Анализ видового богатства и ординация сообществ вырубок светло-хвойных зеленомошных лесов Южного Урала

Автор: Кунафин Азамат Мажитович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Структура и динамика растительных сообществ
Статья в выпуске: 1-5 т.14, 2012 года.
Бесплатный доступ
Изучены восстановительные сукцессии в чернично-зеленомошных лесах союза Dicrano-Pinion. Использовались методы ординации и анализа динамики видового богатства и объема ценофлор. Выявлен сложный тренд изменения богатства и возобновления вторичных пород вследствие развития процесса олуговения напочвенного покрова, ингибирующего ход сукцессий.
Вторичные леса, военное нападение, союз dicrano-pinion, фитосоциологический спектр, южный урал
Короткий адрес: https://sciup.org/148200809
IDR: 148200809 | УДК: 581.55
The analyses of the species richness and ordination of clear-cuts communities ligtconiferous boreal forests of the South Ural
The restoration successions after the clear-cutting of bilberry-green moss light-coniferous forests of alliance Dicrano-Pinion are studied. Оrdination method and analysis of species richness and coenflora volume are used. Revealed a complex change in the trend of richness and restocking of secondary forests due to changes in moisture regime of ground cover, inhibiting the progress of succession.
Текст научной статьи Анализ видового богатства и ординация сообществ вырубок светло-хвойных зеленомошных лесов Южного Урала
Изучение восстановительных сукцессий после сплошных рубок является одним из актуальных вопросов науки о растительности. Однако лишь небольшая часть работ посвящена анализу изменения флористического состава серийных сообществ [3, 4, 11, 13].
В статье представлены результаты ординации и анализа видового богатства сообществ, сформировавшихся после сплошных рубок черничнозеленомошных лесов союза Dicrano-Pinion в центрально-возвышенной части Южного Урала (Белорецкий район Республики Башкортостан).
В схеме геоботанического районирования эта территория находится в пределах Авзяно-Белорецкого прибельского центрально-возвышенного района сосновых, мелколиственных лесов и крупнотравных лугов Белорецко-Субхангуловского центрально-возвышенного округа, входящего в Южно-Уральскую горную провинцию [1].
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для настоящей рабо ты послужили 81 полных геоботанических описаний. Размер геоботанических площадок составлял 100-400 м2 в зависимости от однородности контура. Описания площадок и дальнейшая их обработка выполнялись по методике Браун-Бланке [9, 17]. Для обработки материалов использовались база данных TURBOVEG [14], программы JUICE [16] и CANOCO 4.5 [15]. Возраст молодых вырубок определялся по материалам лесоустройства, а более поздних – по возрасту фонового древостоя в соответствии со стандартной дендрохронологической методикой [2]. После проведения процедуры классификации был проведен расчет показателей альфа-разнообразия, ценофлор и построены фитосоцио-логические спектры [8].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Сообщества сплошных вырубок относятся к ассоциации Pleurospermo uralensis-Pinetum sylvestris Martynenko et al. 2003 союза Dicrano-Pinion класса
Vaccinio-Piceetea [5]. По давности рубок было выявлено 6 стадий восстановительной сукцессии. Результаты анализа взаимосвязи вырубок с характерными фитоценотическими показателями с использованием ординационного анализа представлен на рис.1. Стрелками указано ожидаемое изменение значений переменных сред. Острый угол между стрелками свидетельствует о положительной корреляции между факторами, тупой угол – об отрицательно корреляции. При анализе имеющегося массива данных выявлена отрицательная корреляция между проективным покрытием травяного яруса (CH) и проективным покрытием напочвенных мхов (CM), древесных ярусов (Ct1, Ct2, Ct3) и временем рубок (Age).
