Анализ семенной продуктивности древесных пород в заповеднике «Уссурийский»

Автор: Орехова Т.П., Федина Л.А.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Растениеводство
Статья в выпуске: 5, 2013 года.
Бесплатный доступ
Фенологические наблюдения за цветением и семеношением деревьев проводили в заповеднике «Уссурийский» с 2002 по 2011 г. в широколиственно-кедровом лесу визуальным методом (по шкале Каппера) и с помощью семеномеров. Исследованиями установлено, урожайность семян за годы наблюдений значительно варьировала, максимальная семенная продуктивность отмечена 2004 г. В засушливые годы показатель пустосемянности у деревьев возрастает.
Деревья, семена, урожайность, качество семян, заповедник "уссурийский"
Короткий адрес: https://sciup.org/14083056
IDR: 14083056 | УДК: 630*181.522.523.231.1
The analysis of the arboreal species seed productivity in the natural reserve «Ussuriyskiy»
The phenological observation on tree flowering and seed productivity in the deciduous-cedar forest of the “Ussuriyskiy” Natural Reserve was conducted by the visual Capper's scale method and with the help of seed traps since 2002 to 2011. The research has established that seed yields during the observation years varied significantly, the maximum seed productivity is marked in 2004. The empty seed index of trees increases in dry years.
Текст научной статьи Анализ семенной продуктивности древесных пород в заповеднике «Уссурийский»
В заповеднике «Уссурийский» расположены уникальные по своему составу и строению кедровошироколиственные леса. Они длительное время не испытывали антропогенного влияния и поэтому представляют идеальную модель для изучения разнообразных процессов, происходящих в лесных экосистемах. На сопредельной территории еще в прошлом столетии были проведены Т.В. Самойловой [7] и продолжаются в заповеднике Л.А. Фединой [12] фенологические наблюдения за генеративными процессами у древесных видов, семенная же продуктивность большинства дальневосточных древесных пород пока изучена недостаточно полно. Материалы по семеношению ценных лесообразующих древесных пород фрагментарны. Обобщенные сведения о семенах дальневосточных древесных видов приведены в работах Н.В. Кречетовой с соавторами [3] и Т.П. Ореховой [6].
Цель настоящей работы – установить продолжительность фенофаз цветения и семеношения древесных видов, а также определить количественные и качественные показатели урожая деревьев в условиях заповедника «Уссурийский».
Материал и методы исследования. Наблюдения за семеношением проводили с 2002 по 20011 год на постоянной пробной площади (ППП) 7-2003 размером в 1 га, расположенной во влажном кленово-лещинограбовом кедровнике с липой и пихтой цельнолистной. Древесный ярус фитоценоза образован 675 деревьями, представлен 14 видами. Древостой образуют основные лиственные породы: 243 особи граба сердцевидного, 117 деревьев клена моно , 90 особей клена маньчжурского и 70 стволов ореха маньчжурского . Хвойные породы на данной площади немногочисленны: 40 деревьев сосны корейской и пихты цельнолистной. Крупные деревья в фитоценозе встречаются единично, среди них: ильм японский, ильм лопастный и орех маньчжурский. Наибольшие диаметры ствола (до 80 см) имели особи пихты цельнолистной. Учет урожая плодов и семян деревьев проводили методом семеномеров [2]. Cеменоловушки (36 шт.) были установлены на площади через каждые 20 м. Площадь одного семеномера составила 0,5 м2. Падающие в ловушки семена собирали с мая по ноябрь месяц. Часть ловушек находилась на площади в течение всего года для учета семян, опавших в зим-
„ S • q r ™ ний период. Величину урожая (P) находили по формуле P = ——, где размер пробной площади (S, м2); пло-a • n щадь поверхности одного семеномера (а); число взятых семеномеров (n); количество семян и плодов, собранных со всех семеномеров (q) [2]. Полученные данные пересчитывали на один гектар.
Качество семян устанавливали разными методами: путем взрезывания, окрашивания хлористым тетразолом [5], а также рентгенографическим методом [8] на аппарате Vix-60 (SOFTEX COLTD, Japan). По клас- су развития семян рассчитывали их жизнеспособность по методике Н.Г. Смирновой [8]. Жизнеспособность каждого образца (L) рассчитывали по формуле
_ 1/2( N 3 + N 4 + N 5) х 100% L —
N, где N3, N4, N5 – число семян III, IV, V классов развития в процентах от общего количества их в образце.
