Анализ стабильных изотопов углерода и азота в раковинах наземных брюхоногих моллюсков из мезолитических слоев пещеры двойная на Северо-Западном Кавказе

Автор: Антипушина Ж.А., Леонова Е.В., Тиунов А.В.
Журнал: Краткие сообщения Института археологии @ksia-iaran
Рубрика: Биоархеологические исследования
Статья в выпуске: 234, 2014 года.
Бесплатный доступ
В этом отчете представлены данные об изотопном составе органической матрицы оболочки улитки (Helix spp.). Субфоссильные раковины были взяты из мезолитических слоев древостоя Двойная (предгорья Северо-Западного Кавказа, ущелье Губов, Мостовской области). В августе 2012 года и в августе 2013 года были взяты образцы современных уличных раковин улиток. Устойчивый азотный (15N / 14N) и углеродный (13C / 12C) изотопный состав органической матрицы раковины наземной улитки отражает режим осадков. Было высказано предположение, что слои осадка накапливаются в более сухих условиях (затем присутствует). Согласно радиоуглеродным данным, слои, содержащие субфоссильные оболочки, накапливались между 11,8 и 8,3 кал. АД.
Ранний голоцен, стабильные изотопы, археология, каменный век, северо-западный кавказ, реконструкция климата
Короткий адрес: https://sciup.org/14328634
IDR: 14328634
Текст научной статьи Анализ стабильных изотопов углерода и азота в раковинах наземных брюхоногих моллюсков из мезолитических слоев пещеры двойная на Северо-Западном Кавказе
Естественно-научные методы получают все более широкое применение в археологических исследованиях. Одним из методов реконструкции экологических условий прошлого является анализ стабильных изотопов, особенно при исследованиях культурных напластований, насыщенных остатками добычи древнего человека. Анализ стабильных изотопов углерода 13C/12C (обычно выражаемых в виде S13C) и азота 15N/14N (S15N) в коллагене костей из археологических памятников и погребений давно используется для реконструкции диеты. Установлено, что коллаген костей хищников и травоядных животных различается по изотопному составу, так же как и коллаген костей морских и наземных хищников (Kelly, 2000; Hedges et al., 2006). Разный изотопный состав имеют также остатки растительноядных животных, диета которых содержит растения с разным типом фотосинтеза. Согласно исследованиям, ткани растений, имеющих С4 и CAM пути фотосинтеза, более обогащены тяжелым изотопом углерода 13C относительно С3-растений (O’Leary, 1988). Такие растения произрастают преимущественно в засушливых (аридных) местообитаниях. Закономерность между изотопным составом растительности, условиями увлажнения и содержанием δ13С в раковинах выявлена и для наземных брюхоногих моллюсков. Так, в более засушливых условиях в их рационе преобладают С4-растения, в результате чего происходит обогащение раковин 813С. А в более влажных условиях, наоборот, обеднение (Googfriend, 1988; Googfriend, Ellis, 2000). Такая же закономерность установлена между количеством осадков и содержанием δ15N в тканях растений - в более засушливых условиях содержание S15N увеличивается, а в более влажных уменьшается (Swap et al., 2004; Hartman, Danin, 2010). Таким образом, данный метод позволяет реконструировать динамику режима увлажненности.
Цель данной работы заключается в реконструкции динамики режима увлажненности на основе анализа стабильных изотопов углерода и азота в раковинах наземных брюхоногих моллюсков из мезолитических слоев пещеры Двойная.
