetic state in the structure of the coenopopulation can be caused both by delayed development of plants and by intensive vegetative propagation processes, during which a systematic replenishment of the cenopopulation with young ramets is noted. The chronological age of plants can be used to correctly interpret the ontogenetic structure of the coenopopulation. An increase in the chronological age, accompanied by a gradual transition of plants to a more adult ontogenetic state, may indicate normal plant development. The increase in the length of the stay of the ramet in the virginal ontogenetic state, the correspondence of the chronological age of the plants of the virgin and mid-generation generative state is associated with difficulties in the transition of plants to flowering.
Текст научной статьи Анализ календарного возраста рамет грушанки круглолистной (Pyrola rotundifolia L.) при интерпретации онтогенетической структуры вида
Галимова Алина Раисовна , студентка
Бимкмухаметова Залия Шамилевна, студентка
Т.А. Работнова [3]. Автором отмечалось, что чем условия ценоза ближе к экологическому оптимуму вида, тем больше в составе популяций плодоносящих и хорошо развитых растений. У многолетних растений это объясняется тем, что с ухудшением условий затормаживается не только накопление биомассы, но и генеративное развитие [4]. Особенности возрастного состава популяции следует рассматривать как общевидовое приспособление к удержанию пространства [5]. Вблизи оптимума растения зацветают быстрее и на фоне ускоренного накопления биомассы, так что «видовая функция» средней особи осуществляется на высоком уровне. Онтогенетический состав популяции и процессы смены поколений чрезвычайно разнообразны и зависят от средней продолжительности жизни особи, от экологических условий и т.п. [4].
Цель исследования: использование календарного возраста растений для интерпретации онтогенетической структуры ЦП грушанки круглолистной Pyrola rotundifolia L.
Методика исследования. Объектом исследования были ЦП и надземные раметы P. rotundifolia (парциальные побеги или парциальные кусты) различных онтогенетических состояний. Ю.А. Бобров [6] отмечал, что у грушанки круглолистной общая длительность жизни парциального куста составляет 18-23 лет. Для исследования были выбраны ЦП вида в неморальной зоне, подзоне хвойно-широколиственных лесов (в Республике Татарстан). Здесь в условиях долинно-террасных ландшафтов, в сосняке разнотравном, в микропонижении была обнаружена изолированная ЦП вида общей площадью 30 м2 - 1 ЦП. Особенность рельефа способствовала тому, что на данном участке совместно произрастали как растения бореальных лесов, так и пойменных лугов. Две другие ЦП были изучены в пределах бореальной лесной зоны в подзоне южной тайги в Республике Мари Эл. Это ЦП в березняке орляковом (2 ЦП) и березняке грушанковом (3 ЦП), расположенном в пойме р. Большая Кокшага. Во всех ЦП исследовались только надземные раметы. Целостность корневищ не нарушалась. Изучение онтогенетической структуры ЦП проводили традиционными методами [7]. Для выявления причин высокой доли участия рамет виргинильного онтогенетического состояния в структуре ЦП изучали процесс вегетативного размножения и анализ календарного возраста рамет. Определение календарного возраста растений проводили на основе анатомических срезов. Срезы изготовляли в основании почвенновоздушных побегов (рамет). Для грушанки круглолистной показано, что в хорошо развитой ксилеме можно четко выделить участки с крупными сосудами и участки с мелкими сосудами, которые можно сравнить с ранней и поздней древесиной, образующими годичные кольца у древесных форм [8]. В каждой ЦП было проанализировано не менее 30 рамет различных онтогенетических состояний пре-генеративного периода, число рамет генеративного периода могло быть 15- 20, что связано с низкой долей цветущих рамет в природных ценопопуляци-ях. Для оценки интенсивности вегетативного размножения использовали интегральный показатель – удельную рождаемость ЦП, это отношение числа вновь появившихся растений в сезоне к числу растений, произрастающих на участке [9].
Результаты исследования. Исследования показали, что все ЦП по классификации дельтаомега были молодыми, что связано с высоким участием в онтогенетической структуре особей преге-неративного периода (рис. 1). Доля растений генеративного периода варьировала от 7-18%. Наличие цветущих растений определяло возможность полового процесса. В то же время, единичные проростки были нами обнаружены только в березняке грушан-ковом, и все растения, изъятые для анализа случайным образом, были вегетативного происхождения. Высокое участие растений прегенеративного периода в структуре ЦП могло быть обусловлено различными причинами. Во-первых, недавним заселением местообитания, что возможно при восстановлении подходящих эколого-ценотических условий, во-вторых, хорошо выраженными процессами вегетативного размножения, в-третьих, медленным развитием растений, что ведет к накоплению в структуре ценопопуляции растений данных возрастных групп [10].
