Анализ влияния запасов корма на динамику численности популяции белки (на примере Еврейской автономной области)
")
Автор: Ревуцкая О.Л.
Журнал: Региональные проблемы @regionalnye-problemy
Рубрика: Региональный прогноз. Математическое моделирование
Статья в выпуске: 2 т.13, 2010 года.
Бесплатный доступ
Проведен анализ влияния запасов корма на изменение численности популяции белки (на примере Еврейской автономной области). Для описания тенденций динамики численности использован подход, основанный на математическом моделировании. Проведена верификация моделей и оценено современное состояние популяции белки. Показано, что изменение численности популяции определяется урожайностью орехов кедра в годы, предшествующие размножению.
Динамика численности, белка, кормовые запасы, количественный анализ, математическое моделирование
Короткий адрес: https://sciup.org/14328726
IDR: 14328726 | УДК: 51:599.73(571.621)
Analysis of the squirrel population dynamics dependence on the forage reserve (by the example of the Jewish autonomous region)
We have analyzed the forage reserve influence on the squirrel number change (by the example of the Jewish autonomous region). The approach, based on mathematical modeling, is used for the description of tendencies in the population size dynamics. The models verification is carried out and the present day condition of the squirrel population is estimated. It is shown that a change in the squirrel number is determined by the cedar nut yield in the years, prior to reproduction.
Текст научной статьи Анализ влияния запасов корма на динамику численности популяции белки (на примере Еврейской автономной области)
Наличие и доступность кормов являются одними из важнейших характеристик биотопов, влияющих на воспроизводство и пространственно-временную динамику популяций [6, 17]. Хорошие кормовые и защитные условия биотопов и местообитаний способствуют не только увеличению количества животных, но и укрепляют сопротивляемость организма различным заболеваниям, предупреждают их дальние миграции и выходы в сельскохозяйственные угодья, а также концентрируют животных в местах охоты, что важно при ведении охотничьего хозяйства [17]. Изменение кормовых условий является наиболее широко распространенной причиной колебаний численности зверей. Особенно это отражается на видах, питающихся однообразными кормами [12].
Из охотничье-промысловых видов в Еврейской автономной области (ЕАО) одним из наиболее зависимых от кормовых запасов является белка ( Sciurus vulgaris mantchuricus Thomas, 1909) [5, 7, 12]. Особенности питания обусловливают наибольшую концентрацию особей этого вида в кедрово-широколиственных лесах, где основным кормом служат семена сосны (кедра) корейской ( Pinus koraiensis Siebold et Zucc.) [5, 7, 11]. При хорошем урожае орехов кедра плодовитость белок увеличивается; более того, растет их численность за счет миграции зверьков из других районов менее богатых кормами. В неурожайные годы, наоборот, количество белки сокращается в связи с их миграциями, повышенной смертностью от голода и снижением плодовитости [12].
В данной работе продолжено изучение динамики численности охотничье-промысловых популяций ЕАО, в частности белки. В отличие от предыдущих работ [18], в которых были проанализированы данные учета численности белок с 1994 по 2006 гг. и проведено сравнение тенденций их развития в промысловых и свободных от промысла территориях, здесь исследуется динамика популяции с 1994 по 2009 гг. в зависимости от запасов кормовых ресурсов (орехов кедра). Рассматриваются и сопоставляются тенденции изменения количества этих грызунов в лесных угодьях как в целом по области и ее районам, так и в ряде охотничьих хозяйств и на особо охраняемых природных территориях (ООПТ) ЕАО, расположенных в зоне произрастания кедрово-широколиственных лесов.
Для описания тенденций динамики численности использован подход, основанный на математическом моделировании. Для количественного анализа применены методы регрессионного и корреляционного анализа. На основе модифицированной модели Рикера была формализована зависимость популяционных параметров (рождаемости и выживаемости) от величины запасов корма. В данном случае относительно простая модель позволяет получить оценки репродуктивного потенциала популяции и интенсивности влияния кормовых запасов на демографические параметры.
Район работ
В соответствии с геоботаническим районированием Г.Э. Куренцовой [8] основные массивы кедрово-широколиственных лесов ЕАО объединены в границах Сутар-ско-Помпеевского района широколиственно-елово-кедровых лесов и их производных. Сутарско-Помпеевский район расположен на внутренней части горной системы Малого Хингана и включает основные горные хребты Помпеевский, Сутарский, Шухи-Поктой и южную часть хребта Малый Хинган, верховья рр. Биджан, Сутара, Дитур и среднее течение рр. Помпеевка, Дичун, а также левых притоков р. Бира [16].