Описание памятника
Пещера Двойная находится в 9 км к западу-юго-западу от станицы Барака-евской Мостовского района Краснодарского края и расположена на левом берегу р. Губс (левый приток р. Ходзь), на высоте 46 м над уровнем воды в реке (803 м над у. м.). Пещера состоит из двух соседних полостей - малой (восточной) и большой (западной), не соединяющихся между собой. Пещера открыта П. У Аутлевым в 1962 г. ( Аутлев , 1962; Аутлев, Любин , 1994. С. 13), им был заложен шурф перед восточным гротом, где были найдены артефакты, относящиеся к каменному веку. Полученные данные не были опубликованы, со временем забыты, коллекции утеряны. В 2007 г. во вновь обнаруженной пещере Е. В. Беляевой был поставлен шурф в центре западного грота, где частично вскрыты культурные отложения эпохи мезолита, залегавшие «in situ» под мощным горизонтом обвала ( Леонова , 2011).
Начиная с 2008 г. раскопки ведутся в западном гроте пещеры Двойная под руководством Е. В. Леоновой. Площадь раскопа составляет примерно 20 м2. К настоящему моменту на памятнике выделено три культурных слоя, имеющих свои структурные особенности и отличия в комплексах каменного инвентаря, хотя полностью толща еще не исследована ( Леонова и др. , 2013).
Выявлена следующая стратиграфия (рис. 1):
-
1. Рыхлая толща современных и, вероятно, позднесредневековых отложений, представляющих собой многочисленные тонкие золистые, углистые, супесчаные прослойки, небольшие линзы глины и извести, отложившиеся в результате неоднократного посещения пещеры, которая, вероятно, использовалась как временное убежище от непогоды охотниками, в качестве загона скота и т. п. Мощность слоев – 0,4–1 м. Под линией входа в пещеру в профилях читаются следы столбовых ямок.
-
2. Темно-коричневый суглинок с единичными расщепленными кремнями, костями животных и раковинами моллюсков Helix spp. Представлен локально: на западном профиле у выхода и на восточном профиле (центральной оси пещеры) отсутствует. Верхняя граница маркируется тонкой прерывистой углистой прослойкой (0,01–0,02 м) с неровными нижней и верхней границами. Мощность слоя – от 0,08 до 0,25. В центральной части пещеры (северный профиль) мощность резко уменьшается до 0,04 м, и слой здесь залегает непосредственно на монолитной глыбе известняка.
-
3. Завал из крупных блоков известняка (ближе к входу в пещеру более мелкие обломки и щебень, в центральной части – крупный монолитный блок) – от 0,2 до 0,95 м. На контакте с подстилающим слоем 4 – более мелкие обломки известняка с коричневым суглинком и большим количеством раковин моллюсков Helix spp.
-
4. Более темный, чем нижележащий слой, коричневый суглинок с щебнем и большим количеством раковин моллюсков Helix spp. и отдельными угольками – 0,1–0,6 м (верхний мезолитический слой). Ближе к выходу зафиксировано несколько линз с раковинами моллюсков Helix spp. мощностью до 0,2 м.
-
5. Более светлый, чем перекрывающий и подстилающий слои коричневого суглинка, слой со щебнем и большим числом осколков костей и костного тлена, отдельными угольками, с тончайшими углисто-золистыми и белесыми (разложившийся известняк?) прослойками. Общая мощность слоя – от 0,05 до 0,25 м. Верхняя и нижняя границы частично маркированы небольшими углистыми прослойками мощностью до 0,01–0,02 м.
-
6. Слой серо-коричневого суглинка со щебнем, отдельными угольками, раковины Helix spp. немногочисленны, в центральной части у выхода слой 6 пока не обнаружен. Максимальная мощность слоя – до 0,25 м. От нижележащего слоя отделен тонкой прослойкой мелкого щебня, в северной части раскопа эта прослойка превращается в белесый суглинок (разложившийся известняк?), а южнее, ближе к выходу, залегают крупные обломки известняка.
-
7. Серо-коричневый тонкослоистый суглинок, чередуются более светлые и более темные прослойки. Видимая мощность слоя – до 0,3 м.
Верхний (первый) культурный позднемезолитический слой ассоциируется с четвертым и пятым литологическими слоями, второй раннемезолитический слой сопряжен с шестым литологическим слоем, третий культурный верхнепалеолитический слой соотносится с седьмым литологическим слоем. Ориентировочно позднемезолитический слой.