В правой нижней части диаграммы сгруппировались спелые условно-коренные черничнозеленомошные сосновые леса субассоциации Pleu-rospermo-Pinetum molinietosum caerulea subass. nov.prov. В верхней части диаграммы сгруппировались сообщества на различных стадиях восстановительной сукцессии (группы 2, 3, 5-7), поскольку по видовому составу эти сообщества схожи между собой. Изменяется лишь ценотическая роль (проективное покрытие) типичных таежных зеленых мхов (Pleurozium schreberi, Hylocomium splendens, Di-cranum scoparium, D. polycetum, Ptilium crista-castrensis, Hylocomiastrum umbratum), блока бореальных (Vaccinium myrtillus,Vaccinium vitis-idaea, Lycopodium annotinum, Orthilia secunda, и др.), ге-мибореальных (Calamagrostis arundinacea, Brachy-podium pinnatum, Carex rhizina и др.) и древесных (Pinus sylvestris, Betula pendula, B. pubescens) видов. В правой верхней части диаграммы сгруппировались лесные сообщества (группа 7), вырубка которых проводилась постепенно, и потому напочвенный покров и древостой частично сохранились. В левой нижней части диаграммы в сторону увеличения проективного покрытия трав сгруппировались влажные злаково-широкотравные сообщества с очень плотным и высоким травостоем (группа 4). Данная стадия сукцессии является ингибирующей, поскольку семена древесных пород не могут прорасти сквозь плотный полог травостоя и может существовать десятки лет пока не изменится почвен- но-гидрологический режим в результате воздействия какого-либо экологического фактора.
Был проведен анализ характеристик видового богатства разных стадий сукцессий (табл. 1) и их фито-социологических спектров (рис. 2), эффективность использования которых была показана в ряде работ [4, 6, 7, 12, 13].
Из табл. 1 видно, что в первые 1-3 года после вырубки происходит обеднение видового состава на вырубках (синтаксоны 2, 3) по сравнению с коренным типом леса (синтаксон 1), что связано с выпадением (выгоранием) из напочвенного покро- ва бореальных видов и исчезновения редких видов в связи с резким осветлением и нарушением целостности почвенного покрова во время проведения рубок. Однако при снижении доли участия видов класса Vaccinio-Piceetea (рис. 2, стадии 2, 3), в сложении этих сообществ возрастает доля участия видов светлохвойных лесов класса Brachypodio-Betuletea, субальпийского высокотравья класса Mulgedio-Aconitetea и сибирского широкотравья порядка Abietetalia sibiricae, а также луговых видов класса Molinio-Arrhenatheretea.
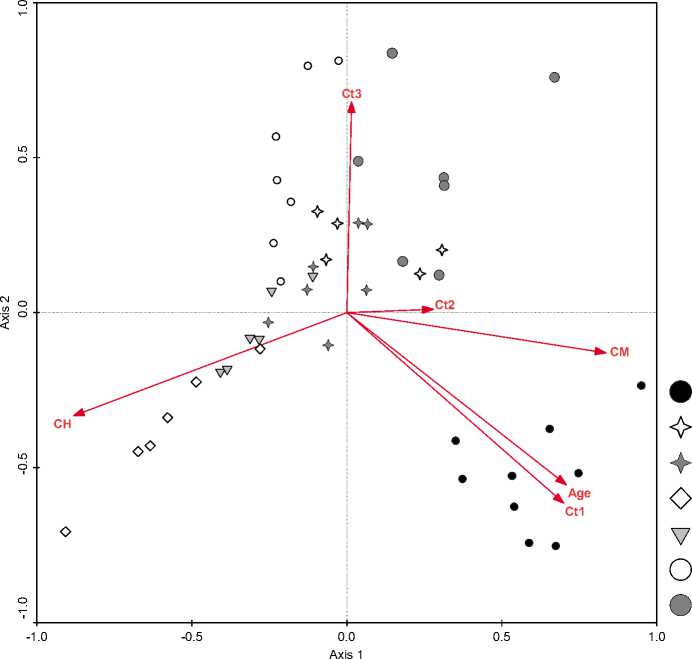
Рис. 1 Ординация (CCA) вырубок на различной стадии восстановления в связи с фитоценотическими показателями и возрастом вырубок. Проекции на две оси максимального варьирования.
