Анализ спектра питания озерной лягушки (Rana ridibunda) на урбанизированных территориях Среднего Поволжья
 на урбанизированных территориях Среднего Поволжья")
Автор: Файзулин А.И., Чихляев И.В., Кривошеев В.А., Кузовенко А.Е.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 1-1 т.12, 2010 года.
Бесплатный доступ
Исследовано питание озерной лягушки Rana ridibunda Pallas, 1771 в условиях различной по степени урбанизации участках (с преимущественно малоэтажной застройкой, в лесопарковой и пригородной зонах) и в контроле. Наибольшая ширина трофической ниши по индексу полидоминантности Sλ отмечена в контроле (Sλ = 17,28), пригородной зоне (Sλ = 16,58), лесопарке (Sλ = 16,38), а наименьшая (Sλ = 11,53) - в условиях малоэтажной застройки.
Питание, среднее поволжье
Короткий адрес: https://sciup.org/148205474
IDR: 148205474 | УДК: 591:
The analysis of food spectrum of a lake frog (rana ridibunda) in the urbanization territory of the Middle Volga region
A food of lake frog Rana ridibunda, Pallas, 1771 was investigated in the sites with a variable degree of an urbanization conditions, in districts mainly with low-storeyed buildings, in a forest park zone, in a residential suburb and in the control. The greatest trophic niche width, according the Sλ index was noted in control conditions (Sλ = 17,28), residential suburb (Sλ = 16,58), in a forest park zone (Sλ = 16,38) and the least (Sλ = 11,53) in the conditions of sites with low-storeyed buildings.
Текст научной статьи Анализ спектра питания озерной лягушки (Rana ridibunda) на урбанизированных территориях Среднего Поволжья
В Поволжье наиболее эвритопным и синантропным видом земноводных является озерная лягушка [4]. В условиях урбанизированной среды питание амфибий исследовано в Подмосковье [9], городах Екатеринбург [2], Воронеж [1] и Липецк [10]. Питание озерной лягушки на урбанизированных территориях Среднего Поволжья ранее не исследовалось. В черте городов можно выделить несколько зон по степени урбанизации с доминирующим типом застройки территории: I - промышленная, II - многоэтажная, III - малоэтажная, также выделяют IV - территорию лесопарков и пригородных лесных массивов и V (К) - контроль, участок, удаленный от черты города [3].
Цель настоящего исследования - анализ и сравнение спектров питания озерной лягушки, обитающей в различных условиях урбанизации в черте г. Тольятти Самарской области.
МАТЕРИАЛ И МЕТОДИКА
Изучение питания озерной лягушки проведено в период с 20 мая по 20 июня 2005 г. в 3 географических пунктах г. Тольятти с различными условиями урбанизации и в контроле: 1 - «малоэтажная застройка» (n = 66) - пойменное оз. Машкино, в зоне с малоэтажной застройкой пос. Федоровка; 2 - «лесопарковая» (n = 76) - пруд ливневой канализации Центрального района города, расположен в лесопарке, примыкающем к зоне с многоэтажной застройкой (ул. Баныкина); 3 - «пригородная зона» (n = 52) - прилегающие к черте г. Тольятти озера Васильевские около с. Васильевка Ставропольского района Самарской области; 4 - «контроль» (n = 59) - пойменные озера НП
Для обобщенной характеристики спектра питания использовали индекс Симпсона для выборочных значений X [14]. Ширину трофической ниши рассчитывали по показателю поли доминантности S x [7], который равен отношению 1 к индексу концентрации Симпсона [14]. Перекрывание трофических ниш оценивали по индексу Мориситы I [7].
У зеленых лягушек отсутствует пищевая специализация, за исключением размерной дифференциации [8], поэтому спектр питания популяции отражает реализованную трофическую нишу. При этом трофическая ниша включает 2 стации - водоем и береговую часть (обычно до 50 м). Соответственно, четко выделяются объекты, добываемые в водоеме, передвигающиеся по поверхности суши и летающие. Для статистической оценки трофическая ниша подразделена на две части - «водную» и «наземную».
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Из 233 пищевых объектов 51,5% (120 экз.) определены до вида, 25,3% (59 экз.) - до рода, 15,0% (54 экз.) - до семейства, 8,2% (38 экз.) -до отряда.