Наибольшее количество раковин брюхоногих моллюсков Helix spp . залегает в нижней части слоя 3 среди обломочного материала и в слое 4, где зафикси-
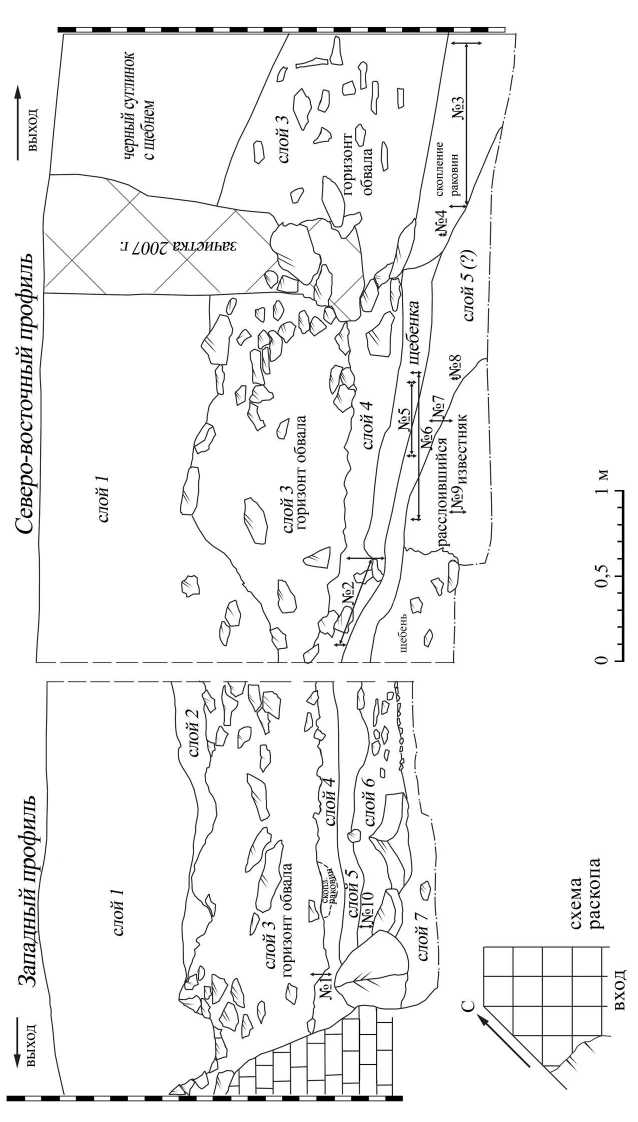
Рис. 1. Профиль раскопа в пещере Двойная с проекциями глубин отбора образцов рованы в привходовой части пещеры скопления раковин моллюсков Helix spp., некоторые из них насчитывали более 5 000 раковин на площади 1,5 м2. В слоях 5 и 6 количество раковин немного уменьшается, а в слое 7 было обнаружено всего 3 раковины моллюсков Helix spp. Ориентировочно возраст слоев может быть намечен несколькими радиоуглеродными датировками. Из отложений, залегающих сразу под слоем обвала, было получено три даты в пределах 10,2–8,3 тыс. лет назад (Леонова, Александрова, 2012. С. 26), указать в настоящий момент конкретные слои, согласно приведенной стратиграфии, затруднительно, поскольку образцы были отобраны из шурфа, заложенного в центре пещеры Е. В. Беляевой в 2007 г., который пока не соединен с раскопом. Для второго мезолитического слоя (слой 6) получено три даты в диапазоне от 11,8 до 8,3 тыс. лет назад (Леонова и др., 2013. Рис. 1).
