Анализ микробных сообществ термальных источников района озера Фумарольное Кальдеры вулкана Узон, Камчатка

Автор: Дворянчикова Е.Н., Кизилова А.К., Кравченко И.К., Гальченко В.Ф.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные ресурсы
Статья в выпуске: 1-6 т.13, 2011 года.
Бесплатный доступ
Впервые проведено комплексное молекулярно-биологическое исследование микробных сообществ 5 термальных источников, расположенных в районе озера Фумарольное кальдеры вулкана Узон, Камчатка. Были использованы методы, основанные на применении ПЦР технологий (ПЦР-детекция, ПЦР-ДГГЭ, клонирование) и гибридизации in situ для анализа как рибосомальных, так и функциональных генов, отвечающих за синтез ключевых ферментов метанотрофии, метаногенеза, автотрофной фиксации СО2, нитрификации. Установлено, что общая численность микроорганизмов в образцах ила составляла от 0,5 до 36,8×106 клеток/мл и метаболически активные эубактерии составляли 22-35%. Методом ПЦР представители Bacteria и также фототрофные прокариоты были обнаружены во всех исследованных источниках, Archаea в четырех, а метанотрофы в трех. Ни в одном из источников не были обнаружены метаногенные и нитрифицирующие археи. Впервые в гидротермах изучено разнообразие метанотрофных сообществ и обнаружено низкое разнообразие, представленное только метанотрофами I типа, наиболее близкими к Methylotermus и Methylobacter. Получено 5 накопительных высокообогащенных монокультур экстремально термофильных метанотрофов, в состав которых входят организмы, значительно отличающиеся от двух известных видов рода Methylotermus.
Микробные сообщества, метанотрофы, накопительные культуры, гибридизация
Короткий адрес: https://sciup.org/148199878
IDR: 148199878 | УДК: 579.64.631.46
Analysis of microbial communities of thermal springs of lake fumarolnoe area, Uzon volcano Caldera, Kamchatka
For the first time complex molecular-biological studies of microbial communities of 5 thermal springs, located around Lake Fumarolnoe, Uzon volcano caldera, Kamchatka, is conducted. The methods based on PCR technologies (PCR-detection, PCR-DGGE, cloning) and fluorescence in situ hybridizations (FISH) for analysis of ribosomal and functional genes, which are responsible for synthesis of key enzymes of methanotrophy, methanogenesis, autotrophic СО2 fixation, nitrification have been used. We found that aggregate number of microorganisms in sludge samples varied from 0.5 to 36.8×106 cells/ml, and metabolically active eubacteria shared 22-35 %. Representatives of Bacteria and phototrophic prokaryotes have been found out by PCR in all thermal springs, Archаea - in four, and methanotrophs - in three. No methanogens and nitrifying archaea were detected in any of the springs. For the first time in hydrotherms methanotrophic communities are studied, and the low diversity presented only methanotrophs of I type, most closely related to Methylothermus and Methylobacter, is revealed. We isolated 5 enrichment monocultures of extremely thermophilic methanotrophs, which structure includes organisms, considerably different from two known species of genera Methylothermus.
Текст научной статьи Анализ микробных сообществ термальных источников района озера Фумарольное Кальдеры вулкана Узон, Камчатка
типами анаэробного дыхания (серо-, нитрат- и железоредукторам, использующим СО), тогда как другие группы микроорганизмов, например, использующие в своем метаболизме метан и/или другие С 1 соединения, исследованы крайне недостаточно. Основные микробиологические исследования выполнены на объектах, расположенных на Восточном термальном поле, а изучение микробных сообществ в термальных источниках района озера Фумарольное практически не проводились. В то же время образование горячих озер на месте кратероподобных воронок является уникальной особенностью Узонских термопроявлений. Крупнейшим среди них является озеро Фумарольное, которое заполняет обширную котловину размером 300 х 600 м с хорошо выраженным береговым обрывом.