Данные о встречаемости пищевых объектов в условиях различной антропопрессии представлены в таблице. Независимо от степени урбанизации, значительную долю в питании составляют жужелицы (13,5-17,0%), например, Pterostichus niger (до 5,7%).
Таблица. Спектр питания популяций озерной лягушки Rana ridibunda на территориях с различной степенью урбанизации
|
Пищевые объекты |
Зоны (по степени урбанизации) |
|||||||
|
Малоэтажная |
Лесопарковая |
Пригородная |
Контроль |
|||||
|
экз. |
% |
экз. |
% |
экз. |
% |
экз. |
% |
|
|
Тип Arthropoda Класс Insecta Отряд Trichoptera, larvae |
4 |
6,1 ± 2,9 |
1 |
1,32 ± 1,31 |
- |
0 |
- |
0 |
|
Отряд Odonata, larvae |
- |
0 |
- |
0 |
3 |
4,8 ± 2,7 |
2 |
3,4 ± 2,4 |
|
П/отряд Zygopterа |
- |
0 |
1 |
1,32 ± 1,31 |
2 |
3,2 ± 2,2 |
- |
0 |
|
Семейство Coenagrionidae |
3 |
4,5 ± 2,6 |
- |
0 |
- |
0 |
- |
0 |
|
П/отряд Anisoptera |
- |
0 |
- |
0 |
2 |
3,2 ± 2,2 |
- |
0 |
|
Отряд Hymenoptera (ближе не опред.) |
- |
0 |
2 |
2,6 ± 1,8 |
3 |
4,8 ± 2,7 |
4 |
6,8 ± 3,3 |
|
Семейство Apoidea |
1 |
1,5 ± 1,5 |
4 |
5,3 ± 2,6 |
1 |
1,6 ± 1,6 |
6 |
10,2 ± 3,9 |
|
Семейство Vespidae |
2 |
3,0 ± 2,1 |
4 |
5,3 ± 2,6 |
2 |
3,2 ± 2,2 |
- |
0 |
|
Семейство Sphecidae |
- |
0 |
1 |
1,3 ± 1,2 |
3 |
4,8 ± 2,7 |
1 |
1,7 ± 1,6 |
|
Семейство Formicidae |
1 |
1,5 ± 1,5 |
5 |
6,6 ± 2,8 |
- |
0 |
6 |
10,2 ± 3,9 |
|
Отряд Coleoptera |
2 |
3,0 ± 2,1 |
1 |
1,3 ± 1,2 |
2 |
3,2 ± 2,2 |
- |
0 |
|
Семейство Coccinellidae |
3 |
4,5 ± 2,6 |
- |
0 |
1 |
1,6 ± 1,6 |
- |
0 |
|
Семейство Carabidae |
10 |
15,2 ± 4,4 |
9 |
11,8 ± 3,7 |
9 |
14,5 ± 4,5 |
8 |
13,6 ± 4,5 |
|
Семейство Curculionidae |
2 |
3,0 ± 2,1 |
5 |
6,6 ± 2,8 |
4 |
6,5 ± 3,1 |
7 |
11,9 ± 4,2 |
|
Семейство Hydrophilidae |
- |
0 |
1 |
1,3 ± 1,2 |
2 |
3,2 ± 2,2 |
1 |
1,7 ± 1,6 |
|
Семейство Buprestidae |
7 |
10,6 ± 3,8 |
- |
0 |
2 |
3,2 ± 2,2 |
2 |
3,4 ± 2,4 |
|
Семейство Chrysomelidae |
14 |
21,2 ± 5,0 |
1 |
1,3 ± 1,2 |
5 |
8,1 ± 3,5 |
2 |
3,4 ± 2,4 |
|
Семейство Silphidae |
3 |
4,5 ± 2,6 |
2 |
2,6 ± 1,8 |
3 |
4,8 ± 2,7 |
1 |
1,7 ± 1,6 |
|
Семейство Scarabidae |
- |
0 |
2 |
2,6 ± 1,8 |
- |
0 |
2 |
3,4 ± 2,4 |
|
Семейство Staphylinidae |
- |
0 |
1 |
1,3 ± 1,2 |
- |
0 |
- |
0 |
|
Семейство Elateridae |
- |
0 |
- |
0 |
- |
0 |
1 |
1,7 ± 1,6 |
|
Семейство Dytiscidae |
2 |
3,0 ± 2,1 |
1 |
1,3 ± 1,2 |