Материал и методы
Субфоссильные раковины Helix spp. были отобраны из литологических слоев 3–6 пещеры Двойная. Раковины собирались в процессе последовательных зачисток раскопа поквадратно, мощность снятий не превышала 2–3 см, но диапазон глубин залегания в пределах одного образца составляет до 30, что связано с естественными перепадами древних поверхностей (рис. 1). Для сравнения в непосредственной близости от памятника были собраны раковины современных виноградных улиток в августе 2012 и 2013 гг.
Для изотопного анализа была выделена органическая часть раковин, для одной пробы отбиралось по 20–25 раковин. Всего было исследовано 10 мезолитических и 2 современных образца.
Раковины были очищены от загрязнений в дистиллированной воде с использованием мягкой щетки. Выделение белка проводилось по модифицированной методике Лонджина ( Longin , 1971) и ДеНиро ( DeNiro, Epstein , 1978). Для выделения органического матрикса раковины выдерживали в 1М-растворе соляной кислоты НСl при 4 °С до полного растворения карбоната. Осадок переносили в пластиковую пробирку (объемом 50 мл) и отмывали от кислоты дистиллированной водой до pH 4,6–4,7. Затем к осадку добавляли 25 мл раствора соляной кислоты НСl (с pH 2,5) и выдерживали при 85 °С в течение суток. После центрифугирования жидкость с растворенным белком аккуратно сливали в стеклянные пробирки и помещали в термостат до полного высыхания. Навески сухого белка были сделаны в тройной повторности; средний вес навески составил 1500 мкг. Анализ стабильных изотопов углерода и азота проводили на масс-спектрометре Thermo Finnigan Delta V Plus на IRMS в сочетании с элементным анализатором Thermo Flash 1112 (ИПЭЭ РАН, Москва). Содержание изотопов углерода и азота выражено в δ (‰) и представляет собой отклонение от международного стандарта (атмосферный азот или VPDB):
SX (%) = [(R1бразец/Rстaндaрт) - 1] X 1000, где Х – это элемент (азот или углерод), а R представляет собой отношение более тяжелого изотопа к более легкому. Анализ образцов проводился относи- тельно референтного газа (N2 и CO2), откалиброванного относительно стандартов МАГАТЭ (глутаминовая кислота USGS 40, δ15N = –4,5; δ13C = –26,389 и глутаминовая кислота USGS 41, δ15N = +47,6; δ13C = +37,626). В качестве рабочего лабораторного стандарта использовали ацетанилид (δ15N = –4,75; δ13C = –26,78). Стандартное отклонение δ13C и δ15N в USGS 40 (N = 8) составило менее 0,2‰. Во всех образцах было определено изотопное соотношение 13C/12C и 15N/14N, а также атомное соотношение C/N.
Результаты и обсуждение
Согласно исследованиям ДеНиро, посвященным изучению степени сохранности белка в костях из археологических памятников ( DeNiro , 1985), белок хорошей сохранности имеет атомное соотношение C/N в интервале от 2,9 до 3,6. Атомное соотношение C/N наших образцов варьировало в диапазоне 3,2–6,7 для субфоссильных раковин. Атомное соотношение C/N в белке из современных раковин, собранных в августе 2012 г., составило 4,9 (табл. 1), что отражает не очень хорошую сохранность белка. Это можно объяснить тем, что в тот полевой сезон были собраны посмертные раковины моллюсков. В августе 2013 г. удалось обнаружить одного живого моллюска. Атомное соотношение C/N в белке из этой раковины составило 3,3.
Согласно результатам изотопного анализа, все древние образцы раковин моллюсков более обогащены тяжелым изотопом углерода 13С, чем раковины современных моллюсков (рис. 2). Наибольшее количество δ13С зафиксировано в образцах № 3 (скопление раковин при входе в пещеру), 5 (низ десквама-ционного горизонта между слоями 4 и 5 и верхняя часть слоя 5) и 8 (нижняя часть слоя 5): – 19,43‰, – 19,4‰ и – 19,33‰ соответственно. Самым облегченным по δ13С из субфоссильных образцов оказался образец № 6 (22,66‰), отобранный из слоя 5.