Жизнеспособность семян берез и ильмов устанавливали методом просвечивания [9]. При анализах использовали образцы семян в 5–10-кратной биологической повторности. Фенологические наблюдения проводили по методикам И.Н. Бейдеман [1], А.В.Тюрина [11] и ботаническим рекомендациям [13]. Урожайность древесных видов учитывали визуально по шкале Каппера [4]. Названия древесных пород даны согласно сводки «Сосудистые растения…, 1985–1996) [10].
Результаты и их обсуждение. Согласно фенологическим наблюдениям в среднем (за 10 лет) длительность фазы цветения у хвойных пород составила 12–13 дней (табл.1, 2). Однако в отдельные годы, например, в 2005 году, у Pinus koraiensis наблюдали длительное цветение побегов в течение 19 дней, в этот же год был зафиксирован максимальный урожай шишек. Длительность периода опадения шишек у хвойных пород составила в среднем до 1 месяца, но в отдельные годы эта фенофаза растягивалась до 53–55 дней (см. табл. 2). Так, в 2009 и 2010 годах наблюдали очень длительный период семеношения этого вида.
Для лиственных древесных пород период цветения длился в среднем от 12 до 19 дней. У Juglans mandshurica он составил 14 дней, а самая длительная фаза цветения была у Carpinus cordata – 19 дней. У лиственных древесных пород период диссеминации в среднем составил от 32 до 41 дня. Семена некоторых видов кленов и ясеня маньчжурского оставались висеть на деревьях до весны. Самым длительным в 2009 году был процесс опадения плодов у Juglans mandshurica – 67 дней.
Сведения о длительности репродуктивных фенофаз, полученные нами, согласуются с данными, приведенными Т.В. Самойловой за 1933 год [7]. Однако средние значения длительности фенофазы цветения и семеношения, рассчитанные за 10 лет наших наблюдений, оказались значительно ниже (см. табл.1,2). Современные условия произрастания древесных видов, вероятно, отличаются от климата середины прошлого столетия.
Анализ наблюдений за семенной продуктивностью (по шкале Каппера) показал, что невозможно установить какую-либо периодичность в плодоношении наблюдаемых древесных видов (табл. 1). Например, у Pinus koraiensis отмечали отсутствие семеношения в 2002, 2003, 2006 и 2010 годах. Как правило, после урожайного года наблюдалось или понижение, или полное отсутствие семян. За 10-летний период наблюдений в условиях заповедника урожай в 1–2 балла у Pinus koraiensis был отмечен в 2005 и 2009 годах. У пихты цельнолистной отсутствие семеношения наблюдали в 2006 и 2007 годах. Тем не менее, перед периодом отсутствия семян и после него эта древесная порода формировала урожай интенсивностью в 1–3 балла почти ежегодно. Однако качество семян пихты часто оставалось низким. Высокая жизнеспособность семян (до 80%) выявлена только в 2004 и 2009 годах.
Интенсивность семеношения древесных пород, баллы по шкале Каппера
Таблица 1
|
Вид |
2002 г. |
