Анализ распространения крымского скорпиона (Arachnida, Scorpiones, Euscorpiidae) c замечаниями о его биогеографическом статусе на Крымском полуострове
 c замечаниями о его биогеографическом статусе на Крымском полуострове")
Автор: Кукушкин О.В.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Научные сообщения
Статья в выпуске: 3 т.22, 2013 года.
Бесплатный доступ
Анализ распространения и биотопических предпочтений крымского скорпиона ( Euscorpius tauricus ) в Крыму выявляет его абсолютную синантропность, что позволяет высказать предположение о случайной интродукции этого термофильного и относительно ксерофильного вида в исторический период из Южно-Эгейского региона.
Крым, распространение, биогеографический статус, синантропия, интродукция
Короткий адрес: https://sciup.org/148314898
IDR: 148314898 | УДК: 595.46:591.9+591.152
Текст краткого сообщения Анализ распространения крымского скорпиона (Arachnida, Scorpiones, Euscorpiidae) c замечаниями о его биогеографическом статусе на Крымском полуострове
pers. comm.; V.Ya. Fet, in progress). На основании этих данных было высказано предположение о его вероятной интродукции в Таврике в течение исторического периода (Кукушкин, Ковблюк, 2011; Кукушкин, 2012). Однако, вопрос о биогеографическом статусе E. tauricus в Крыму (реликт или интродуцент) до настоящего времени остается дискуссионным. Как узкоареальный эндемик крымский скорпион внесен в национальную Красную книгу в категории III – «редкий» вид (Євтушенко, 2009). Несомненно, это оригинальное и совершенно неопасное для человека паукообразное является украшением природы Крыма, имеет высокую научную ценность и – независимо от своего биогеографического статуса – нуждается во всемерной охране, однако изучение его распространения и экологии имеет не только прикладное (природоохранное), но и большое теоретическое значение в свете решения вопроса о генезисе фауны Крымского полуострова.
Методы исследования
Поиск скорпионов осуществлялся во все сезоны в период с 1996 по 2012 г. включительно на всем протяжении южного макросклона Крымских гор, в теплых горных местностях северного макросклона, а также в приморских областях Керченского полуострова (в ночное время с использованием ручного фонаря). Способность кутикулы многих скорпионов к флуоресценции в ультрафиолетовом освещении (Frost et al., 2001), значительно облегчающая их поиски в темное время суток, была отмечена нами и у E. tauricus (Кукушкин, 2012), однако в процессе полевых исследований не использовалась. Учитывались также сведения, полученные от вызывающих доверие респондентов – как правило, опытных зоологов.
Для изучения факторов, определяющих границы ареала E. tauricus на Крымском полуострове, использовались современные ГИС-технологии. При определении десятичных координат локалитетов задействовавали геоинформационный ресурс сайта Для анализа вклада переменных биоклиматических параметров, влияющих на распространение вида, применили компьютерную программу MAXENT 3.3.3e (, позволяющую, в частности, определить степень влияния (в процентах) 19 переменных биоклиматических параметров среды, а также высоты местности н.у.м. на область распространения изучаемого таксона (Phillips, Dudik, 2008). Для моделирования потенциального ареала – по 38 точкам находок вида в Крыму (табл. 1) – была использована климатическая база WorldClim ( (минимальное разрешение – 30 arc-seconds или ~ 1 км на пиксель).
Результаты и обсуждение
Современный уровень знаний не позволяет ответить на вопрос о биогеографическом статусе крымского скорпиона в Украине однозначно. С одной стороны, как говорилось выше, методами молекулярной филогении выявлено близкое родство скорпионов Крыма и ряда островов Эгейского моря (Южные Киклады), причем максимальное сходство отмечено с популяцией острова Парос (Brewer et al., 2005), население которого поддерживало в течение исторического периода тесные политико-экономические связи с античными государствами Таврики (см.: Зубарь, 1993; Виноградов, Золотарев, 1998). Не вдаваясь здесь в историко-археологические подробности, скажем, что случайная интродукция скорпиона в Крыму выходцами с Южных Киклад, принимавшими участие в основании Херсонеса Таврического, либо более поздний его завоз из торговых центров Малой Азии (см.: Кадеев; 1970; Лесная, 2012), представляется событием достаточно реалистичным. Широко известно, что некоторые виды рода Euscorpius Thorell, 1876 – прежде всего относительно крупный и индифферентный к фактору влажности E. italicus (Herbst, 1890) – имеют выраженную тенденцию к антропохории, что позволило им широко расселилиться в исторический период не только в Средиземноморье и/ или Причерноморье, но даже далеко за пределами Европейского континента (Crucitti et al., 1998; Fet, Braunwalder, 2000; Fet et al., 2001; Fet, Kovařík, 2003; Fet et al., 2005). В Крыму известны виды членистоногих, явно непреднамеренно интродуцированные в исторический период скорее всего с Эгейского побережья Малой Азии: например, крупная двупароногая многоножка Eurygyrus ochraceus C.L. Koch, 1847 (Callipodida, Schizopetalidae), обнаруженная мною в конце 1980-х гг. в двух пунктах Гераклейского полуострова – в том числе поблизости от одной из крупнейших гаваней античного Херсонеса (Enghoff, 2006; Golovatch, 2007; Stoev et al., 2010).