Цель исследования: изучение микробного разнообразия в термальных источниках района озера Фумарольное кальдеры вулкана Узон (Камчатка) с помощью методов молекулярной экологии. Особое внимание было уделено мета-нокисляющим организмам, сведения о которых практически отсутствуют.
Материалы и методы.
Отбор образцов и определение физикохимических параметров среды. Образцы цианобактериальных матов и ила были отобраны в ходе экспедиционных исследований в июле 2010 г. Определение физико-химических параметров гидротерм (температуры, pH, Eh) производили с помощью наборов Aquamerk («Merck», Германия). Результаты представлены в табл. 1. Непосредственно на месте отбора 2 см3 образца вносили во флаконы объемом 120 мл с 20 мл среды «П» [3], разбавленной в 5 раз, и инкубировали в атмосфере метан: воздух (1:1) в течение 7 суток в источнике Заварзина (56,7оС). Полученные обогащенные образцы (первичная накопительная культура) были использованы в лаборатории для проведения исследований методами ПЦР и FISH, а также для выделения накопительных культур метано-трофов.
Выделение ДНК и ПЦР-детекция прокариот различных таксономических и функциональных групп. Препарат тотальной ДНК выделяли с помощью коммерческого набора реактивов (Wizard Genomic DNA Purification Kit, «Prome-ga», США) в соответствии с протоколом производителя с минимальными модификациями. Для увеличения выхода микробной ДНК мы проводили обработку лизирующим буфером при повышенной температуре (80оС) и встряхивании. Для амплификации фрагментов рибосомального гена 16S рРНК бактерий использовали систему праймеров 341 F-907 R [4], для архей – 344F-915R [5]. Для детекции прокариот различных функциональных групп (метанотрофы, фотосинтетики, метаногены, нитрификаторы) использовали ПЦР-амплификацию фрагментов генов ключевых ферментов различных процессов метаболизма. Для метанотрофов применяли систему вырожденных праймеров А189F и A682R [6], амплифицирующих фрагмент гена pmoA , кодирующего синтез β-субъединицы метанмонооксигеназы (ММО), ключевого фермента метанокис-ления, а для обнаружения прокариот, обладающих генами автотрофной фиксации углекислоты в цикле Кальвина ( cbbL ) применяли праймерную систему cbbl1F- cbbLR [7].
Присутствие метаногенных архей детектировали с помощью праймерной системы mcrAF /mcrAR [8], позволяющей амплифицировать фрагмент гена mcrA, кодирующего α-субъеди-ницу фермента метил-коэнзим М, а нитрифицирующих архей – праймерной системой Crena-moA23f и CrenamoA616 [9] для детекции фрагмента amoA архей. Оценку разнообразия метано-трофов проводили методом денатурирующего градиентного гель-электрофореза (ДГГЭ) с использованием праймера A189F(GC) с GC-клампом для обеспечения разделения смеси ампликонов pmoA в полиакриламидном геле [10]. Состав реакционной смеси для ПЦР и температурно-временной профиль реакции соответствовали таковым в упомянутых выше работах. Амплификацию проводили на приборе MyCycler («BioRad», США). Анализ продуктов ПЦР проводили в 1,2% агарозном геле, окрашенном бромистым этидием. Выделение и очистку ПЦР фрагментов осуществляли с помощью набора Wizard PCR Preps («Promega», США), согласно рекомендациям производителя.
Оценка бактериального разнообразия методом клонирования фрагмента гена 16S рРНК. Очищенные ПЦР-фрагменты клонировали с помощь pGEM-T easy vector system I (Promega, США) в компетентных клетках E.coli DH10B. Для создания каждой библиотеки клонов случайным образом было отобрано по 50 колоний, проявивших положительную реакцию (белые). С материалом колоний была поставлена ПЦР реакция с универсальными плазмидными праймерами M13F и M13R. Наличие вставки оценивали методом агарозного электрофореза. Генетический материал клонов, содержащих вставки гена 16S рРНК, был секвенирован в сервисной лаборатории с помощью набора реактивов Big Dye Terminator v.3.1 на автоматическом секвенаторе ABI 3730 (Applied Biosystems Inc., США).