2 |
3,2 ± 2,2 |
2 |
3,4 ± 2,4 |
|
Отряд Hemiptera |
0 |
7 |
9,2 ± 3,3 |
1 |
1,6 ± 1,6 |
3 |
5,1 ± 2,8 |
|
|
Семейство Scutelleridae |
- |
0 |
1 |
1,3 ± 1,2 |
1 |
1,6 ± 1,6 |
- |
0 |
|
Семейство Pentatomidae |
0 |
- |
0 |
- |
0 |
2 |
3,4 ± 2,4 |
|
|
Отряд Ortoptera |
- |
0 |
- |
0 |
- |
0 |
2 |
3,4 ± 2,4 |
|
Семейство Tettigonidae |
- |
0 |
1 |
1,3 ± 1,3 |
1 |
1,6 ± 1,6 |
1 |
1,7 ± 1,6 |
|
Отряд Homoptera |
- |
0 |
- |
0 |
- |
0 |
1 |
1,7 ± 1,6 |
|
П/отряд Cicadinea |
- |
0 |
4 |
5,3 ± 2,6 |
2 |
3,2 ± 2,2 |
- |
0 |
|
Семейство Aphrophoridae |
1 |
1,5 ± 1,5 |
- |
0 |
1 |
1,6 ± 1,6 |
2 |
3,4 ± 2,4 |
|
Отряд Diptera |
1 |
1,5 ± 1,5 |
- |
0 |
- |
0 |
- |
0 |
|
Отряд Dermaptera |
1 |
1,5 ± 1,5 |
- |
0 |
- |
0 |
3 |
5,1 ± 2,9 |
|
Класс Arachnida |
3 |
4,5 ± 2,6 |
- |
0 |
1 |
1,6 ± 1,6 |
- |
0 |
|
Тип Mollusca Класс Gastropoda |
2 |
3,0 ± 2,1 |
2 |
2,6 ± 1,8 |
1 |
1,6 ± 1,6 |
- |
0 |
|
Класс Bivalvia |
- |
0 |
1 |
1,3± 1,2 |
4 |
6,5 ± 3,1 |
- |
0 |
|
Тип Chordata П/тип Vertebrata Класс Mammalia |
- |
0 |
- |
0 |
2 |
3,2 ± 2,2 |
- |
0 |
|
Отряд Rodentia |
- |
0 |
4 |
5,3 ± 2,6 |
2 |
3,2 ± 2,2 |
- |
0 |
|
Класс Amphibia Отряд Anura |
- |
0 |
3 |
3,9 ± 2,2 |
- |
0 |
- |
0 |
|
Класс Pisces |
4 |
6,1 ± 2,9 |
12 |
15,8 ± 4,3 |
- |
0 |
- |
0 |
В зоне малоэтажной застройки доминируют в питании жуки-листоеды, также высока их доля в условиях пригорода, а в зоне контроля доля в питании ниже. Обычны в рационе муравьи (в условиях лесопарковой зоны и контроля 9,4—10,1%), представленные 5 видами — Formica fusca, F. rufa, F. imitans, F. cunicularia и Lasius niger. Остальные перепончатокрылые, представленные летающими формами, составляют 18,8% и 16,9% от всех экологических форм в рационе. Это — осы Polistes dominulus, Vespula vulgaris, пчелы Apis mellifera, Bombus sp., а также Polistes nimpha, Eucera sp., Halict sp., Polistes sp., Trypoxylon sp.
Наиболее высокая доля жуков-долгоносиков (7 экз. 5 видов) обнаружена в питании озерной лягушки на контрольном участке — Otiorrhynchus fullo , O. ovatus , O. velutinus, Pseu-docleonus cinereus и Hylobius transversovittatus . В лесопарковой зоне и в зоне малоэтажной застройки отмечены позвоночные животные: мальки и молодь рыб Carassius auratus , Perca fluviatilis , личинки озерной лягушки, а также мелкие млекопитающие — Sorex sp.