С другой стороны, на сегодняшний день нельзя исключать относительно недавнюю (в эоплейстоцене или даже плейстоцене) колонизацию скорпионами Крыма непосредственно из Анатолии, в период одной из значительных регрессий Черноморского бассейна, к чему имеются определенные зоогеографические (Lukhtanov, Budashkin, 2007; Мирошниченко, 2011) и палеогеографические (Андреев, 2010) предпосылки. Данные по молекулярной филогеографии некоторых видов Euscorpius , обладающих фрагментированным ареалом в Западной Европе, Северной Африке и на островах Средиземного моря, говорят о том, что в некоторых случаях возраст дизъюнкций может быть весьма древним (Graham et al., 2012). Анатолийские представители семейства Euscorpiidae до сих пор изучены крайне недостаточно (см.: Fet, Braunwalder, 2000; Koç, Yağmur, 2007; Tropea et al., 2012), и вероятность находок E. tauricus при дальнейших иследованиях на западном и северо-западном побережье Турции расценивается специалистами как весьма высокая (В.Я. Фет, pers. comm.).
На хорошо изученном скорпиологами побережье Болгарии известна единственная находка вида, принадлежащего к роду Euscorpius (к еще неописанному виду комплекса «E. carpathicus») (Fet, Soleglad, 2007), поэтому возможность проникновения скорпиона в Крым с восточных Балкан нами не рассматривается. Отсутствие в Крыму E. (Polytrichobothrius) italicus и E. (Alpiscorpius) mingrelicus (Kessler, 1874) широко распространенных на Западном Кавказе и черноморском побережье Анатолии (Fet, Braunwalder, 2000), и обитание здесь локально распространенной южноэгейской формы не является аргументом в пользу низкой вероятности ее интродукции здесь, так как значительная роль в расселении антропохорных видов животных принадлежит их экологическим преференциям и случайным факторам, а не расстоянию и даже не интенсивности торговых связей (см.: Дузь и др., 2012; Матвеев и др., 2013). Поэтому кратчайший («внутричерноморский») путь расселения не является единственным. В первом приближении, наиболее вероятным районом, исходным для колонизации скорпионом Крыма, представляется западное побережье Анатолии с прилежащими континентальными островами (полуостров Чешме, остров Xиос?) либо некоторые острова архипелага Южные Киклады (Парос или Делос, лежащий всего в 25 км севернее предыдущего, более крупного острова). Однако недостаточная изученность филогенетической структуры рода Euscorpius в целом и комплекса форм «E. tauricus», в частности, на сегодняшний день не позволяет прийти к однозначному заключению о генезисе крымской популяции E. tauricus.
В связи с вышесказанным особый интерес приобретают сведения об особенностях распространения и биотопического распределения E. tauricus в Крыму. Все известные на сегодняшний день находки вида на Крымском полуострове приведены в табл. 1.
Таблица 1 (начало)
Распространение E. tauricus на Крымском полуострове
|
N п/п |
Локалитет; административная территория |
Географические координаты (десятичные) |
Источник (-и) сведений |
|
|
Широта (N) |
Долгота (E) |
|||
|
1 |
г. Евпатория; Ев |
45.19625216954871 |
33.37921142578125 |
Бялыницкий-Бируля (1917); Фет (1989) |
|
2 |
г. Севастополь; Св |
44.60757930079818 |
33.51722717285156 |
Бялыницкий-Бируля (1917); Фет (1989) |
|
3 |
* черта г. Севастополь, Херсонес; Св |
44.60967213376329 |
33.49448204040527 |
Кукушкин (2004) |
|
4 |
г. Севастополь, ул. Батумская; Св |
44.61839209584067 |
33.525375723838806 |
О.В. Кукушкин, неопубл. данные |
|
5 |
* г. Севастополь, ИнБЮМ; Св |
44.5995891793583 |
33.51808547973633 |
С.E. Аносов, pers. comm.; Кукушкин, Ковблюк (2011) |
|
6 |
* г. Севастополь, Корабельная сторона, Аполлонова бухта; Св |
44.61577741590874 |
33.54300186038013 |
Ю.А. Красиленко, pers. comm. |
|
7 |
* г. Инкерман, крепость Каламита; Св |
44.60280904022024 |
33.60966145992279 |
Кесслер (1874); Фет (1989); Кукушкин (2004) |
|
8 |
г. Балакалава, ул. Калича; Св |
44.49730121746426 |
33.59990358352661 |
Бялыницкий-Бируля (1917); Фет (1989); О.В. Кукушкин, неопубл. данные |
|
9 |
с. Резервное; Св |