2003 г. |
2004 г. |
2005 г. |
2006 г. |
2007 г. |
2008 г. |
2009 г. |
2010 г. |
2011 г. |
|
Pinus koraiensis |
0 |
0 |
1 |
1–2 |
0 |
1 |
0–1 |
2 |
0 |
0–1 |
|
Abies holophylla |
3 |
1–2 |
3 |
2 |
0 |
0 |
1 |
3 |
3 |
2 |
|
Quercus mongolica |
2 |
1 |
5 |
1 |
2 |
1 |
1–2 |
3 |
4 |
3 |
|
Carpinus cordata |
1 |
1 |
3 |
2 |
3 |
2 |
1 |
2 |
3 |
3 |
|
Acer mono |
5 |
0–1 |
4 |
2 |
2 |
4 |
1 |
4 |
4 |
4 |
|
Acer mandshuricum |
3 |
0–1 |
4 |
1 |
1 |
3 |
1 |
4 |
3 |
4 |
|
Ligustrina amurensis |
2 |
2 |
4 |
1 |
2 |
4 |
1 |
1 |
2 |
3 |
|
Fraxinus mandshurica |
1 |
1 |
3 |
2 |
3 |
3 |
1 |
2 |
5 |
4 |
|
Juglans mandshurica |
1 |
1 |
2 |
3 |
3 |
2 |
1 |
5 |
1 |
3 |
Таблица 2
|
Вид |
2002 г. |
2003 г. |
2004 г. |
2005 г. |
2006 г. |
2007 г. |
2008 г. |
2009 г. |
2010 г. |
2011 г. |
Варьирование длительности фенофаз и их среднее значение (дней) |
|
Pinus koraiensis |
* 0 |
13 |
0 |
19 |
9 |
9 |
14 |
11 |
9 |
0 |
9–19 (12) |
|
**0 |
0 |
0 |
29 |
0 |
20 |
29 |
51 |
53 |
0 |
20–53 (36) |
|
|
Abies holophylla |
9 |
19 |
16 |
0 |
0 |
0 |
19 |
9 |
11 |
13 |
9–19 (13) |
|
57 |
31 |
27 |
0 |
0 |
0 |
29 |
55 |
42 |
31 |
27–57 (27) |
|
|
Quercus mon-golica |
11 |
8 |
15 |
18 |
12 |
10 |
12 |
8 |
8 |
25 |
8–25 (12) |
|
35 |
30 |
30 |
31 |
40 |
29 |
42 |
52 |
39 |
31 |
29–52 (36) |
|
|
Carpinus cordata |
21 |
17 |
25 |
25 |
28 |
27 |
19 |
13 |
9 |
12 |
9–28 (19) |
|
41 |
37 |
25 |
32 |
26 |
53 |
30 |
26 |
22 |
31 |
22–53 (32) |
|
|
Acer mono |
23 |
12 |
14 |
7 |
8 |
5 |
18 |
9 |
8 |
11 |
5–23 (11) |
|
33 |
52 |
43 |
35 |
39 |
39 |
54 |
42 |
32 |
32 |
32–54 (40) |
|
|
Acer mandshuricum |
12 |
12 |
20 |
11 |
9 |
5 |
8 |
19 |
15 |
8 |
5–20 (11) |
|
52 |
24 |
55 |
41 |
39 |
32 |
43 |
43 |
21 |
58 |
21–58 (40) |
|
|
Ligustrina amurensis |
8 |
10 |
16 |
8 |
8 |
17 |
21 |
6 |
8 |
8 |
6–21(11) |
|
36 |
29 |
46 |
30 |
29 |
24 |
23 |
38 |
28 |
63 |
23–63 (34) |
|
|
Fraxinus mandshurica |
13 |
17 |
11 |
11 |
9 |
9 |
12 |
11 |
17 |
11 |
9-17 (12) |
|
41 |
31 |
28 |
32 |
39 |
42 |
40 |
35 |
46 |
56 |
28–56 (39) |
|
|
Juglans mandshurica |
21 |
8 |
14 |
13 |
15 |
17 |
10 |
8 |
15 |
25 |
8–25 (14) |
|
41 |
31 |
30 |
42 |
21 |
57 |
46 |
67 |
35 |
48 |
21–67 (41) |
* Продолжительность цветения, дн.; ** продолжительность семеношения, дн.
Длительность фенофаз цветения и семеношения деревьев в заповеднике «Уссурийский»
Урожаи плодов и семян отдельных видов составляют общий урожай всего фитоценоза. Объем урожая всего фитоценоза оценивался нами по поступлению плодов и семян в ловушки. Результаты учетов представлены на рисунке 1. Семенная продуктивность деревьев значительно варьировала за период наблюдений. Минимальный урожай отмечали в 2003 году, а максимальный – в следующем году. Этот год и следует считать семенным. В 2007 и 2009 годах произошло незначительное повышение семенной продуктивности деревьев (см. рис.1).