Фоновыми видами в условиях лесопарковой зоны являются жуки-златки Eurythyua guercus и брюхоногие моллюски Planorbaris corneus , Physa fontinalis . В условиях пригорода отмечены двустворчатые моллюски Dreissina polymor-pha — 6,5%. На порядок ниже встречаемость в рационе жуков-мертвоедов ( Silpha obscura , S. carinata ) , щелкунов ( Agriotes lineatus, A. obscures ), хрущей Melolontha hippocastani , Apho-dius varians ), чернотелки ( Crypticus quisquilius ), карапузика ( Hister bipustulatus ), плавунцов ( Co-lymbetes striatus ), уховертки ( Labidura rip aria ). Наиболее редки в питании клопы, водные формы которых представлены водомерками Gerris lacustris , гладышем Plea leachi , плавтом Ilyocoris cimicoides и семейством Corixidae; наземные формы клопов представлены семействами Myodochidae и Cydnidae.
Оценка ширины трофической ниши представлена на рис. 1. Судя по графику, наибольшая ширина трофической ниши наблюдается в условиях пригорода, наименьшая — в зоне малоэтажной застройки. При этом в зоне условного контроля трофическая ниша уже, чем в пригородной зоне. Следует отметить, что ширина трофической ниши у видов без пищевой специализации отражает кормовую базу. Известно, что трансформация среды приводит к снижению фаунистического разнообразия беспозвоночных животных, например, жужелиц, составляющих значительную долю рациона озерной лягушки. Возрастание значения «меры концентрации» говорит о доминировании в питании отдельных групп пищевых объектов, например, особи озерной лягушки питаются молодью рыб в скоплениях.
В наземной части трофической ниши озерная лягушка питается ползающими (жуки, клопы, муравьи), прыгающими (прямокрылые, цикады) и летающими (стрекозы, двукрылые, пчелы, осы) пищевыми объектами; в водной части — плавающими (водные жуки, личинки насекомых, мальки и молодь рыб), ползающими (брюхоногие и двустворчатые моллюски).
Соотношение наземных и водных объектов в питании озерной лягушки приведено на рис. 2.
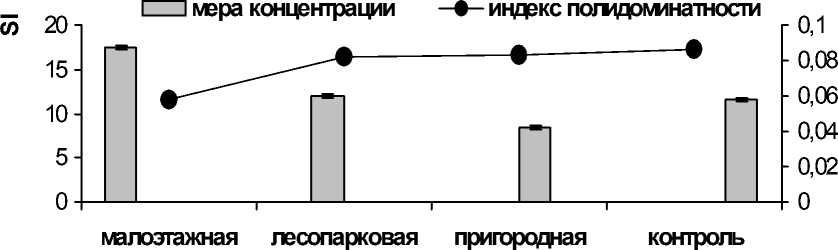
Рис. 1. Ширина трофической ниши (S A ) и индекс полидоминантности ( А ) по данным спектра питания озерной лягушки из зон с разной степенью урбанизации
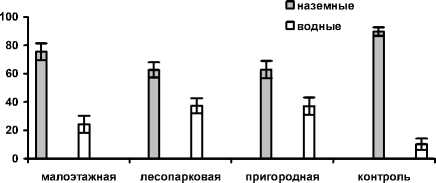
Рис. 2. Экологические формы (водные и наземные) объектов питания в рационе озерных лягушек из зон с разной степенью урбанизации
Различия по встречаемости экологических групп между выборками из лесопарковой, пригородной и малоэтажной зон статистически недостоверны на 5%-ном уровне значимости, при этом доля водных форм здесь составляет 24,2—37,3%. В условиях контроля эта доля (10,2%) ниже по сравнению с остальными вы борками, однако статистически достоверных различий с ними не выявлено.
Оценка перекрывания трофических ниш проведена по индексу Мориситы I x . В нашем случае данный индекс показывает сходство трофических спектров. Наибольшее сходство отмечено для популяции, обитающей в зоне условного контроля, с пригородной зоной (I x = 0,860) и с лесопарком (I x = 0,761); наиболее отличается спектр питания в контроле от зоны малоэтажной застройки (I x = 0,568). При этом рацион в зоне малоэтажной застройки сходен с пригородной зоной (I x = 0,844), наименьшее сходство — с лесопарковой зоной (I x = 0,532). Также сходны рационы в пригороде и лесопарковой зоне (I x = 0,717).