В состав лесообразующих пород этих лесов входят: кедр корейский, ель аянская, пихта белокорая, липа амурская, орех маньчжурский и другие. Хорошо развит подлесок из лещины маньжурской, чубушника тонколистного, элеуторококка колючего и многих других растений [16]. Таким образом, кедрово-широколиственные леса выделяются большим разнообразием и обилием растительных кормов, являются кормовой базой и местом обитания различных видов промысловых зверей (белок, колонков, изюбрей, кабанов, косуль, медведей, соболей и др.) и птиц [11].
Кедровые леса области произрастают на площади 159,9 тыс. га (10,12 % от общей площади основных лесообразующих пород) [9]. Они расположены преимущественно в Облученском районе, на территории Октябрьского и Биробиджанского районов занимают сравнительно меньшую площадь [8, 9]. Среди древесных пород Еврейской автономии кедр корейский занимает особое положение. Он занесен в Красную книгу ЕАО [9, 14].
Материалы исследования
Оценка параметров моделей проводилась на основе годовых отчетов по зимнему маршрутному учету (ЗМУ) численности популяции белки. Данные ЗМУ анализировались на всей территории ЕАО с 1994 по 2009 гг., Облу-ченского, Октябрьского и Биробиджанского районов, общества охотников и рыболовов ЕАО (ООиР) с 1999 по 2009 гг., общества с ограниченной ответственностью (ООО) «Сутара» с 1993 по 2009 гг., ООО «Ирбис» и общественной региональной организации (ОРО) «Диана» с 2003 по 2009 гг., заказников «Шухи-Поктой» и «Журавлиный» с 1994 по 2009 гг., заповедника «Бастак» с 1992 по 2009 гг. Обратим внимание, что в статистику численности белки в районах области не включены данные заповедника. Фактически в работе были проанализированы все доступные данные учета для тех территорий, на которых произрастает и плодоносит кедр корейский.
Сведения о запасах кедра корейского были предоставлены специалистами Управления лесами Правительства ЕАО. Оценки урожайности кедра экстраполируются как на всю территорию области, так и на отдельные ее районы. Также были использованы оценки урожайности в лесных угодьях ООО «Ирбис».
Кроме того, в работе учтены данные опросов специалистов Управления по охране и использованию объектов животного мира Правительства ЕАО, охотничьих хозяйств (ООиР, «Ирбис») и ООПТ.
Методы исследования и оценки параметров
Обозначим через xn численность популяции в n -ом году. В условиях повышенной интенсивности конкурентных взаимоотношений между особями более адекватной оказывается модель Рикера:
x n +1 = ax n e — bx" , (1) где параметр a – репродуктивный потенциал популяции, то есть скорость годового воспроизводства в отсутствии лимитирования, параметр b определяет характер развития популяции и является мерой влияния плотностной зависимости. Параметр b равен величине обратной численности xn , при которой функция xn+ 1( xn ) – численность в следующем году – достигает максимально возможного значения M ( М = a ( be )-1). Оценки параметров модели (1) позволяют определить равновесное значение численности, которое находится по формуле K=(lna)/b [18, 19].
Заметим, что параметры популяционной модели (1) постоянны во времени, а условия среды, в которой обитает популяция, непрерывно изменяются. Для популяции белки такими внешними условиями, в первую очередь, выступают кормовые запасы, в случае их недостатка, они являются ограничивающим фактором, зависящим от плотности. Поэтому для описания колебаний численности белки модель Рикера была дополнена оценками внешних факторов. В частности, была учтена зависимость функции воспроизводства от обеспеченности популяции кормовыми ресурсами.
Обозначим через Cn-1 и Cn – урожайность ресурсов кедра в смежные годы, тогда коэффициент bn, характеризующий в модели (1) конкурентную способность особей между периодами размножения (n-1)-го и n-го годов, можно представить в виде функции bn=-b0+b1Cn-1+b2Cn. По- добная зависимость коэффициента лимитирования от урожайности корма была вполне успешно использована при моделировании популяции маньчжурской белки [1, 20].
Таким образом, модифицированная модель Рикера, учитывающая влияние интенсивности урожая кормов на выживаемость животных, примет вид
- bx„ xn+1 = axne n .
Модель Рикера с учетом влияния кормовых запасов на рождаемость, имеет вид
-bnx„ + btC„ .+ b-,C„ v -- rt"V О 0 n 1 n—1 2 n xn+1 = axne ,
где произведение aebCn —1 + b 2 Cn фактически является коэффициентом рождаемости (среднее число новорожденных, приходящихся на одну взрослую особь).