Денатурирующий градиентный гель-электрофорез. Разделение смеси ампликонов pmoA проводили с помощью прибора DCode Universal Mutation Detection System («BioRad»,США) при постоянной температуре 60оС, напряжении 200 V и градиенте денатурантов (формамид, мочевина) 35-60% в течение 6 ч. Полученные гели окрашивали раствором этиди-ум бромида и документировали с помощью имидж-системы Gel Doc System («BioRad», США). Характерные видимые полосы (bands), содержащие ДНК, были вырезаны, полученные после элюирования растворы ДНК были очищены и использованы для определения нуклеотидных последовательностей методом секвенирования.
Гибридизация in situ с флуоресцентномечеными олигонуклеотидными зондами (FISH). Фиксацию образцов раствором параформальдегида и гибридизацию препаратов с флуоресцентно-мечеными олигонуклеотидными зондами проводили при температуре 46оС в соответствии с методикой [11]. Для специфической детекции метанотрофов II типа использовали зонд М-450 (30% формамида в гибридизационном буфере), а метанотрофов I типа – смесь зондов М-84 и М705 (20% формамида) [12].Для детекции представителей домена Bacteria использовали смесь универсальных зондов EUB 338mix [13]. Синтез зондов, меченных флуоресцентным красителем Су3, выполнен компанией «Синтол» (Москва, Россия). Общую численность бактерий определяли в препаратах, окрашенных раствором ДНК-специфичного красителя ДАФИ. Количество гибридизованных с зондами клеток подсчитывали с помощью микроскопа AxioImager D1 («Karl Zeiss», Германия) в 50 полях зрения с использованием светофильтров Zeiss 20 для Cy3-меченных зондов и Zeiss 49 для подсчета клеток, окрашенных ДАФИ, с последующим расчетом на 1 мл обогащенного образца.
Выделение и анализ накопительных культур. Накопительные культуры из обогащенных образцов получали путем регулярных пересевов один-два раза в месяц. Инкубацию вели в статических условиях при температуре 60оС. Процесс роста метанотрофных бактерий при последовательных пересевах контролировали с помощью световой микроскопии, FISH и ПЦР детекции метанотрофов. Представленные результаты анализа были получены для накопительных культур, полученных через 3 месяца инкубации (3-4 пассажа).
Анализ нуклеотидных последовательностей. Анализ полученных нуклеотидных последовательностей фрагментов генов проводили с помощью программного пакета BLAST [].
Результаты и обсуждение. Характеристики гидротерм. В июле 2010 г. был проведен отбор образцов ила и цианобактериальных матов из жерла термальных источников Культурный, Терра, Строма, Квадрат и Глаз дракона, расположенных в районе озера Фумарольное кальдеры вулкана Узон. Физико-химические характеристики вод изученных гидротерм представлены в табл. 1. Все исследованные источники были высокотемпературными со слабокислой или близкой к нейтральной реакцией среды.
Таблица 1. Характеристики термальных источников
|
Название источника |
Координаты |
Температура, оС |
pН |
Eh, мВ |
|
|
N |
E |
||||
|
Культурный |
54030. 115` |
1590 59. 279` |
63,0 |
5,2 |
80 |
|
Терра |
54029. 887` |
1590 59. 410` |
65,0 |
6,2 |
73 |
|
Строма |
54029. 854` |
1590 59. 477` |
56,1 |
5,3 |
5 |
|
Квадрат |
54029. 862` |
1590 59. 485` |
64,5 |
6,3 |
- 22 |
|
Глаз Дракона |
54029. 885` |
1590 59. 468` |
59,8 |
6,2 |
н.о. |
Примечание: н.о. – не определяли.
ПЦР-детекция прокариот. На первом этапе исследования была проведена амплификация фрагментов таксономических и функциональных генов прокариот. Примеры детекции целевых организмов продемонстрированы на рис. 1.