44.469362167379415 |
33.67464065551758 |
Кукушкин (2004) |
|
10 |
мыс Сарыч, маяк; Св |
44.388530946568245 |
33.73985454440123 |
И.С. Турбанов, pers. comm. |
|
10 |
мыс Сарыч, маяк; Св |
44.388530946568245 |
33.73985454440123 |
И.С. Турбанов, pers. comm. |
|
11 |
г. Бахчисарай; Бх |
44.7504211508603 |
33.874197006225586 |
Фет (1989); В.В. Савчук, pers. comm. |
|
12 |
г. Симферополь; Сф |
44.96358334269838 |
34.08388137817383 |
Бялыницкий-Бируля (1917); Фет (1989); О.В. Кукушкин, неопубл. данные |
|
13 |
пгт Форос, парк; Ял |
44.38992697266835 |
33.78450393676758 |
Г.А. Прокопов, pers. comm. |
|
14 |
пгт Симеиз; Ял |
44.40435407378617 |
34.0081787109375 |
Бялыницкий-Бируля (1917); Фет (1989) |
|
15 |
* пгт Алупка; Ял |
44.417444240638126 |
34.044785499572754 |
Бялыницкий-Бируля (1917); Фет (1989) |
|
16 |
пгт Гаспра; Ял |
44.43739574189946 |
34.10937309265137 |
Фет (1989) |
|
17 |
** пгт Массандра; Ял |
44.51297551471756 |
34.18283376842737 |
Н.М. Ковблюк, pers. comm. |
|
18 |
пгт Ореанда; Ял |
44.45878010882453 |
34.1370964050293 |
Бялыницкий-Бируля (1917); Фет (1989) |
|
19 |
г. Ялта |
44.49532678709199 |
34.167115688323975 |
Бялыницкий-Бируля (1917); Фет (1989) |
|
20 |
* Никитский ботанический сад; Ял |
44.511632965369735 |
34.23411726951599 |
Бялыницкий-Бируля (1917); Фет (1989, 2004) |
|
21 |
* пгт Никита; Ял |
44.51699594354417 |
34.23367738723755 |
О.В. Кукушкин, неопубл. данные |
|
22 |
пгт Гурзуф; Ял |
44.54569975608054 |
34.29567128419876 |
Бялыницкий-Бируля (1917) |
|
23 |
пгт Артек; Ял |
44.55708405292247 |
34.31532382965088 |
Фет (1989) |
|
24 |
окр. пгт Артек; Ял |
44.56764827811433 |
34.31756615638733 |
A.E. Красников, pers. comm. |
|
25 |
окр. пгт Партенит, г. Аюдаг, поляна Ай-Констант; Ал |
44.566402380282255 |
34.345917105674744 |
Д.Б. Старцев, pers. comm. |
|
26 |
пгт Партенит; Ал |
44.5765369464722 |
34.34627652168274 |
Фет (1989) |
|
27 |
окр. пгт Малый Маяк, Кучук-Ламбатский «каменный хаос»; Ал |
44.609779064809615 |
34.36446189880371 |
Д.Б. Старцев, pers. comm. |
|
28 |
г. Алушта; Ал |
44.66908016070617 |
34.411303997039795 |
Фет (1989) |
|
29 |
с. Веселое (=Кутлак); Сд |
44.85725299042211 |
34.8796820640564 |
М.Н. Косарева, pers. comm. |
|
30 |
пгт Новый Свет; Сд |
44.82662206675736 |
34.911691546440124 |
О.В. Кукушкин, неопубл. данные |
|
31 |
Судакская долина; Сд |
44.84114269070498 |
34.97325897216797 |
Бялыницкий-Бируля (1917); Фет (1989) |
|
32 |
окр. г. Судак, г. Палвани-Оба; Сд |
44.83949185310403 |
34.95270252227783 |
О.В. Кукушкин, неопубл. данные |
|
33 |
* черта г. Судак, г. Кыз-Кулле, генуэзская крепость; Сд |
44.84272501598849 |
34.95744466781616 |
Rathke (1837); Бялыницкий-Бируля (1912); Кукушкин, Ковблюк (2011) |
|
34 |
Карадагский заповедник, Биостанция; Фд |
44.91221312808351 |
35.200817584991455 |
Ю.И. Будашкин, pers. comm.; Кукушкин, Ковблюк (2011) |
|
35 |
* г. Феодосия, ул. Десантников, бульвар В. Коробкова и подвальные помещения городской поликлиники |
45.02044019475835 |
35.38762807846069 |
О.Г. Розенберг, В.В. Савчук, pers. comm.; О.В. Кукушкин, неопубл. данные |
|
36 |
черта г. Феодосия, урочище Старый Карантин, генуэзская крепость; Фд |
45.02233988655459 |
35.40672540664673 |
О. В. Кукушкин, неопубл. данные |
|
37 |
окр. г. Феодосия, мыс Ильи; Фд |
45.015180639361915 |
35.408549308776855 |
Е.В. Гладилина, pers. comm. |
|
38 |
г. Керчь; Кр |
45.34186021140748 |
36.46812915802002 |
Фет (1989) |
Обозначения: Ев - Евпаторийский горсовет; Св - Севастопольский горосовет; Бх -Бахчисарайский район; Сф – Симферопольский район; Ял – Ялтинский горсовет; Ал – Алуштинский горсовет; Сд – Судакский район; Фд – Феодосийский горсовет; Кр – Керченский горсовет; значком « * » отмечены локалитеты, где вид является, по нашим данным, многочисленным или вполне обычным (из прочих пунктов известны немногочисленные, как правило, единичные, находки); « ** » - локалитет № 17, где единственный скорпион был отмечен на скале в парке, рассматривался нами в числе синантропных местообитаний.