Модели 2 и 3 позволяют проследить изменения значений выживаемости и рождаемости соответственно в зависимости от урожайности кормовых запасов.
Для оценки интенсивности реального годового воспроизводства в данный период наблюдений было использовано уравнение линейного роста (модель Мальтуса) X = б ■ x , которое имеет решение (при x ( 0 ) = x0 ): x ( t ) = x 0 ■ e б t . Дискретный аналог этой модели:
xn+1 = a ■ xn содержит параметр a = eб , характеризующий процесс годового воспроизводства, включая миграционный баланс [18].
Модель Рикера (1) можно представить в виде экспоненциальной функции xn/xn+1 от xn. Тогда оценка параметров сводится к вычислению коэффициентов соответствующего регрессионного уравнения и может быть осуществлена с помощью любого доступного статистического пакета. Задача оценки параметров моделей (2) и (3) заключалась в подборе таких значений a, b0, b1, b2, при которых последовательность xn* (модельная численность) наилучшим образом аппроксимирует известную последовательность xn (фактическая численность). Значения xn* определялись по уравнениям (2) и (3) численно методом Квази-Ньютона в программе MathCAD 14. Подбирались наборы указанных параметров, обеспечивающие минимумы величины L = ^(ln xn — ln x* )2 . Значение параметра а в модели Мальтуса (4) можно определить, построив экспоненциальный тренд вида y = rxe±Ex по вре менному ряду данных численности, где a = e±б .
Результаты и обсуждение
Оценки параметров моделей Мальтуса и Рикера и соответствующие значения стационарных и максимальных численностей, полученные для белки на основе учетных данных разных территорий, представлены в табл. 1.
Оценки параметров уравнений (2) и (3) и начальной численности ( x 0) коэффициентов детерминации ( R 2) и
Таблица 1
Оценки численностей и параметры моделей динамики белки, обитающей на территории ЕАО
|
Территория |
Учетная численность в последний год, шт. |
Максимальная учетная численность, шт. |
Модель Мальтуса |
Модель Рикера |
||
|
a |
a |
M, шт. |
K, шт. |
|||
|
ЕАО (1994–2009 гг.) |
23555 |
81393 |
0,9815 |
2,5 |
31050 |
30750 |
|
ЕАО (1999–2009 гг.) |
23555 |
81393 |
1,03 |
2,97 |
74310 |
27235 |
|
Облученский район |
17561 |
57354 |
1,02 |
2,93 |
58550 |
21500 |
|
Октябрьский район |
4392 |
19830 |
1,01 |
3,5 |
17580 |
6300 |
|
Биробиджанский район |
1602 |
3769 |
1,22 |
1,86 |
4650 |
1550 |
|
Заповедник «Бастак» |
485 |
3462 |
1,05 |
2,1 |
2940 |
1031 |
|
ООиР (Облученский район) |
426 |
1154 |
1 |
3,94 |
2320 |
807 |
|
Заказник «Шухи-Поктой» |
1044 |
1949 |
1,09 |
1,44 |
2880 |
730 |
|
ООО «Ирбис» |
3831 |
6624 |
1,13 |
2,53 |
8440 |
3100 |
ошибки аппроксимации ( А ) для фактического и модельного рядов данных приведены в табл. 2.
Обсудим полученные результаты оценки параметров моделей параллельно с анализом закономерностей пространственно-временной динамики численности популяции белки, выявляемых на основе годовых отчетов по учету численности.
Современный ареал обитания белки приурочен к кедрово-широколиственным лесам Малого Хингана, проходящим по охотничьим угодьям Облученского, Октябрьского и Биробиджанского районов. На равнинных пространствах автономии, которые преобладают в ее восточной части, белка малочисленна [5, 7, 10].
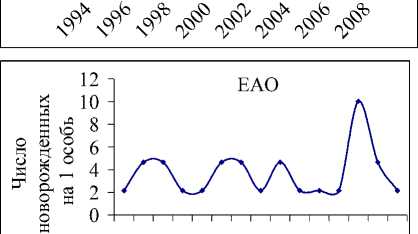
В популяции белки на территории ЕАО с 1994 по 2009 гг. происходило снижение численности, регистрируемое на фоне сильных нерегулярных колебаний (рис. 1а). Однако если рассматривать данные за последние 11 лет, то видно (рис. 1б), что наметилась тенденция увеличения численности популяции (экспоненциальный тренд положителен). Скорее всего, стабилизация происходит в связи с уменьшением промыслового давления (возможно, в силу нерентабельности охоты на белку [13]) и улучшением условий после введенного в 1994 г. запрета на вырубку кедра корейского [9, 14].