По данным В.Л. Вершинина [2], в Екатеринбурге отмечаются изменения в структуре пищевых связей сеголеток остромордой лягушки в основном за счет их укорочения и повышения прессинга на почвенных беспозвоноч- ных, разнообразие и численность которых снижены в городской черте. Сходные данные получены для Подмосковья [9]. Для г. Воронежа в питании озерной лягушки отмечены гидробионты (Gastropoda, Odonata, larvae) при отсутствии рыб; из наземных кормов доминируют хортобионты (Odonata, Aphididae, Chrysomelidae, Curculionidae, Apionidae), антобионты включают паразитических Hymenoptera, Heleidae, Bibioni-dae, Ephydridae, а герпетобионты представлены семействами Carabidae, Formicidae [1]. По данным для г. Липецка, основу питания озерной лягушки, независимо от степени антропогенной нагрузки на биотоп, составляют моллюски (34,5%), жесткокрылые (27,1%), пауки (9,5%), полужесткокрылые (7,0%) и стрекозы (6,9%) [10].
Таким образом, наши данные по питанию озерной лягушки в условиях г. Тольятти в целом сходны с данными для городов Воронеж и Липецк. В монографии С.Л. Кузьмина [8] приводятся сведения, судя по которым, спектр питания озерной лягушки значительно отличается в разных частях ареала. По нашим данным, спектр питания озерной лягушки в условиях урбанизации имеет существенные различия по сравнению с зоной контроля. Это связано с особенностями кормовой базы и с изменениями трофической ниши — повышением доли «водных» пищевых объектов, например, молоди рыб (15,8%) в лесопарковой зоне и зоне малоэтажной застройки г. Тольятти.
Авторы благо дарят Г.С. Розе нберга за ценные замечания и |П.И. Антонова) за помощь при определении пищевых объектов.
ÑÏÈÑÎÊ ËÈÒÅÐÀÒÓÐÛ
-
1. Бутов Г.С., Хиирва Л.Н. Особенности питания земноводных и пресмыкающихся в антропогенных биотопах г. Воронежа // Вести. Воронеж. ун-та 2003. № 2. С. 108-115.
-
2. Вершинин ВЛ. Динамика питания сеголеток бурых лягушек в период завершения метаморфоза // Экология, 1995. № 6. С.68-75.
-
3. Вершинин В.Л. Экологические особенности популяций амфибий урбанизированных территорий: Авто-реф. дне.... д-ра биол. наук. Екатеринбург, 1997. 47 с.
-
4. Гаранин В.И. Земноводные и пресмыкающиеся Волжско-Камского края. М.: Наука, 1983. 175 с.
-
5. Дробенков С.М. Трофическая структура и межвидовые взаимоотношения в сообществах земноводных и пресмыкающихся - энтомофагов наземных и прибрежных биогеоценозов Беларуси // Вопросы герпетологии. Материалы третьего съезда Герпетол. о-ва им. А.М. Никольского. Пущино; М.: МГУ, 2008. С.112-116.
-
6. Кузьмин С .Л. Теория оптимального пищедобывания и избирательное питание земноводных // Ж урн. общ. биол. 1992. Т. 53, № 1. С. 47-55.
-
7. Кузьмин С.Л. Трофология хвостатых земноводных. Экологические и эволюционные аспекты. М.: Наука, 1992. 170 с.
-
8. Кузьмин СЛ. Земноводные бывшего СССР. М.: Т-во науч. изд. КМК, 1999. 298 с.
-
9. Леонтьева ОА Бесхвостые земноводные как биоиндикаторы антропогенных изменений в экосистемах: Автореф. дис. ... канд. биол. наук. М., 1990. 20 с.
-
10. Никашин И.А. Эколого-морфологические признаки популяций озерной лягушки ( Rana ridibunda Pall.) как средство оценки антропогенного воздействия на водные экосистемы (на примере Липецкой области): Автореф. дис. ... канд. биол. наук. Липецк, 2007. 20 с.
-
11. Определитель насекомых европейской части СССР. Т. I. М.- Л.: Наука. 1964. 845 с.