Из вышеприведенных данных следует, что ареал E. tauricus в Крыму занимает все южное побережье Горного Крыма и выдающийся в море Гераклейский полуостров, по большей части относящийся к юго-западному предгорью. По два пункта обитания вида выявлено во внутренних районах Крыма (в западном предгорье) и на равнинном побережье.
Подавляющее большинство (92,1%; n=38) пунктов находок крымского скорпиона приурочены к синантропным местообитаниям: старым жилым строениям (послевоенный одноэтажный частный сектор и монументальные здания дореволюционной постройки), дорожным крепидам и стенам из дикого камня, античным и средневековым руинам. Количество особей скорпионов, встреченных за годы исследований в природных биотопах, было ничтожным – менее 1% от общего количества регистраций вида (свыше 1000). При интенсивных многолетних полевых исследованиях (моих и арахнологов Н.М. Ковблюка и А.А. Надольного), охвативших всю южную часть Горного Крыма, скорпион за пределами населенных пунктов не отмечался ни разу. Может считаться достоверно установленным его отсутствие в семиаридных приморских редколесьях, мезофильных дубово-грабинниковых шибляках и еще более влажных ясенево-кленовых и грабово-буковых лесах верхнего пояса южного макросклона. Между тем, в Западной Европе и Европейском Средиземноморье распространение скорпионов рода Euscorpius синантропной зоной далеко не ограничивается, и они вполне обычны в природных биотопах, аналоги которых широко представлены в Крыму (Colombo, 2006, 2009). Встречи крымского скорпиона в природных биотопах (ассоциации Junipereta excelsae, Querceta pubescentis и Pistacieta muticae) единичны (табл. 1), и он никогда не отмечался на удалении свыше нескольких (двух – трех) десятков метров от древних или современных руин, что заставляет предполагать переход синантропных популяций к обитанию в естественных ландшафтах по мере разрушения построек. Так, пос. Малый Маяк (=Биюк-Ламбат) расположен близ места локализации древнегреческого порта Лампада, а на г. Аюдаг сохранились остатки одного из наиболее значительных в Крыму комплексов средневековых укрепленных поселений (Фирсов, 1990). У восточного подножья г. Палвани-Оба, где скорпион изредка встречается в скалистых редколесьях Juniperus excelsa , была расположена древнейшая, позднеантичная, часть г. Судак, а в средние века – порт; укрепления, ныне совершенно разрушенные, располагались и на самой горе.
Описанные особенности распространения E. tauricus проступают особенно четко при сравнении с обыкновенной сольпугой Galeodes araneoides (Pallas, 1772) – аборигеном Крымского полуострова и ксерофилом, также тяготеющим к субсредиземноморским ландшафтам нижнего приморского пояса. Если большинство особей G. araneoides встречено в природных биотопах либо в мелких населенных пунктах, «вписанных» в естественные ландшафты, а пункты находок этого паукообразного ясно агрегируются в трех районах крымского побережья, характеризующихся наиболее аридным и жарким климатом (Кукушкин, 2013, in press.), то распространение E. tauricus на побережье Горного Крыма имеет несколько иной характер, и его находки тяготеют к населенным пунктам c давней историей – прежде всего к портовым центрам.