Стадии сукцессии по давности рубок: 1 – > 110 лет , 2 – 1 год, 3 – 3 года, 4 – ингибирующая стадия (5-20 лет), 5 – 8 лет, 6 – 15 лет, 7 – 15-20 лет. Факторы: Age – время вырубки; проективное покрытие (древостоя – Ct1 – 1 ярус, Ct2 – 2 подъярус, Ct3 – 3 подъярус), травостоя (CH), напочвенных мхов (CM).
Таблица 1 Флористическое разнообразие сообществ вырубок на различных стадиях восстановления
|
Показатели биоразнообразия |
Синтаксоны |
|||||||
|
1 \ |
2 \ |
3 \ |
4 \ |
5 \ |
6 \ |
7 |
||
|
Альфа-разнообразие |
||||||||
|
Общее |
среднее |
53,9 |
49,5 |
45,8 |
74,6 |
55,0 |
55,7 |
53,9 |
|
варьирование |
46-64 |
41-57 |
37-56 |
70-81 |
52-61 |
52-62 |
47-59 |
|
|
Высших сосудистых |
среднее |
41,7 |
37,2 |
34,1 |
57,0 |
40,0 |
42,0 |
41,9 |
|
варьирование |
33-47 |
30-46 |
26-41 |
53-61 |
36-45 |
35-49 |
32-56 |
|
|
Объем ц |
енофлоры |
|||||||
|
Общее |
90 |
91 |
74 |
112 |
89 |
91 |
112 |
|
|
Высших сосудистых |
84 |
85 |
69 |
109 |
84 |
87 |
106 |
|
Примечание . Стадии сукцессии по давности рубок: 1 – > 110 лет , 2 – 1 год, 3 – 3 года, 4 – ингибирующая стадия (5-20 лет), 5 – 8 лет, 6 – 15 лет, 7 – 15-20 лет.
К 5-му году на вырубках, приуроченных преимущественно к выровненным участкам в нижних частях склонов, где нет эрозии почв и происходит накопления влаги, наблюдается сильное олугове-ние, о чем свидетельствуют высокие показатели альфа-разнообразия. В результате, в сообществах на этой стадии сукцессии происходит увеличение видового разнообразия за счет усиления ценотиче-ских позиций видов влажных лугов класса Molinio-Arrhenatheretea и опушечных видов класса Trifo-lio-Geranietea . Эта стадия ингибирующего луга длится неопределенный срок (данная стадия автором зафиксирована на вырубках сроком давности от 5 до 20 лет). Таким образом, происходит процесс торможения сукцессии [10], при этом луговые и опушечные виды сильно угнетают развитие не только бореальных видов , но и всего древесного подроста (табл.1 синтаксон 4, рис. 2 стадия 4).
На 8-ми и 15-ти летних вырубках (таб. 1 синтак-соны 5, 6), не затронутых стадией ингибирующего луга, идет процесс активного разрастания вторичных лесообразователей (Betula pendula, B. pubes-cens) и внедрения в эти сообщества светолюбивых гемибореальных видов класса Brachypodio-Betuletea, луговых и широкотравных видов классов Molinio-Arrhenatheretea и порядка Abietetalia si-biricae (рис. 2, стадии 5, 6), за счет которых видовая насыщенность возрастает (табл. 1, синтаксоны 5, 6), по сравнению с вырубками 1-3 лет, но при этом доля участия видов класса Vaccinio-Piceetea сохраняется примерно на том же уровне.