-
12. Определитель насекомых Европейской части СССР. Т. II. М.-Л.: Наука. 1965. 668 с.
-
13. Определитель насекомых Европейской части СССР. Т. III. Перепончатокрылые Л.: Наука. 1978. 584 с.
-
14. Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 287 с.
-
15. Шляхтин Г.В., Голикова В.Л. Методика полевых исследований экологии амфибий и рептилий. Саратов: Изд-во Сарат. ун-та, 1986. 78 с.
-
16. Ясюк В.П. Водные моллюски: Определитель, краткий справочник по экологии водных моллюсков Самарской области. Самара, 2005. 92 с .
THE ANALYSIS OF FOOD SPECTRUM OF A LAKE FROG (RANA RIDIBUNDA) IN THE URBANIZATION TERRITORY OF THE MIDDEL VOLGA REGION
institute of ecology of the Volga River basin RAS, Togliatti 2Ulyanovsk State Pedagogical University, Ulyanovsk 3 Samara zoological park, Samara
A food of lake frog Rana ridibunda, Pallas, 1771 was investigated in the sites with a variable degree of an urbanization conditions, in districts mainly with low-storeyed buildings, in a forest park zone, in a residential suburb and in the control. The greatest trophic niche width, according the S x index was noted in control conditions (S x = 17,28), residential suburb (S x = 16,58), in a forest park zone (S x = 16,38) and the least (S x = 11,53) in the conditions of sites with low-storeyed buildings.
Список литературы Анализ спектра питания озерной лягушки (Rana ridibunda) на урбанизированных территориях Среднего Поволжья
- Бутов Г.С., Хицова Л.Н. Особенности питания земноводных и пресмыкающихся в антропогенных биотопах г. Воронежа//Вестн. Воронеж. ун-та. 2003. № 2. С. 108-115.
- Вершинин В.Л. Динамика питания сеголеток бурых лягушек в период завершения метаморфоза//Экология, 1995. № 6. С.68-75.
- Вершинин В.Л. Экологические особенности популяций амфибий урбанизированных территорий: Автореф. дис.… д-ра биол. наук. Екатеринбург, 1997. 47 с.
- Гаранин В.И. Земноводные и пресмыкающиеся Волжско-Камского края. М.: Наука, 1983. 175 с.
- Дробенков С.М. Трофическая структура и межвидовые взаимоотношения в сообществах земноводных и пресмыкающихся -энтомофагов наземных и прибрежных биогеоценозов Беларуси//Вопросы герпетологии. Материалы третьего съезда Герпетол. о-ва им. А.М. Никольского. Пущино; М.: МГУ, 2008. С.112-116.
- Кузьмин С.Л. Теория оптимального пищедобывания и избирательное питание земноводных//Журн. общ. биол. 1992. Т. 53, № 1. С. 47-55.
- Кузьмин С.Л. Трофология хвостатых земноводных. Экологические и эволюционные аспекты. М.: Наука, 1992. 170 с.
- Кузьмин С.Л. Земноводные бывшего СССР. М.: Т-во науч. изд. КМК, 1999. 298 с.
- Леонтьева О.А. Бесхвостые земноводные как биоиндикаторы антропогенных изменений в экосистемах: Автореф. дис. … канд. биол. наук. М., 1990. 20 с.
- Никашин И.А. Эколого-морфологические признаки популяций озерной лягушки (Rana ridibunda Pall.) как средство оценки антропогенного воздействия на водные экосистемы (на примере Липецкой области): Автореф. дис. … канд. биол. наук. Липецк, 2007. 20 с.
- Определитель насекомых европейской части СССР. Т. I. М.-Л.: Наука. 1964. 845 с.
- Определитель насекомых Европейской части СССР. Т. II. М.-Л.: Наука. 1965. 668 с.
- Определитель насекомых Европейской части СССР. Т. III. Перепончатокрылые Л.: Наука. 1978. 584 с.
- Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 287 с.
- Шляхтин Г.В., Голикова В.Л. Методика полевых исследований экологии амфибий и рептилий. Саратов: Изд-во Сарат. ун-та, 1986. 78 с.
- Ясюк В.П. Водные моллюски: Определитель, краткий справочник по экологии водных моллюсков Самарской области. Самара, 2005. 92 с.