(%)
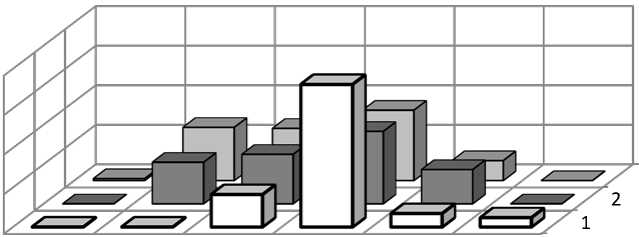
p j im v g1-g2-g3 ss
Онтогенетические состояния
Рис. 1. Онтогенетические спектры ЦП грушанки круглолистной
Основным способом самоподдержания численности всех ЦП было вегетативное размножение. Об интенсивных процессах вегетативного размножения свидетельствует показатель удельной рождаемости (табл. 1). В условиях южной тайги данный показатель варьировал от 0,4 до 0,5. Интенсивность вегетативного размножения в зоне хвойно-широколиственных лесов в сосняке была значительно ниже. В 2017 г. показатель удельной рождаемости был равен нулю.
Анализ спектра календарного возраста показал, что ЦП различаются по преобладающей возрастной группе рамет. В ЦП южной тайги высока доля молодых растений, которые соответствовали имматурному и ювенильному онтогенетическому состоянию, что и определяло высокое участие растений данной онтогенетической группы в спектре (рис. 1, 2). В зоне хвойно-широколиственных лесов
(сосняк разнотравный) отмечалась незначительная доля рамет, чей календарный возраст составил 1-3 года. Даже с учетом возможной погрешности при определении возраста растений непосредственное наблюдение показало, что в сезоне 2017 г. в сосняке раметы не появились вовсе. Учитывая и незначительную долю рамет второго года, можно говорить о затруднениях процесса самоподдержания численности ЦП грушанки в зоне хвойно-широколиственных лесов.
Таким образом, одной из причин высокого участия рамет прегенератиного состояния в онтогенетических спектрах является хорошо выраженный процесс вегетативного размножения. Однако это не объясняет преобладание в онтогенетической структуре ЦП растений вирнинильного онтогенетического состояния.
Таблица 1. Изменчивость демографических показателей ценопопуляции P. Rotundifolia
|
Показатели Местообитания |
ДельтаОмега |
Тип ЦП |
g2/v |
Удельная рождаемость |
Плотность |
|
сосняк разнотравный |
0,22-0,39 |
молодая |
0,09 |
0 |
8,6 |
|
березняк орляковый |
0,23-0,45 |
молодая |
0,5 |
0,4 |
20 |
|
березняк грушанковый |
0,21-0,41 |
молодая |
0,3 |
0,5 |
33,3 |
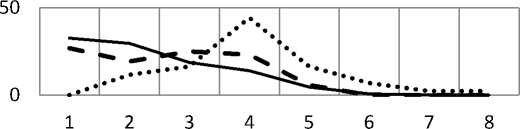
Сосняк разнотравный Березняк орляковый
Березняк грушанковый
Рис. 2. Соотношение рамет различного календарного возраста в изучаемых ЦП (спектр календарного возраста ЦП)
Анализ календарного возраста рамет вир-гинильного онтогенетического состояния. Самая многочисленная онтогенетическая группа во всех ЦП – растения виргинильного онтогенетического состояния. Анализ календарного возраста показал, что раметы достигают виргинильного онтогенетического состояния уже на второй год. В условиях светлого березняка грушанкового доля таких растений составила около 30%, в сосняке разнотравном – немногим более 3%, что говорит о более медленном развитии рамет. Отсутствие двулетних растений виргинильного онтогенетического состояния в березняке орляковом может быть связано как с малым объемом выборки для анализа, так и более медленным развитием растений (рис. 2).
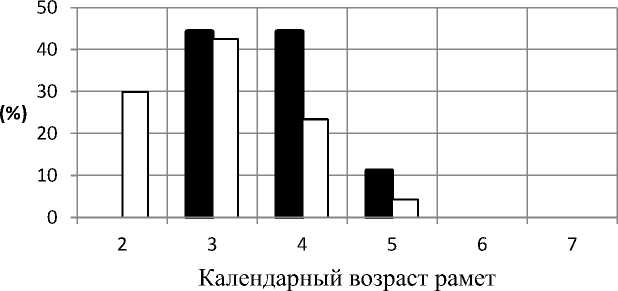
Березняк орляковый Березняк грушанковый в пойме
Рис. 3. Соотношение растений виргинильного онтогенетического состояния различного календарного возраста в изучаемых ЦП
В качестве показателя развития растений был проанализирован показатель средний возраст растений виргинильного онтогенетического состояния. Можно отметить, что в березняке гру-шанковом этот показатель был минимальным (табл. 2) и составил 3 года, в сосняке разнотравном и березняке орляковом средний показатель календарного возраста растений составил 4 года.