В сообществах на вырубках 15-20 лет, рубка которых проводилась постепенно, напочвенный покров и древостой достаточно хорошо сохранились , показатели видовой насыщенности незначительно снижаются (табл.1, синтаксон 7). В фитосоциоло-гическом спектре (рис.2, стадия 7) также отмечено повышение доли участия видов гемибореальных светлохвойных лесов класса Brachypodio-Betuletea , усиление ценотических позиций видов лугов и сибирского широкотравья классов Molinio-Arrhenatheretea, Mulgedio-Aconitetea и порядка Abietetalia sibiricae и небольшое снижение доли участия видов класса Vaccinio-Piceetea , что объясняется постепенным осветлением древесного полога и более щадящим методом рубки.
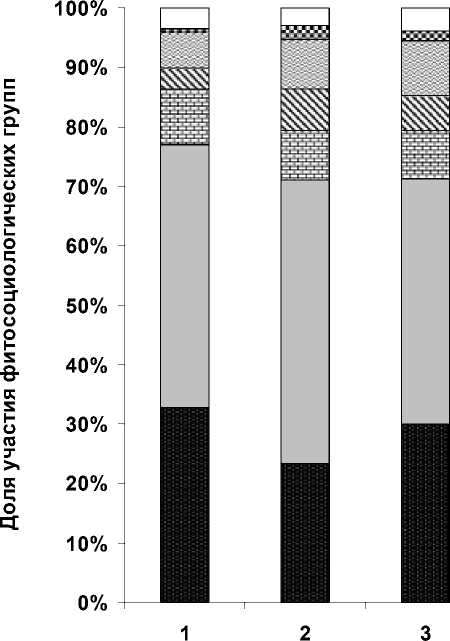
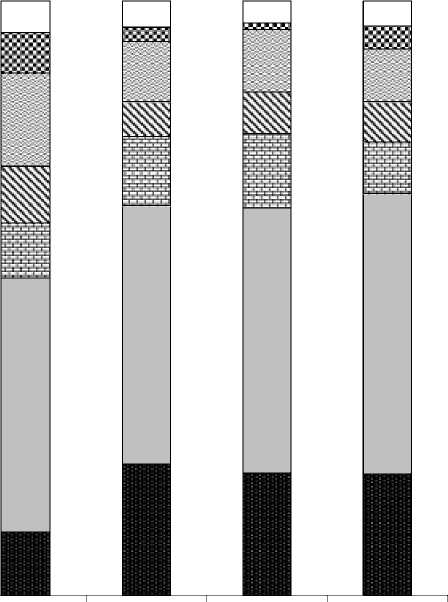
V-P B-B Q-F Mul-A+ABIET SIB M-A TR-G+F-B Прочие
Рис. 2. Фитосоциологический спектр сообществ вырубок на различных стадиях восстановления
Примечание. V- P - виды класса Vaccinio-Piceetea, B-B - виды класса Brachypodio-Betuletea, Q-F - виды класса Querco-Fagetea , Mul-A + Ab sib - субальпийское высокотравье класса Mulgedio-Aconitetea и порядка Abietetalia sibiricae, M-A - виды класса Molinio-Arrhenatheretea , T-G+F-B - виды классов Trifolio-Geranietea и Festuco-Brometea . Стадии сукцессии по давности рубок: 1 - > 110 лет , 2 - 1 год, 3 - 3 года, 4 - ингибирующая стадия (5-20 лет), 5 - 8 лет, 6 - 15 лет, 7 - 15-20 лет.
Таким образом, на выявленных ранних стадиях восстановительной сукцессии после сплошных рубок в сообществах ассоциации Pleurospermo uralen-sis-Pinetum sylvestris наблюдается закономерное снижение, а затем увеличение видового богатства за счет изменения обилия и ценотической роли видов классов Vaccinio-Piceetea, Brachypodio-Betuletea, Molinio-Arrhenatheretea, Trifolio-Geranietea и активного возобновления вторичных лесообразовате-лей. Исключение составляет стадия ингибирующего луга, в сообществах которой резко возрастает видовое богатство, усиливается роль видов влажных лугов и опушек классов Molinio-Arrhenatheretea и Tri-folio-Geranietea и может длиться неопределенное время [10].