Следовательно, E. tauricus в Крыму может считаться видом полностью синантропным – в отличие, например, от крымского геккона, Mediodactylus kotschyi danilewskii (Strauch, 1887), для которого ранее также постулировалась абсолютная синантропность, а впоследствии (в 1990-2010-е гг.) были выявлены его многочисленные экзоантропные популяции (см.: Кукушкин, 2009). Об антропохории крымского скорпиона свидетельствует его высокая численность в древних портовых центрах Южной Таврики и его находки в городах далеко за пределами побережья, где он способен образовывать устойчивые популяции – обычно неподалеку от транспортных узлов. Например, на протяжении XX столетия (в период с 1907 по 1997 гг.) скорпион неоднократно отмечался в г. Симферополь, лежащем в северных предгорьях на границе с равниной: винных погребах (Бялыницкий-Бируля, 1917), в частном секторе близ железнодорожного вокзала и в общежитии студентов-биологов (наши данные). На постоянное обитание скорпиона в г. Бахчисарай, отмечавшегося там в конце прошлого столетия К.А. Ефетовым (Фет, 1989), указывает его документированная фотографией находка во второй половине 2000-х гг. (В.В. Савчук, pers. comm.).
В то же время даже в мягких условия крымского побережья однократный завоз скорпионов не всегда приводит к формированию устойчивых популяций. Так, две находки E. tauricus были сделаны Ю.И. Будашкиным в начале – середине 1980-х гг. в поселке Карадагского заповедника [тогда – Карадагского отделения Института Биологии Южных Морей (ИнБЮМ)]. Поскольку скорпион при длительных стационарных исследованиях на Карадаге впоследствии никем не отмечался, логично предположить, что, по всей видимости, имел место его случайный завоз из находящегося в г. Севастополь учреждения-куратора (вероятнее всего, с лабораторной посудой). Еще в начале XXI в. скорпион был вполне обычен в полуподвальных помещениях старого аквариума Севастопольской биологической станции (здание ИнБЮМ в центре города).
Анализ биотопического распределения позволяет сделать вывод, что, во всяком случае, в современных климатических условиях E. tauricus находит свой экологический оптимум в жилье человека и руинах – в биотопах, характеризующихся наиболее стабильным (инерционным) микроклиматом (рис. 1).
Этот же вывод подтверждается результатами анализа влияния биоклиматических параметров на область распространения вида при помощи MAXENT. Наиболее значимым оказывается влияние таких факторов как: размах годовой температуры; сезонность осадков и их среднегодовая сумма; средняя температура наиболее влажного сезона, в Южном Крыму приходящегося на холодное время года (табл. 2). Совокупный вклад перечисленных факторов составляет 90%. Теоретически, именно такого распределения вкладов и следует ожидать при анализе распространения вида восточносредиземноморского происхождения. Заметим, что использование данного программного обеспечения и климатической базы WorldClim для изучения распространения синантропного вида, по нашему мнению, является вполне корректным, так как его местообитания в подавляющем большинстве случаев подвержены влиянию всех климатических факторов (руины под открытым небом, груды бута, заборы из дикого камня и т. п.).

А Б

В
Рис. 1. Типы биотопов
E. tauricus в Крыму: А – юго-восточный район Херсонесского городища; Б – черта г. Балаклава;
В – зарастающие руины поселения на г. Аюдаг
Этот же вывод подтверждается результатами анализа влияния биоклиматических параметров на область распространения вида при помощи MAXENT. Наиболее значимым оказывается влияние таких факторов как: размах годовой температуры; сезонность осадков и их среднегодовая сумма; средняя температура наиболее влажного сезона, в Южном Крыму приходящегося на холодное время года (табл. 2). Совокупный вклад перечисленных факторов составляет 90%. Теоретически, именно такого распределения вкладов и следует ожидать при анализе распространения вида восточносредиземноморского происхождения. Заметим, что использование данного программного обеспечения и климатической базы WorldClim для изучения распространения синантропного вида, по нашему мнению, является вполне корректным, так как его местообитания в подавляющем большинстве случаев подвержены влиянию всех климатических факторов (руины под открытым небом, груды бута, заборы из дикого камня и т. п.).