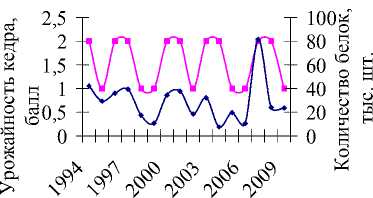
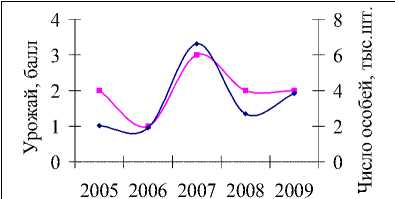
Многолетние данные учета численности и урожайности кедра (с 1994 по 2009 гг.) показывают, что увеличение популяции белки происходит на следующий год пос- ле высоких урожаев орехов (рис. 2а). Например, всплески численности, отмечаемые в 1997, 2000, 2003 и 2007 гг., наблюдались после хороших урожаев семян кедра в предыдущих сезонах размножения. Неурожаи кормов в 1998, 2001, 2004, 2007 гг. (рис. 2,а), наоборот, привели к резкому уменьшению численности в следующем году, поскольку белки мигрировали осенью предыдущего года в поисках хороших кормовых угодий [1–3, 5, 7, 10]. В частности, кормовые миграции белок были установлены в Биробиджанском, Облученском [5] и Октябрьском районах ЕАО.
В 2004 г. наблюдалась самая низкая численность белки за рассматриваемый период (7507 особей, по сравнению с предыдущим годом популяция уменьшилась почти в 3 раза), несмотря на хороший урожай семян кедра в 2003 г. В 2007 г. отмечался всплеск численности белки (увеличение почти в 8 раз), так в 2006 г. она составляла 10439, а в 2007 – уже 81393. Однако для этих двух лет урожай кедра оценивался на 2 балла по 3-балльной шкале, то есть, урожай оценивался как средний. Вместе с тем, резкое снижение и увеличение количества белок, в первую очередь, определяется слабым или особо обильным урожаем кедра [5, 7]. По-видимому, для белок кормовых запасов в 2003 г. было не достаточно, а в 2007 г., наоборот, много. В связи с этим, при верификации моделей (2) и (3) данные урожая кедра были уточнены для этих двух лет, в
Таблица 2
Значения параметров моделей (2) и (3), полученные по данным оценок численности белок и запасов орехов кедра корейского в ЕАО
|
Территория |
a |
b 0 |
b 1 |
b 2 |
x 0 |
A (%) |
R2 |
|
ЕАО (модель 2) |
9,2 |
1.4⋅10 - 4 |
1.2 ⋅10 - 5 |
2.9 ⋅10 - 5 |
27050 |
25,2 |
0,72 |
|
ЕАО (модель 3) |
0,82 |
3.2⋅10 - 5 |
1⋅ 10 - 4 |
0,77 |
42111 |
24,1 |
0,91 |
|
Облученский район (модель 2) |
7,6 |
2⋅10 - 4 |
1.5⋅10 - 5 |
4.7⋅10 - 5 |
8714 |
22,2 |
0,88 |
|
Облученский район (модель 3) |
1 |
6.1⋅10 - 5 |
0,078 |
0,651 |
10380 |
32,3 |
0,95 |
|
Октябрьский район (модель 2) |
9,1 |
6.9⋅10 - 4 |
4.9⋅10 - 5 |
1.4 ⋅10 - 4 |
1597 |
20,5 |
0,82 |
|
Октябрьский район (модель 3) |
1 |
2⋅10 - 4 |
0,025 |
0,717 |
3273 |
32 |
0,89 |
|
ООО «Ирбис» (модель 2) |
3,6 |
9.7⋅10 - 4 |
5.6⋅10 - 6 |
3.1⋅10 - 4 |
1974 |
1,6 |
0,99 |
|
ООО «Ирбис» (модель 3) |
1 |
3.4⋅10 - 4 |
0,028 |
0,61 |
1849 |
2,7 |
0,99 |
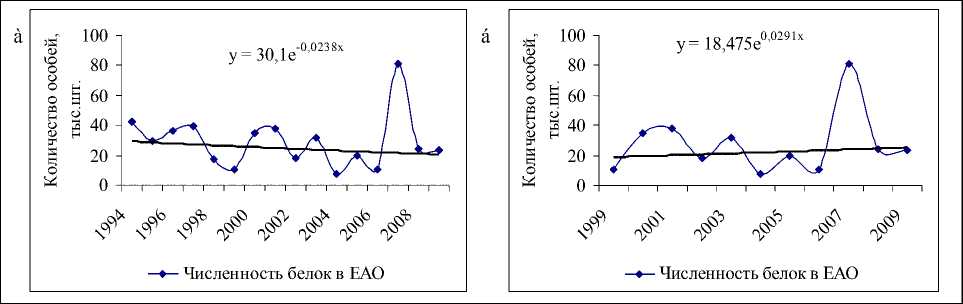
Рис.1. Данные учета численности белки на территории ЕАО и соответствующие экспоненциальные тренды: а) с 1994 по 2009гг.; б) с 1999 по 2009 гг.