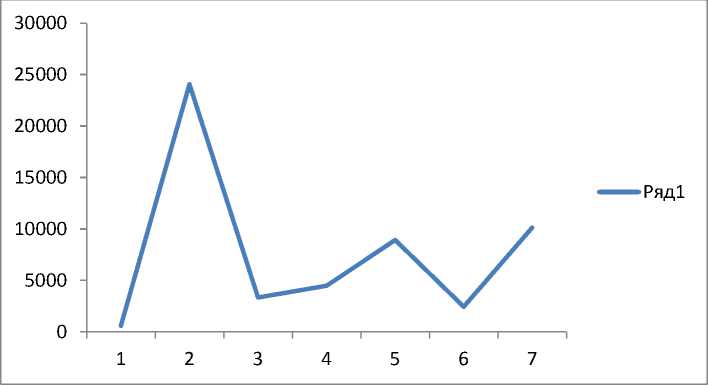
Рис.1. Динамика семенной продуктивности древесных пород на ППП7-2003 (шт/га):
1 – 2003 г., 2 – 2004 г.; 3 – 2005 г.; 4 – 2006 г.; 5 – 2007 г.; 6 – 2008 г.; 7 – 2009 г.
Как видно из кривой урожайности, сразу после обильного семеношения деревьев обычно наступает 1–2-летний период уменьшения семенной продуктивности. Вероятно, такая занономерность связана с физиологическим состоянием деревьев, которым необходимо прежде всего накопить в тканях достаточное количество запасных соединений для формирования будущего урожая. Особое внимание нами уделялось изучению семенной продуктивности хвойных лесообразующих пород. Динамика урожая сосны корейской и пихты цельнолистной представлена на рисунке 2. Максимальный урожай семян пихты цельнолистной на исследуемой территории зафиксирован только в 2004 году. В последующие годы семеношение оставалось очень низким, а его повышение произошло только в 2007 году. Максимальный урожай шишек у сосны корейской наблюдали только в 2005 году, затем семенная продуктивность деревьев также понизилась. Пики максимальной урожайности этих древесных пород сдвинуты на один год, поскольку сроки формирования репродуктивных органов у видов различны. Формирование шишки сосны корейской происходит в течение трех вегетационных периодов.
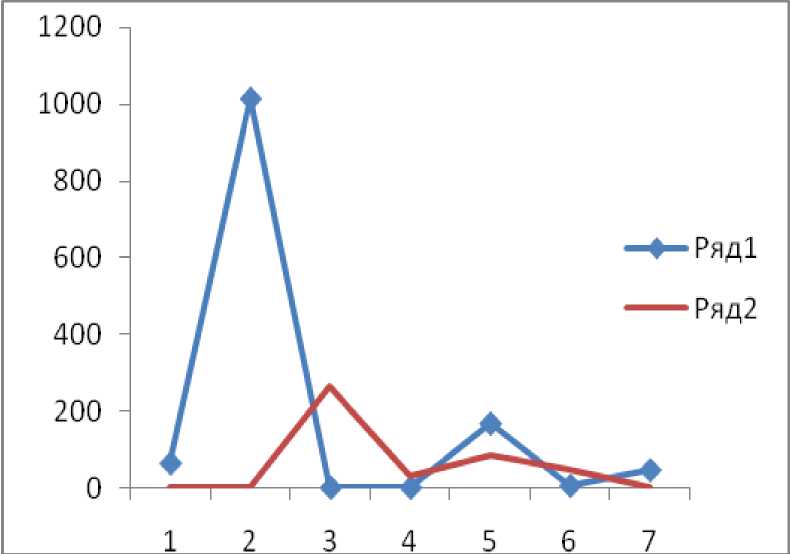
Рис. 2. Урожай семян хвойных пород на ППП7-2003 (шт/га): 1 – 2003 г.; 2 – 2004 г.; 3 – 2005 г.; 4 – 2006 г.;
5 – 2007 г.; 6 – 2008 г.; 7 – 2009 г.; ряд 1 – количество семян пихты цельнолистной;
ряд 2 – количество семян сосны корейской
Урожай и качество семян лиственных древесных породы изменялось каждый год. Орешки липы амурской, маньчжурской и Таке почти всегда имели очень низкую жизнеспособностью. Пустосемянность достигала в отдельные годы 60–83%. В засушливые летние периоды (июнь-июль) 2005–2007 годов наблюдали периоды, когда деревья сбрасыванили множество еще недоразвитых семян или цветочных бутонов. Вероятно, липа так реагирует на водный дефицит. Количество сброшенных семян составило более 55% всего урожая. В урожае крылаток клена маньчжурского и к. моно обычно находилось от 13 до 33% пустых семян. Высоким качеством семян из всех древесных пород отличались из всех лиственных пород только ясень маньчжурский и граб сердцевидный. Пустосемяннность у этих видов не превышала 10–15%.