Таблица 2
Вклад биоклиматических параметров в формирование области распространения E. tauricus в Крыму
|
Биоклиматический параметр |
Принятая аббревиатура |
Единицы измерения |
Вклад (%) |
|
Годовой размах температур |
BIO 7 |
оС [х 10] (BIO 5 – BIO 6) |
43,5 |
|
Сезонность осадков |
BIO 15 |
CV, % |
29,9 |
|
Среднегодовые осадки |
BIO 12 |
мм |
9,1 |
|
Средняя температура наиболее влажного сезона |
BIO 8 |
оС [х 10] |
7,5 |
|
Высота местности |
alt |
м н.у.м. |
3,7 |
|
Минимальная температура наиболее холодного месяца |
BIO 6 |
оС [х 10] |
3,3 |
|
Температурная сезонность |
BIO 4 |
SD [х 100] |
1,9 |
|
Средняя температура наиболее сухого сезона |
BIO 9 |
оС [х 10] |
1,0 |
|
Осадки наиболее сухого месяца |
BIO 14 |
мм |
0,1 |
E. tauricus весьма термофилен и выше 300 м н.у.м. не регистрировался (наиболее высоко расположенный пункт находок – с. Резервное (=Кучук-Мускомия), лежащее в Варнутской долине над мысом Айя, куда скорпион, вероятно, был занесен с Южного берега Крыма); примерно до таких же отметок высот (270-280 м н.у.м.) скорпион прослежен и в окрестностях г. Ялта (участок Массандра – Никита). В целом ареал E. tauricus в Крыму очерчивается областью со среднегодовой температурой около 10-14оС (обычно выше 11оС), средней температурой самого холодного месяца от –0,5 до почти +5оС (как правило, не ниже +1оС) (Атлас, 2003). Значительного снижения численности скорпионов после экстремально суровых зим, когда температура воздуха в местах обитания вида на крайнем западе и востоке побережья Горного Крыма падала до –18, –27оС, а среднемесячные температуры января и февраля были отрицательными (например, зима 2005-2006 гг.), нами не отмечалось. Однако, на наш взгляд, это характеризует не столько пределы термотолерантности вида, сколько высокую надежность его зимовочных убежищ. Описаны случаи натурализации наиболее термофильных видов рода в районах с прохладным климатом севернее 50-й параллели. Так, благополучная популяция южноевропейско-североафриканского E. ( Tetratrichobothrius ) flavicaudis (DeGeer, 1778) существует в доках острова Шеппи (Кент, Англия) свыше 140 лет (Benton, 1993). Длительное время известна также колония этого вида на железнодорожной станции в Эссексе (Eversham, Arnold, 1992).
В сравнении с западноевропейскими и анатолийско-кавказскими представителями рода (Бялыницкий-Бируля, 1917; Fet, Kuntner, Sket, 2001; Vignoli et al., 2005; Colombo, 2006, 2009; Tropea et al., 2012) крымский скорпион, населяющий преимущественно засушливые приморские местности с годовой суммой осадков от 300 до 700 мм, является, безусловно, ксерофильным видом. Годовое количество осадков в местах его обитания приближаются к таковому в Северной Африке (Graham et al., 2012) и бассейне Эгейского моря. Однако он довольно чувствителен к фактору влажности и повсеместно предпочитает селиться на изобилующих глубокими и узкими щелями массивных стенах (в Херсонесе, например – почти четырехметровой толщины) и в мощных нагромождениях бутового камня неподалеку от руин.
Об отсутствии подходящих условий для обитания крымского скорпиона в природе, даже на Южном берегу Крыма, по-видимому, свидетельствует и тот факт, что он практически не колонизует примыкающие к руинам участки природных биотопов, несмотря на то, что плотность его синантропных популяций может достигать весьма высоких значений – десятков особей на 100 м2 (Кукушкин, 2004). Описание сходных случаев, когда интродуцент является вполне успешным на ограниченной территории, за пределы которой по малопонятным причинам не выходит, содержатся в работе B. Eversham и H. Arnold (1992, P. 56-57). Переход скорпионов к экзоантропному образу жизни происходит при разрушении и последующем зарастании руин, их «поглощении» природной растительностью. Таким образом, складывается впечатление, что E. tauricus почти не выходит за пределы своей изначальной области распространения, ограниченной в прошлом различного назначения постройками. Об этом же свидетельствуют данные по распространению крымского скорпиона, относящиеся к XIX – началу XX вв. (Бялыницкий-Бируля, 1917) – к периоду, когда Южный берег Крыма был застроен в значительно меньшей степени, чем в настоящее время.
Избирательности субстрата, описанной для нескольких видов скорпионов, обитающих в Риме и его пригородах (Crucitti et al., 1998), нами не отмечено. E. tauricus населяет строения и руины из известняка, габбро-диабаза, песчаника, ракушечника, бетона, самана. Более высокие значения плотности его популяций, отмечавшиеся нами на строениях из палеоценового (нуммулитового) и сарматского известняков, могут быть объяснены, исходя из легкости обработки камня, в результате чего обеспечивается лучшая подгонка блоков, и – как следствие – создаются оптимальные для скорпиона глубокие и узкие убежища. Не последняя роль принадлежит также большой популярности этого податливого и «теплого» строительного материала в древности и его широкому распространению на Гераклейском полуострове и его ближней округе – в хоре античного Херсонеса, с возникновением которого мы связываем колонизацию скорпионом Таврики.