Преобладание растений виргинильного онтогенетического состояния в ЦП связано с длительностью пребывания в данном онтогенетическом состоянии. Максимальное пребывание растений в виргинильном онтогенетическом состоянии отмечалось в сосняке разнотравном – до 7 лет, что может свидетельствовать о затруднении перехода растений в генеративный период (рис. 2). Длительность пребывания рамет в виргиниль-ном онтогенетическом состоянии в березняках зоны южной тайги не превышала 5 лет. В то же время разница развития растений на границе ареала в условиях сосняки и березняка орляково-го зоны южной тайги не выявлена, разница по среднему показателю возраста растений вирги-нильного онтогенетического состояния между двумя фитоценозами южной тайги была статистически значима. Это позволяет предположить, что вид в пределах занимаемого ареала приурочен к специфическим местообитаниям, эколого-ценотический условия которых оказывают большее значение на развитие растений и онтогенетическую структуру ЦП.
Таблица 2. Календарный возраст растений виргинильного онтогенетического состояния
|
Показатели Местообитания |
n |
Медиана |
Варьирование признака (мин-макс) |
Доверительный интервал для медианы |
p-level парные сравнения |
|
сосняк разнотравный |
32 |
4 |
2-7 |
4-5 |
1/3=0,000002 2/3=0,03 |
|
березняк орляковый |
29 |
4 |
3-5 |
3-4 |
|
|
березняк грушанковый |
47 |
3 |
2-5 |
2-4 |
В условиях березняка грушанкового увеличение календарного возраста сопровождалось последовательным переходом растений в более взрослое онтогенетическое состояние. Разница между календарным возрастом растений различных онтогенетических состояний статистически значима (табл. 3). В то же время, в условиях сосняка разнотравного, разница между календарным возрастом растений вир-гинильного и средневозрастного онтогенетических состояний выявлена не была, растения виргиниль-ного и средневозрастного генеративного состояния имели один и тот же календарный возраст. Это позволяет предположить, что условия низкой освещенности лимитируют нормальное развитие растений, а увеличение календарного возраста не способствует к переходу растений в иное онтогенетическое состояние.
Таблица 3. Календарный возраст растений различных онтогенетических состояний в березняке-грушанковом
|
Показатели Онтогенетические группы |
Медиана |
Изменчивость признака (мин-макс) |
Доверительный интервал для медианы |
p-level |
|
j |
1 |
1-2 |
1-1 |
(1/2)=0,00014; (2/3)=0,00005; (3/4)=0,00014 |
|
im |
2 |
1-5 |
2-2 |
|
|
v |
3 |
2-5 |
2-4 |
|
|
g2 |
4 |
2-6 |
4-5 |
Выводы:
-
1. Высокая доля растений виргинильного онтогенетического состояния в структуре ЦП может быть обусловлена как интенсивными процессами вегетативного размножения, так и замедленным развитием рамет. Так, в сосняке разнотравном преобладание растений виргинильного онтогенетического состояния связано с замедленным развитием, в березняке орляковом и березняке грушанковом – интенсивными процессами вегетативного размножения, быстрым переходом в виргинильное онтогенетическое состояние.
-
2. При отсутствии стационарных наблюдений за развитием маркированных растений, определение календарного возраста растений различных онтогенетических состояний может способствовать корректной интерпретация онтогенетической структуры ЦП.
-
3. В ЦП P. rotundifolia, существующей на границе ареала, наблюдаются затруднения поддержания численности на основе процессов полового и
- вегетативного размножения, а потому неконтролируемый сбор растительного сырья может привести к исчезновению ЦП.
Список литературы Анализ календарного возраста рамет грушанки круглолистной (Pyrola rotundifolia L.) при интерпретации онтогенетической структуры вида
- Олейникова, Е.М. Стержнекорневые травы юго-востока средней России: автореф. дис… д.б.н. -Воронеж, 2015. 43 с.
- Фардеева, М.Б. Экологические и биоморфологические закономерности пространственно-онтогенетической структуры популяций растений, динамика и мониторинг: автореф. дис… д.б.н. -Казань, 2014. 48 с.
- Работнов, Т.А. Вопросы изучения состава популяций для целей фитоценологии//Проблемы ботаники. 1950. Вып. 1. С. 465-483.
- Завадский, К.М. Вид и видообразование. -Л.: Наука, 1968. 396 с.
- Уранов, А.А. Жизненное состояние вида в растительном сообществе//Бюл. МОИП. Отд. биол. 1960. Т. 65. Вып. 3. С. 77-92.
- Бобров, Ю.А. Грушанковые России. Монография.-Киров: ВятГГУ, 2009. 137 с.
- Животовский, Л.А. Онтогенетические состояния, эффективная плотность и классификация популяций растений//Экология. 2001. № 1. С. 3-7.
- Современные подходы к описанию структуры растения/Под ред. Н.П. Савиных, Ю.А. Боброва. Монография. -Киров: ООО «Лобань», 2008. 355 с.
- Гиляров, А.М. Популяционная экология. Учеб. пособие. -М.: Изд-во МГУ, 1990. 191 с.
- Дубровная, С.А. Жизненный цикл и регенерационные ниши травянистых растений в лесных сообществах//Сибирский лесной журнал. 2016. № 3. С. 24-33.