Известно, что на формирование репродуктивных органов большое влияние оказывают климатические условия. Мы попытались определить влияние осадков, выпавших в год формирования плодов, на процент пустосемянности у древесных видов (рис. 3).
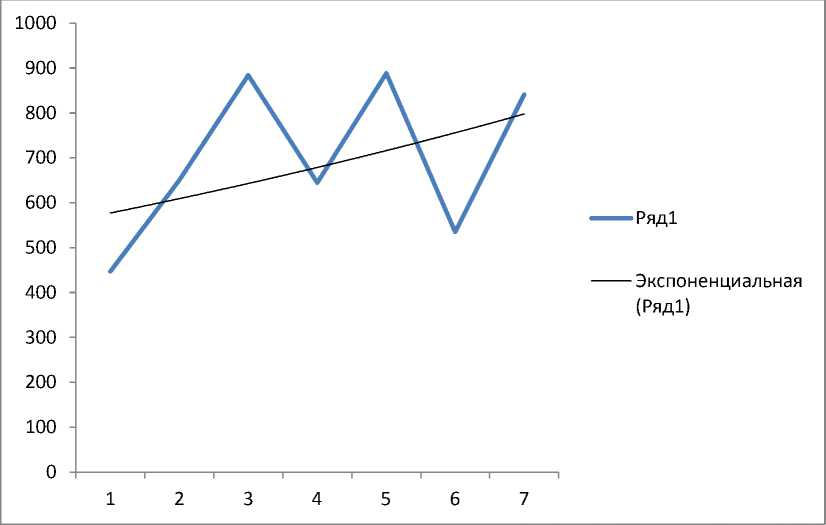
Рис. 3. Общее количество осадков (мм в год): 1 – 2003 г., 2 – 2004 г.; 3 – 2005 г.; 4 – 2006 г.; 5 – 2007 г.; 6 – 2008 г.; 7– 2009 г.
Оказалось, что эти показатели имеют обратно пропорциональную зависимость. Об этом свидетельствуют кривые тренда рассмотренных данных (рис. 3,4). Чем меньшее количество выпадает за год осадков, тем большее количество пустых семян образуется на деревьях (см. рис. 3, 4). Например, самый большой объем пустых семян (более 90%) зафиксирован в 2003 году, когда выпало наименьшее (450 мм) количество осадков. Минимальное количество пустых семян у деревьев обнаружено в заповеднике в 2005 и 2007 годах, в эти годы количество выпавших осадков превысило 900 мм.
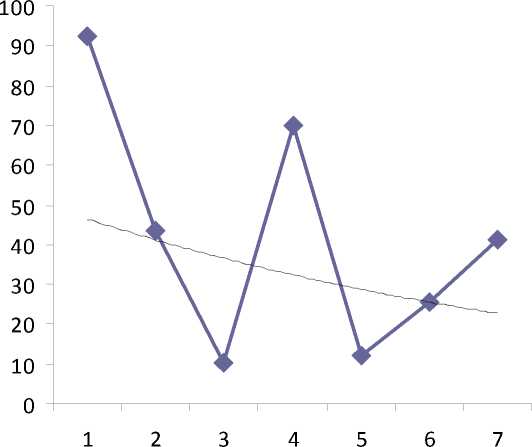
Ряд1
Экспоненциальный (Ряд1)
Рис.4. Динамика пустосемянности у древесных видов на ППП7-2003 (%):
1 – 2003 г., 2 – 2004 г.; 3 – 2005 г.; 4 – 2006 г.; 5 – 2007 г.; 6 – 2008 г.; 7– 2009 г.
Выводы
Таким образом, в результате проведенных нами 10-летних наблюдений установлено, что длительность фенологических фаз цветения у изученных видов деревьев в среднем составляет от 1 до 3 недель, а период диссеминации растянут от 1 до 2 месяцев. В зависимости от климатических параметров изменялись как сроки, так и длительность фенофаз. Не установлено какой-либо периодичности в плодоношении деревьев в заповеднике «Уссурийский».
В засушливый летний период обнаружено, что у деревьев из родов Tilia и Acer происходит опадение части сформированных цветков и недоразвитых плодов. Вероятно, это является защитной реакцией растений на водный дефицит.
Установлено, что процент пустых и недоразвитых семян у деревьев имеет обратно пропорцианаль-ную зависимость с количеством осадков, выпавших в год формирования этих семян.