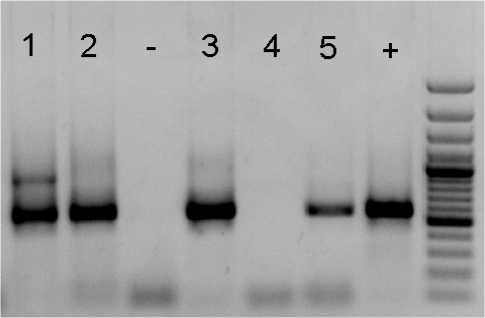
а)

б)
Рис. 1. Визуализация результатов ПЦР с помощью агарозного электрофореза: а – археи, система праймеров 344F-915R; б – метанотро-фы, система праймеров А189F и A682R; 1 – Культурный; 2 – Квадрат; 3 – Терра; 4 –Глаз Дракона; 5 –Строма; «–» отрицательный контроль; «+» положительный контроль, организмы Methanosarcina lacustris (а ), Methylosinus trichosporium (б).
Результаты анализа показали присутствие Bacteria во всех исследованных источниках и Arch-аea в четырех (рис. 2а). Анализ функциональных генов также продемонстрировал различия в составе микробных сообществ. Метанотрофные бактерии были детектированы только в 3 источниках (рис. 1б), а фототрофные – во всех 5 источниках.
Из термальных источников Камчатки с кислой реакцией среды (pH 3,5) были выделены ацидо-термофильные метанокисляющие представители филума Verrucomicrobia [14, 15], в то время как сведения о метанотрофах в высокотемпературных нейтральных источниках Камчатки в литературе отсутствуют. Таким образом, в нашем исследовании впервые было зафиксировано их присутствие в горячих источниках Камчатки.
Распространение фототрофных цианобактерий и анаэробных фотосинтезирующих бактерий в термальных источниках изучено для достаточно большого количества объектов, расположенных в России, Исландии, США и Китае [16], поэтому неудивительным был факт обнаружения организмов, обладающих генами первичной ассимиляции СО 2 во всех исследованных источниках. Для осадков источника Культурный фрагмент гена cbbL был секвенирован, и полученная нуклеотидная последовательность продемонстрировала сходство с депонированной в GenBank последовательностью фототрофной бактерии Chloroflexus . Ни в одном из исследованных источников нам не удалось детектировать присутствие аммонийокисляющих архей, хотя в работе (Zhang et al., 2008) было обнаружено присутствие нитрифицирующих кренархей в высокотемпературных источниках Камчатки [17]. Ни в одном из источников не были обнаружены метаногенные археи. Результаты ПЦР-детекции прокариот в источниках суммированы в табл. 2.
Анализ бактерий в обогащенных образцах методом FISH. Общее количество микроорганизмов, выявленных в обогащенных образцах (окраска ДАФИ), составляло от 5 до 36 х 106 клеток на 1 мл осадка, а количество метаболически активных бактерий (EUB338 mix) варьировало от 2,3 до 21 х 106 клеток на 1 мл осадка (табл. 3). Доля активных бактерий была максимальной (35,6%) в образцах источника Квадрат, а в остальных варьировала от 22,3% до 31,5%.
Таблица 2. Результаты амплификации рибосомальных и функциональных генов бактерий и архей в образцах осадков термальных источников
|
Образование целевого продукта |
|||||||
|
Группа прокариот |
Целевой ген |
Праймерные системы |
Термальные источники |
||||
|
1 |
2 |
3 |
4 |
5 |
|||
|
все Bacteria |
16S рРНК |
341 F -907 R |
+ |
+ |
+ |
+ |
+ |
|
метанотрофные бактерии |
pmoA |
А189F- A682R |
+ |
- |
+ |
+ |
- |
|
фототрофные бактерии |
cbbL |
cbbl1F- cbbLR |
+ |
+ |
+ |
+ |
+ |
|
все Archea |
16S рРНК |
344F-915R |
+ |
+ |
+ |
- |
+ |
|
метаногенные археи |
mcrA |
mcrAF /mcrAR |
- |
- |
- |
- |
- |
|
нитрифицирующие кренархеи |
amoA |
CrenamoA23f -CrenamoA616 |
- |
- |
- |
- |
- |
Примечание: источники: 1 – Культурный, 2 – Терра, 3 – Строма, 4 – Квадрат, 5 – Глаз Дракона; «+» -означает детекцию целевого организма, «-» - не обнаружено.