Работа поддержана грантами РФФИ № 10-04-00534-а и № 11-04-10138-к.
Список литературы Анализ видового богатства и ординация сообществ вырубок светло-хвойных зеленомошных лесов Южного Урала
- Жудова П.П. Геоботаническое районирование Башкирской АССР. Уфа, 1966. 123 с.
- Корчагин А.А. Определение возраста деревьев умеренных широт//Полевая геоботаника. Т. II. М.;Л.: Изд. АН СССР. 1960. С. 209-241.
- Крышень А.М. Растительные сообщества вырубок Карелии. М.; Наука, 2006. 262 с.
- Кунафин А.М., Широких П.С., Мартыненко В.Б. Оценка эффективности восстановительной сукцессии после рубок с использованием фитосоциологических спектров//Известия Самарского НЦ РАН. 2011. Т. 13. № 5(2). С. 86-89.
- Мартыненко В.Б. Синтаксономия лесов Южного Урала как теоретическая основа развития системы их охраны: Дис.. докт. биол. наук. Уфа, 2009. 495 с.
- Мартыненко В.Б., Миркин Б.М. О формальных и неформальных оценках флористического разнообразия (на примере сосняков Южного Урала)//Экология. 2003. № 5. С. 336-340.
- Миркин Б.М., Мартыненко В.Б., Широких П.С., Наумова Л.Г. Анализ факторов, определяющих видовое богатство сообществ лесов Южного Урала//Журнал общей биологии. 2010. Т. 71. № 2. С. 131-143
- Миркин Б.М., Наумова Л.Г. Наука о растительности (история и современное состояние основных концепций). Уфа: Гилем, 1998. 413 с.
- Миркин Б.М., Наумова Л.Г., Соломещ А.И. Современная наука о растительности. М.: Логос. 2000. 264 с.
- Смирнова О.В., Торопова Н.А., Луговая Д.Л., Алейников А.А. Методология исследования популяционной организации и сукцессионной динамики лесных экосистем (биогеоценозов)//Методические подходы к экологической оценке лесного покрова в бассейне малой реки./Л.Б. Заугольнова, Т.Ю. Браславская (отв. ред.). М.: Товарищество научных изданий КМК. 2010. С. 20-40.
- Уланова Н.Г. Восстановительная динамика растительности сплошных вырубок и массовых ветровалов в ельниках южной тайги (на примере европейской части России). Автореф. дис.. доктора биол. наук. М., 2006. 46 с.
- Широких П.С., Мартыненко В.Б. Закономерности изменения фиторазнообразия лесов в синтаксономическом пространстве//Флора и растительность Южно-Уральского государственного природного заповедника/Кол. авторов. Под ред. Б.М. Миркина. Уфа: Гилем, 2008. C. 241-255.
- Широких П.С., Мартыненко В.Б., Кунафин А.М., Миркин Б.М. Особенности флористического состава некоторых типов вторичных лесов Южно-Уральского региона//Бюллетень МОИП. Отделение биологическое. 2012. Т. 117, Вып. 2. С. 42-54.
- Hennekens S.M. TURBO(VEG). Software package for input, processing, and presentation of phytosociological data. User’s guide. IBN-DLO, University of Lancaster, Lancaster, 1996. 59 p.
- Ter Braak, C. J. F. & Šmilauer P. Reference manual and CanoDraw for Windows User’s guide: Software for Canonical Commuhity Ordination (version 4.5). Microcomputer Power. Ithaca, NY, USA. 2002. P. 500.
- Tichý L., Holt J., Nejezchlebová M. 2011. JUICE. Program for management, analysis and classification of ecological data. 2nd edition. Brno: Masaryk university. 61 p.
- Westhoff V., Maarel E. van der. The Braun-Blanquet approach//Classification of plant communities/Ed. R.H.Whittaker. The Hague. 1978. P.287-399.