Образец № 3 обозначен на графике, но не включен в общую кривую, так как сложно достоверно определить стратиграфическую позицию раковин из скопления при входе в пещеру уже за капельной линией. Скопление (полностью не раскрыто) представляет собой линзу (видимая мощность до 40 см), состоящую из светло-коричневого суглинка, щебня и раковин моллюсков Helix spp., без внутренней стратификации. На наш взгляд, оно образовалось в результате периодической эвакуации мусора из пещеры и\или частичного переотложения культурных остатков в результате эрозии. Последний фактор мог привести к обратной стратиграфии в пределах линзы, когда более древние отложения перекрывают более молодые.
В целом схожий график получился по данным содержания δ15N. Наименее обогащенные изотопом азота δ15N оказались современные образцы (2,95‰ и 5,3‰), а также образцы № 2 (низ слоя 4), 6 (слой 5) и 9 (низ слоя 5?): 4,12‰, 3,38‰, 4,24‰ соответственно (рис. 3). Максимальное содержание изотопа азота δ15N зафиксировано в образце № 8 (14,65‰), отобранном в нижней части слоя 5.
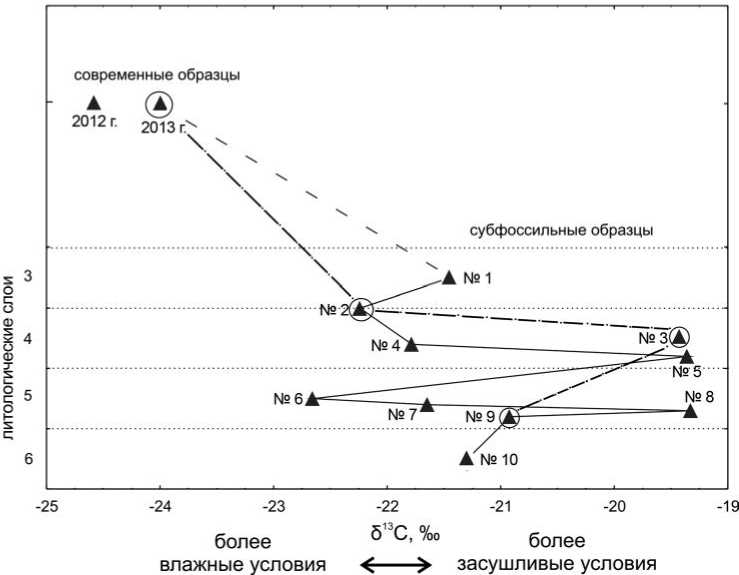
Рис. 2. Динамика содержания δ13С в органическом матриксе раковин субфоссильных и современных виноградных улиток
Как отмечалось выше, атомное соотношение C/N у семи из десяти субфос-сильных образцов было выше 3,6 (табл. 1). Для полноты картины на графиках мы приводим все полученные данные, параллельно построив две кривые: одну, включающую данные всех образов, и другую с хорошей сохранностью белка. В целом общая динамика колебаний режима увлажненности, реконструируемая по всей совокупности данных и по образцам с атомным соотношением C/N в пределах от 2,9 до 3,6, совпадает (рис. 2, 3).