Анализ обогащенных образцов методом FISH выявил высокие значения численности ме-танотрофов I типа (γ- Proteobacteria ) в источниках Культурный, Строма и Квадрат. Количество клеток здесь достигало 1,5-2,0 х 106 клеток на 1 мл осадка. В источниках Терра и Глаз Дракона количество метанотрофов было значительно ниже
– 105 и 104 клеток на 1 г осадка, соответственно. Доля метанотрофов I типа от метаболически активных эубактерий составляла: Культурный – 4,3%; Терра – 3,2%; Строма – 4,2%; Квадрат – 3,4%; Глаз Дракона – 0,2%. Метанотрофы II группы (зонд М-450) ни в одном из исследованных источников обнаружены не были.
Таблица 3. Численность клеток в обогащенных образцах, определенная методом гибридизации in situ с флуоресцентными олигонуклеотидными зондами и окраской ДАФИ.
|
Источник |
Культурный |
Терра |
Строма |
Квадрат |
Глаз Дракона |
|
окраска / гибридизация |
количество клеток, х106 /мл осадка |
||||
|
ДАФИ |
24±2,2 (68.2%) |
11±3,9 (20,1%) |
33±5,6 (73,5%) |
36±4,5 (61%) |
5±0,68 (68,4%) |
|
EUB338 mix |
9,7±0,39 (27,6%) |
4,2±1,4 (26,8%) |
10±1,6 (22,3%) |
21±3,3(35,6%) |
2,3±0,81 (31,5%) |
|
М-84 + М-705 |
1,5±0,46 (4,3%) |
0,5±0,19 (3,2%) |
1,9±0,61 (4,2%) |
2±0,49 (3,4%) |
0,013±0,002(0,2%) |
Оценка разнообразия бактерий в образце ила источника Культурный методом клонирования 16S рРНК. Клонирование ПЦР-фрагментов позволило оценить разнообразие бактерий в источнике Культурный. Из анализа результатов клонирования (табл. 4) следует, что доминирующим организмом (29 клонов) в источнике является метанотроф, наиболее близкий к штамму НВ [16]. Минорные компоненты (1-5 клонов) представлены термофильными организмами, проявившими высокую степень сходства с бактериями, выделенными из термальных источников Камчатки, Йеллоустонского национального парка, Тибета и геотермальных почв Новой Зеландии.
ПЦР-ДГГЭ анализ фрагмента гена pmoA. Для 4 источников, в которых был амплифициро-ван фрагмент гена pmoA, был проведен ПЦР-ДГГЭ анализ, позволяющий оценить разнообразие метанокисляющих бактерий. В табл. 5 представлены данные по анализу нуклеотидных последовательностей характерных полос. Проведенное исследование показало, что во всех источниках присутствуют метанотрофы, наиболее близкие к представителям рода Methylothermus, но отличающиеся от двух описанных видов (79-91% сходства нуклеотидных последовательностей). В образцах из источников Терра и Глаз Дракона были обнаружены последовательности, наиболее близкие к некультивируемым представителям рода Methylomonas (79-90% сходства). Только в образце из источника Терра обнаружены 2 мета-нотрофных организма, в то время как во всех остальных образцах – по одному. В источниках Культурный, Терра, Квадрат идентифицированы метанотрофы, проявившие высокую степень сходства с термофильным метанотрофом НВ [18]. Для метанотрофа из образца Строма ближайшим родственником был Methylothermus sub-terraneus из горячих подземных водоносных слоев золотодобывающих рудников Японии [19].