Потенциальное распространение крымского скорпиона на Крымском полуострове, по результатам анализа программы MAXENT, представлено на рис. 2. Из вышеприведенного видно, что существует высокая вероятность его обнаружения в западной половине Юго-Восточного побережья Крыма между г. Судак и г. Алушта, в пониженной западной части Главной горной гряды (прежде всего в Байдарской долине) и в юго-западном предгорье – преимущественно в границах территорий Севастопольского горосвета и Бахчисарайского района. Если предполагать выявление неизвестных экзоантропных популяций E. tauricus скорее всего не приходится, то благополучные синантропные группировки могут быть обнаружены и в районах с субоптимальными для вида климатическими параметрами – очевидно, что синантропия существенно расширяет область потенциального распространения вида, затрудняя в то же время его выявление. К таким районам (темно-серый цвет на карте) относятся: крайний восток Главной гряды и восточное предгорье, крайний юг Керченского Причерноморья (мыс Чауда, замыкающий Феодосийский залив с востока) и Западное побережье с прилежащей частью равнины. Действительно, лишь недавно E. tauricus был найден в черте г. Феодосия. Многочисленная популяция обнаружена в начале второй декады августа 2010 г. 23.09.2010 в данном пункте, расположенном в западной, припортовой, части города (табл. 1), были предприняты специальные поиски вида. В течение получаса (между 1730 и 1800) на площади около 5 м2 было добыто три особи E. tauricus (рис. 3); еще две ювенильных особи наблюдались. Биотоп скорпиона в Феодосии типичен для Крыма в целом: невысокое нагромождение строительного мусора и блоков известняка у экспонированной на северо-восток стены высокой каменной постройки, воздвигнутой в XIX в. или ранее. Неподалеку имеются и иные фрагменты старой кладки, поросшие папоротниками Ceterach officinarum и Asplenium viridis, где, по сведениям местного населения, также встречаются скорпионы. Подходящие для обитания скорпионов руины выявлены также поблизости – на улицах Циолковского и Греческой.
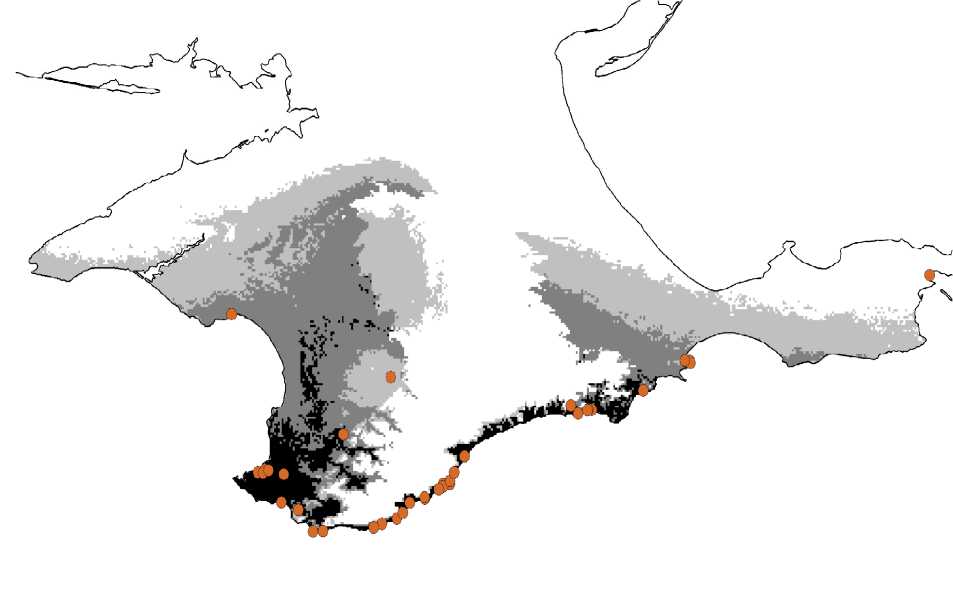
Рис. 2. Потенциальный ареал E. tauricus в Крыму. Области с оптимальными для вида климатическими параметрами отмечает черная заливка; области, пригодные для обитания вида в большей или меньшей степени, выделены, соответственно, темносерой и светло-серой заливкой; точками обозначены известные пункты его находок

Рис. 3. E. tauricus из г. Феодосия. Leg. О.В. Кукушкин и О.Г. Розенберг. Слева направо: juv. ♀, juv. ♂, ad. ♀
Ближайший пункт Юго-Восточного побережья Крыма, откуда известен крымский скорпион – г. Судак (Rathke, 1837; Бялыницкий-Бируля, 1917; Фет, 1989; наши данные). Этот портовый центр в древности поддерживал тесные торговые связи с Феодосией – Кафой. Таким образом, находка этого антропохорного вида в г. Феодосия, сохранившем многочисленные следы средневековой (генуэзской) застройки, была вполне ожидаемой. Дополнительные сведения о находках скорпионов в Феодосии, полученные позднее при опросах респондентов, приводятся в табл. 1. Надо сказать, что данный пункт ареала не является самым северным на востоке Крыма. Так, К.А. Ефетовым крымский скорпион обнаружен в г. Керчь (Фет, 1989). Однако на Керченском полуострове скорпион имеет явно менее широкое распространение, чем на Юго-Восточном побережье Горного Крыма, и, по-видимому, вообще встречается здесь крайне редко (возможно, уже исчез) (Рис. 2). Так, он не найден нами 7.05 и 3-4.09.2010 г.при тщательных поисках на хорошо сохранившихся массивных руинах позднеантичного городища Илурат в окрестностях с. Ивановка, а также в 2007-2010 гг. при многократных обследованиях городища Киммерик на западном склоне г. Опук. Краеведы Ю.Н. Ляшенко и В.В. Сербин сообщают, что скорпион в течение последних двух – трех десятилетий в г. Керчь и его окрестностях определенно не отмечался ни в частном секторе, ни на раскопках расположенных в припроливье многочисленных античных поселений. Не было его находок и на хорошо исследованном автором в 1998-2009 гг. мысе Чауда с его старым маяком и остатками поселений и постов боспорского и римского периодов. Для прояснения вопроса об обитании скорпиона в г. Керчь необходимо провести поиски в крепостях Керчь и Ени-Кале – наиболее вероятных, по нашему мнению, пунктах обитания вида.