Несмотря на отличие в атомном соотношении C/N (3,3 и 4,9), значения δ13С двух современных образцов очень близки – – 24,01 и – 24,59 ‰ соответственно. Значения δ15N этих современных образцов различались гораздо сильнее (5,30 и 2,95 ‰ соответственно). Мы предположили, что в этом случае динамика δ13С достовернее отражает изменение условий увлажнения исследованного микрорегиона в период 11,8–8,3 тысяч лет назад. Как было сказано выше, существует обратная зависимость между количеством осадков и содержанием δ13С и δ15N. На основе динамики значений δ13C можно предположить, что формирование изученных на данный момент отложений происходило в более засушливых условиях (относительно современных). По количеству осадков Мостовской район относится к зоне избыточного увлажнения, по данным European Climate Assessment and Dataset ( http://eca.knmi.nl ; Klein Tank et al. , 2002),
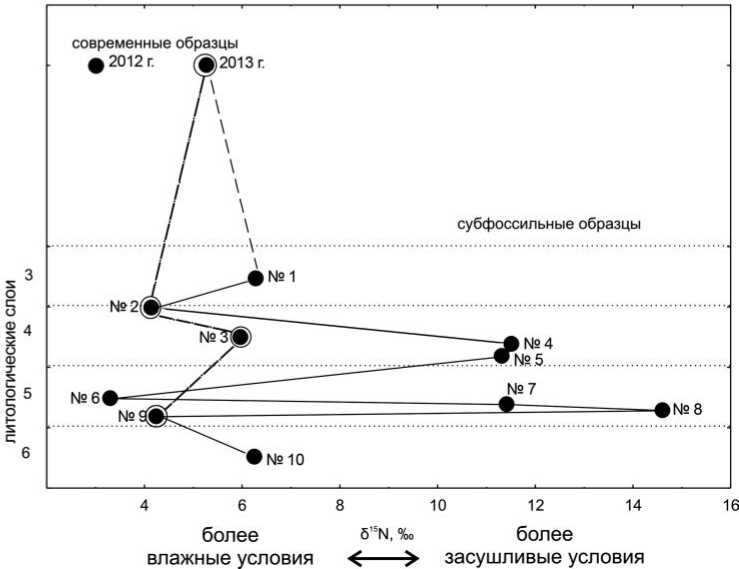
Рис. 3. Динамика содержания δ15N в органическом матриксе раковин субфоссильных и современных виноградных улиток
Таблица 1. Содержание стабильных изотопов углерода и азота в раковинах виноградных улиток (образцы приведены в хронологической последовательности)
При этом формирование 4-го и 5-го слоев происходило при несколько большем количестве осадков, а наиболее засушливые условия фиксируются на уровне контакта четвертого и пятого, а также пятого и шестого слоев.
Отчасти полученные данные подтверждаются стратиграфией. Реконструируемые на основе изотопного анализа наиболее засушливые условия сопряжены с горизонтами десквамации (образцы № 1, 5, 9 (рис. 1)).
Таким образом, анализ стабильных изотопов позволил наметить несколько осцилляций влажностного режима в раннем голоцене в предгорьях Северо-Западного Кавказа на протяжении примерно трех тысяч лет. Применение этого вида анализа представляется перспективным для изучения материалов первобытности и реконструкции природной среды. Накопление новых данных позволит скорректировать полученные результаты, а расширение географических рамок – проследить более общие закономерности взаимосвязи развития и смены культур на фоне климатических изменений.
Список литературы Анализ стабильных изотопов углерода и азота в раковинах наземных брюхоногих моллюсков из мезолитических слоев пещеры двойная на Северо-Западном Кавказе
- Александрова О.И., Антипушина Ж.А., Чернышева Е.В., 2013. Комплексные исследования пещеры Двойная: предварительные результаты//Новые материалы и методы археологического исследования. Материалы II Междунар. конф. молодых ученых/Ред. В.Е. Родинкова., А.Н. Федорина. М. С. 31-32.
- Аутлев А.У., 1962. Отчет об археологических экспедициях, проведенных Адыгейским научно-исследовательским институтом в 1962 году//Архив ИА РАН. Р-I. № 2467.
- Аутлев П.У., Любин В.П., 1994. История исследования палеолита Губского бассейна//Неандертальцы Гупсского ущелья на Северном Кавказе. Майкоп. С. 12-21.
- Леонова Е.В., 2009. К вопросу о хронологии и периодизации позднеплейстоценовых -раннеголоценовых памятников Северо-Западного Кавказа (по материалам последних исследований в Губском ущелье)//Российская археология. № 4. С. 94-107.