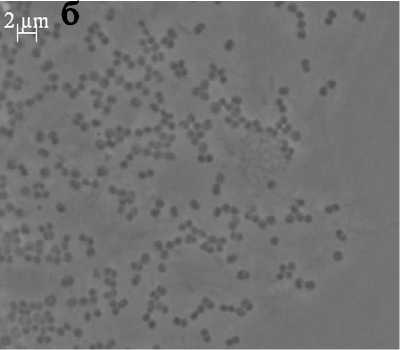
Характеристика накопительных культур термофильных метанотрофов. Из образцов ила источника Культурный, а также источников Глаз дракона и Строма путем последовательных пересевов на селективных средах были выделены 5 накопительных культур метанотрофов, присутствие которых в обогащенных образцах нам не удалось детектировать. Накопительные культуры представляли собой ассоциации метанокисляю-щих бактерий с гетеротрофными спутниками, однако доля метанокисляющих бактерий в них была высокой. Морфология метанотрофных клеток, дающих сигнал с зондом М-84+М-705 (I группа метанотрофов), была сходной для всех накопительных культур. Это были короткие, крупные (около 1 мкм) округлые палочки, которые хорошо идентифицировались методом фазового контрастирования препаратов живых клеток. Электронно-микроскопический анализ интактных культур метанотрофов культуры К12 выявил присутствие полярного жгутика, а анализ срезов обнаружил мембранные структуры, характерные для метанотрофов I типа. Анализ последовательностей pmoA показал, что во всех накопительных культурах присутствуют метанотрофы, наиболее близкие к Methylotermus.
Таблица 4. Разнообразие бактерий в образце ила источника Культурный методом клонирования 16S рРНК
|
Номер образца |
Ближайший представитель |
Процент сходства покрытие/сходство |
|
1 (Культурный, 29 клонов) |
Thermophilic methanotroph HB (U89302) Uncultured bacterium clone SL_5.95 (AY550713) |
98%(99%) 98%(94%) |
|
2 (Камчатка 14, 15 клонов) |
Uncultured Chlorobi bacterium clone VSM5Q1u76 16S ribosomal RNA gene, partial sequence (EU631213) Uncultured green sulfur bacterium clone SM1H02 16S ribosomal RNA gene, partial sequence (AF445702)---- |
95% 94% |
|
3 (Культурный, 5 клонов) |
Thermoactinomyces sacchari (AM161154) Uncultured bacterium clone LaC15L24(EF667582) Propionibacterium acnes (AM161153) |
98% (83%) 73%(89%) 9%(83%) |
|
4 (Культурный, 1 клон) |
Uncultured bacterium clone ZB_P10_C06 (GQ328567) из источника Заварзина в Узоне Uncultured Acidobacteriaceae bacterium (AM935486) |
83% (100%) 83% (95%) |
|
5 (Культурный, 1 клон) |
Pseudomonas putida (AM161157) |
98% (95%) |
|
6 (Культурный, 2 клона) |
Uncultured bacterium clone ZB_P14_B04 16S (GQ328675 из источника Заварзина в Узоне Candidate division OP10 clone OPB80 (AF027089) из горячего источника Йеллоустонского парка |
73%(97%) 73%(96%) |
|
7 (Культурный, 1 клон) |
Uncultured bacterium clone ZB_P13_F12 (GQ328655) из источника Заварзина в Узоне |
73%(85%) |
|
8 (Культурный, 1 клон) |
Uncultured bacterium clone TP146 (EF205576) hot springs in central Tibet Uncultured Acidobacteria (AM749745) from geothermal soils in New Zealand |
78%(97%) 78%(94%) |
Таблица 5. Результаты BLAST анализа нуклеотидных последовательностей характерных ДГГЕ полос, полученных при разделении ПЦР фрагментов гена.