C равнинного Западного побережья Крыма известны лишь две давних находки E. tauricus . Первая сделана в конце XIX в. куратором Зоологического музея Московского университета Н.М. Кулагиным на территории бывшего Евпаторийского уезда (Бялыницкий-Бируля, 1917). Вторая относится к 1913 г.: в музее Севастопольской биологической станции хранился экземпляр «из камня, привезенного из Евпатории» (Fet, 1988; Фет, 1989). Обе эти находки следует признать сомнительными, однако обитание скорпиона в г. Евпатория, тем не менее, вполне вероятно, учитывая роль этого города как порта на протяжении всей его истории и тесные древние связи античной Керкинитиды с Херсонесом. Результаты анализа потенциального ареала вида с использованием программы MAXENT также указывают на возможность сплошного распространения вида на участке между Севастополем и Евпаторией (рис. 2).
Таким образом, анализ распространения E. tauricus в Крыму выявляет абсолютную его синантропность, что, наряду с предварительными данными молекулярных исследований, проведенных другими исследователями, позволяет вынести предположение о его случайной интродукции здесь исторический период из Эгейского региона. Возможность завоза скорпиона в Крым из разных пунктов его ареала в южной части Эгейского моря возникала неоднократно – даже в новейшей истории. Так, во время проведения Первой Архипелажской экспедиции 1768-1774 гг. административный центр 27 населенных островов, находящихся под русской военно-морской администрацией, располагался именно на острове Парос [порт Ауза (Naousa)], а по окончании кампании, имевшей своими главными последствиями заключение Кучук-Кайнарджийского мира и открытие для российского торгового флота черноморских проливов, несколько тысяч архипелажских греков, ранее принявших русское подданство, переселились в Крым, где из них были сформированы гарнизоны, размещенные сперва в крепостях Керчь и Ени-Кале, а затем и по всему южному побережью от Феодосии до Балаклавы (Широкорад, 2006). Как говорилось выше, скорпион впервые был обнаружен в Крыму спустя два десятилетия после этих событий. Однако на сегодняшний день версия о проникновении E. tauricus в Таврику в античную эпоху (во время основания Херсонеса либо позже с Эгейского побережья Малой Азии или прилежащих островов) представляется наиболее реалистичной. Предположение о возможности неоднократного завоза скорпиона в Крым из различных близко расположенных пунктов Эгейского региона может быть подтверждено или опровергнуто при изучении генетической структуры его популяций в Крыму и Восточном Средиземноморье sensu stricto . При этом следует учитывать, что область, занимаемая E. tauricus на Крымском полуострове в настоящее время, по площади может существенно превосходить природный ареал предковой популяции. Весьма важным фактором, способствовавшим колонизации Южного Крыма одной из форм комплекса « E. tauricus », могла являться ее ксерофилия.
Список литературы Анализ распространения крымского скорпиона (Arachnida, Scorpiones, Euscorpiidae) c замечаниями о его биогеографическом статусе на Крымском полуострове
- Андреев В.М. Проблема Понтиды и долины древних рек в Черном море // Геология и полезные ископаемые Мирового океана. 2010. № 2. С. 47-50.
- Атлас. Автономна республiка Крим. Київ; Сiмферополь, 2003. 31 c.
- Бялыницкий-Бируля А.А. Фауна России и сопредельных стран. Паукообразные (Arachnoidea). Т. 1. Скорпионы. Петроград: Типография Импер. АН, 1917. 224 c.
- Виноградов Ю.Г., Золотарев М.И. Год рождения Херсонеса Таврического // Херсонесский сборник. 1998. Вып. IX. С. 36-46.
- Дузь С.Л., Кукушкин О.В., Назаров Р.А. О находке туркестанского геккона, Tenuidactylus fedtschenkoi (Sauria, Gekkonidae), в юго-западной Украине // Современная герпетология. 2012. Т. 12. № 3/4. С. 123-133.