- Леонова Е.В., 2011. Раскопки памятника каменного века навес Чыгай и археологические исследования на р. Губс в 2007 г.//АО 2007. М. С. 291-292.
- Леонова Е.В., Александрова О.И., 2012. Динамика культурных процессов в верхнем палеолите -мезолите Северо-Западного Кавказа (по материалам многослойных памятников навес Чыгай и пещера Двойная)//«Историко-культурное наследие и духовные ценности России». Программа фундаментальных исследований Президиума РАН. М.: РОССПЭН. С. 21-28.
- Леонова Е.В., Александрова О.И., Антипушина Ж.А., Сердюк Н.В., Спиридонова Е.А., Тесаков А.С., 2013. Комплексные исследования многослойных памятников каменного века в Губском ущелье//Фундаментальные проблемы квартера, итоги изучения и основные направления дальнейших исследований. VIII Всероссийское совещание по изучению четвертичного периода. Ростов-на-Дону. С. 373-375.
- DeNiro M.J., 1985. Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction//Nature. Vol. 317. P 806-809.
- DeNiro M.J., Epstein S., 1978. Influence of diet on the distribution of carbon isotopes in animals//Geochimica et Cosmochimica Acta. Vol. 42. P. 495-506.
- European Climate Assessment and Dataset (http://eca.knmi.nl)
- Goodfriend G.A., 1988. Mid-Holocene rainfall in the Negev Desert from 13C of land snail shell organic matter//Nature. Vol. 333. P. 757-760.
- Goodfriend G.A., Ellis G.L., 2000. Stable carbon isotope record of middle to late Holocene climate changes from land snail shells at Hinds Cave, Texas//Quaternary International. Vol. 67. P. 47-60.
- Hartman G., Danin A., 2010. Isotopic values of plants in relation to water availability in the Eastern Mediterranean region//Oecologia. Vol. 162. P. 837-852.
- Hedges R.E.M., Stevens Rh.E., Koch P.L., 2006. Isotopes in bones and teeth//Isotopes in palaeoenvironmental research. Dordrecht. P. 117-146.
- Kelly J.F., 2000. Stable isotopes of carbon and nitrogen in the study of avian and mammalian trophic ecology//Canadian Journal of Zoology. Vol. 78. P. 1-27.
- Klein Tank A.M.G., Wijngaard J.B., Können G.P., Böhm R., Demarée G., Gocheva A., Mileta M., Pashiardis S., Hejkrlik L., Kern-Hansen C., Heino R., Bessemoulin P., Müller-Westermeier G., Tzanakou M., Szalai S., Pálsdóttir T., Fitzgerald D., Rubin S., Capaldo M., Maugeri M., Leitass A., Bukantis A., Aberfeld R., van Engelen A.F.V., Forland E., Mietus M., Coelho F., Mares C., Razuvaev V., Nieplova E., Cegnar T., López J. Antonio, Dahlström B., Moberg A., Kirchhofer W., Ceylan A., Pachaliuk O., Alexander L.V., Petrovic P., 2002. Daily dataset of 20th-century surface air temperature and precipitation series for the European climate assessment // International Journal of Climatology. Vol. 22. P. 1441-1453.
- Longin R., 1971. New method of collagen extraction for radiocarbon dating//Nature. Vol. 230. P. 241-242.
- O’Leary M.H., 1988. Carbon isotopes in photosynthesis. Fractionation techniques may reveal new aspects of carbon dynamics in plants//BioScience. Vol. 38. P. 328-336.
- Swap R.J., Aranibar J.N., Dowty P.R., Gilhooly W.P., Macko S.A., 2004. Natural abundance of 13C and 15N in C3 and C4 vegetation of southern Africa: patterns and implications//Global Change Biology. Vol. 10. P. 350-358.