|
Источник |
Ближайший родственник |
Покрытие/сходство |
|
Культурный |
Uncultured bacterium clone SL_5.95 (AY550713) Thermophilic methanotroph HB (U89302) |
99/80 99/79 |
|
Терра (смесь) |
Uncultured bacterium clone SL_5.32 (AY550731) Thermophilic methanotroph HB (U89302) |
100/95 100/95 |
|
Uncultured bacterium (AM910134) Uncultured Methylomonas sp . (AB500822) |
100/80 100/79 |
|
|
Строма |
Methylothermus subterraneus (AB536748) |
100/91 |
|
Квадрат |
Uncultured bacterium clone SL_5.95 (AY550713) Thermophilic methanotroph HB (U89302) |
93/88 93/88 |
|
Глаз Дракона |
Uncultured bacterium (AM910134) Uncultured Methylomonas sp . (AB500822) |
95/90 95/87 |
Выводы: в результате проведенных исследований впервые получены характеристики микробных сообществ гидротерм Камчатки с помощью комплексного молекулярно-биологического подхода, основанного на использовании ПЦР технологий (ПЦР-детекция, ПЦР-ДГГЭ, клонирование) и гибридизации in situ для анализа рибосомальных генов, а также функциональных генов, отвечающих за синтез ключевых ферментов метанотрофии, метаногенеза, автотрофной фиксации СО2, нитрификации). Впервые изучено распространение и разнообразие метанотрофных сообществ в осадках нейтральных высокотемпературных источников. Установлено, что разнообразие термофильных метанотрофов в исследованных источниках ограничивается бактериями, наиболее близкими к Methylotermus и Methylobacter. Получено 5 накопительных высокообогащенных (20-50%) монокультур метанотрофов, в состав которых входят организмы, значительно отличающиеся от двух известных видов рода Methy-lotermus. Культуры являются экстремальными термофилами и их рост осуществляется при значениях температур не ниже 50оС. Дальнейшие исследования предполагают более детальные исследования обнаруженных организмов, выделение чистых культур и описание новых таксонов термофильных метанотрофов.
Работа выполнена при поддержке грантов РФФИ № 08-04-00164-а, 09-04-10113-к и 10-04-10127-к.
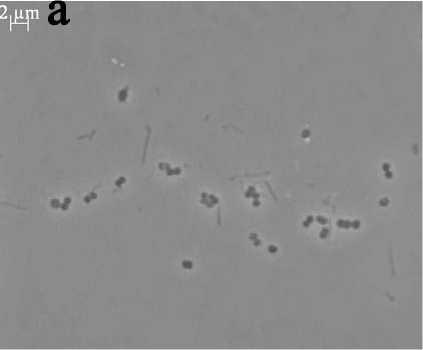
Рис. 2. Морфология клеток накопительных культур метанотрофов: а – К14, источник Культурный; б – К12, источник Строма (фазовый контраст)
Список литературы Анализ микробных сообществ термальных источников района озера Фумарольное Кальдеры вулкана Узон, Камчатка
- Brock, T.D. Microbiological studies of thermal habitats of the central volcanic region, North Island, New Zealand./T.D. Brock, M.L. Brock//N. Z. J. Marine and Freshwater Res. 1971. V.5. P. 233-258.
- Труды института микробиологии имени С.Н. Виноградского: Вып.16: Термофильные микроорганизмы//Ин-т микробиологии им. С.Н. Виноградского РАН. -М.: МАКС Пресс, 2011. 364 с.
- Гальченко, В.Ф. Метанотрофные бактерии. -М.: ГЕОС, 2001. 500 с.
- Muyzer, G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA/G. Muyzer, E.C. de Waal, A.G. Uitterlinde//Appl. Environ. Microbiol. 1993. V. 59. P. 695-700.
- Casamayor, E.O. Identification of and spatiotemporal differences between microbial assemblages from two-neighboring sulfurous lakes: comparison by microscopy and denaturing gradient gel electrophoresis/E.O. Casamayor, H. Schafer, L. Baneras//Appl. Environ. Microbiol. 2000. 66. С. 499-508.
- Holmes, A.J. Evidence that particulate methane monooxigenase and ammonia monooxigenase may be evolutionarily related/A.J. Holmes, A. Costello, M.E. Lidstrom, G.C. Murrell/FEMS Microbiol. Lett. 1995. V. 132. P. 203-208.
- Спиридонова, Е.М. Система олигонуклеотидных праймеров для амплификации генов рибулозо-1,5-бисфосфаткарбоксилазы/оксигеназы у бактерий различных таксономических групп/Е.М. Спиридонова, И.А. Берг, Т.В. Колганова и др.// Микробиология. 2004. № 2. С. 377-380.
- Luton, P.E. The mcrA gene as an alternative to 16S rRNA in the phylogenetic analysis of methanogen populations in landfill/P.E. Luton, J.M. Wayne, R.J. Sharp, P.W. Riley//Microbiology. 2002. V. 148. P. 3521-3530.
- Tourna, M. Growth, Activity and Temperature Responses of Ammonia-Oxidising Archaea and Bacteria in Soil Microcosms/M. Tourna, T.E. Freitag, G.W. Nicol, J.I. Prosser//Environ. Microbiol. 2008. Vol. 10. P. 1357-1364.
- McDonald, I.R. Molecular ecology techniques for the study of aerobic methanotrophs/I.R. McDonald, L. Bodrossy, Y. Chen, C.J. Murrell//Appl. Environ. Microbiol. 2008. V. 74. P. 1305-1315.
- Stahl, D.A. Nucleic Acid Techniques in Bacterial Systematics/D.A. Stahl, R. Amann//Development and Application of Nucleic Acid Probes. Eds. E. Stakebrandt and M. Goodfellow Chichester: Wiley,1991. P. 205-248.
- Eller, G. Group-specific 16S rRNA targeted probes for the detection of type I and type II methanotrophs by fluorescence in situ hybridization/G. Eller, S. Stubner, P. Frenzel//FEMS Microbiol. Lett. 2001. V. 198. P. 31-37.
- Diams, H. The domain-specific probe EUB338 is insufficient for the detection of all Bacteria: Development of a more comprehensive probe set/H. Diams, A. Bruhl, R. Amann et al./Syst. Appl. Microbiol. 1999. V. 22. P. 434-444.
- Pol, A. Op den Camp. Methanotrophy below pH 1 by a new Verrucomicrobia species/A. Pol, K. Heijmans, H.R. Harhangi//Nature. 2007. V. 450. P. 874-878.
- Bodrossy, L. A novel methane-oxidising -Proteobacterium/L. Bodrossy, K. Kovács, I.R. McDonald, K. Murrell//FEMS Microbiol. Lett. 1999. V. 170. P. 335-341.
- Madigan, M.T. Anoxygenic phototrophic bacteria from extreme environments//In: Discoveries in Photosynthesis. Advances in Photosynthesis and Respiration, 2005. Vol. 20, XII., 969-983, DOI: DOI: 10.1007/1-4020-3324
- Chuanlun, L. Zhang. Global Occurrence of Archaeal amoA Genes in Terrestrial Hot Springs/Chuanlun L. Zhang, Qi Ye, Zhiyong Huang et al.//Appl. Environ. Microbiol. 2008. V.74. P. 6417-6426.
- Bodrossy, L. A novel methane-oxidising -Proteobacterium/L. Bodrossy, K. Kovács, I.R. McDonald, K. Murrell//FEMS Microbiol. Lett. 1999. V. 170. P. 335-341.
- Hirayama, H. Methylothermus subterraneus sp.nov., a moderately thermophilic methanotrophic bacterium from a terrestrial subsurface hot aquifer in Japan/H. Hirayama, Y. Suzuki, M. Abe et al.//Int. J. Syst. Evol. Microbiol. 2010. DOI 10.1099/ijs.0.028092-0.