2003 г. было решено присвоить урожайности кедра балл 1 (урожай слабый), в 2006 г. – балл 3 (урожай хороший).
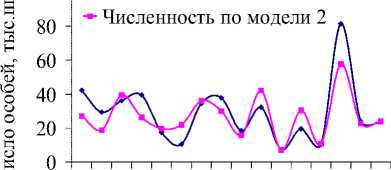
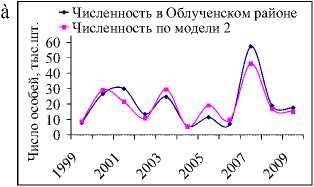
Совпадение тенденций динамики модельных и фактических данных (рис. 2б, 2в) позволяют судить о существенном влиянии запасов кедра на развитие популяции грызунов. Соотношение между величинами b1 и b2 моделей 2 и 3 (табл. 2) показывает, что численность белки определяется главным образом кормовыми условиями предыдущего года. Отклонения графиков, вероятно, вызваны действием неучтенных в моделях факторов. Например, неурожайный на кедровые орехи 1994 г. привел лишь к незначительному снижению численности, возможно, в этот год был урожай других видов корма, благодаря чему не произошло массовое снижение числа белок.
Согласно модифицированной модели Рикера (2), репродуктивный потенциал белок равен 9,2, это означает, что годовая продуктивность одной самки (в отсутствии лимитирования) составляет 18–19 особей. Наряду с этим, в соответствии с оценками модели (3), коэффициент рождаемости варьируется в зависимости от интенсивности плодоношения кедра (рис. 2г). Например, после слабого урожая орехов у одной самки в среднем за год рождается примерно 4–5 детенышей, после среднего – 9–10 и хорошего – 20. При этом выживаемость зверьков изменяется от 0,1 до 0,56. Заметим, что модельные оценки не противоречат данным литературных источников [5, 7, 10, 12], согласно которым перезимовавшие самки за год приносят 1–2 выводка, при этом количество молодых особей в одном выводке колеблется от 2 до 7 (редко до 10).
Примечательно то, что максимальный коэффициент рождаемости, отмечавшийся в 2007 г., в рассматриваемом временном интервале больше, чем скорость годового воспроизводства в отсутствии лимитирования. Вероятно, в год обильного урожая запасы кедра способ-
à
-■- Запасы орехов кедра корейского
-•- Численность белок в ЕАО
á
H 100 — -•- Численность белок в ЕАО
§ 80
R о
S
У
Численность белок в ЕАО
Численность по модели 3
>S
o^ Jo ^
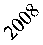
o^ ofe cfr c\V x^ сф ^
-•- Значение коэффициента рождаемости
Рис. 2. Динамика численности белки в ЕАО в зависимости от запасов орехов кедра корейского (ряд данных об урожае кедра смещен на 1 год вперед) (а). Фактическая и модельная численность белки в ЕАО по модели 2 (б) и 3 (в). Динамика коэффициента рождаемости для белок, обитающих в ЕАО (г)
á
Численность белок в Октябрьском районе
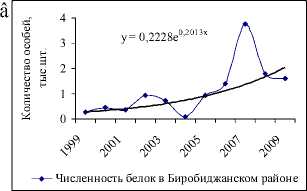
Рис. 3. Данные учета численности белки на территории Облученского (а), Октябрьского (б), Биробиджанского (в) районов с 1999 по 2009 гг. и соответствующие им экспоненциальные тренды ствовали не только максимальному приросту местной популяции белки, но и миграции особей с сопредельных территорий области. Вместе с тем, при среднем урожае кедра коэффициент рождаемости оказывается меньше репродуктивного потенциала чуть ли не в два раза.
Территориальное распределение популяции белки по районам области зависит от размещения типичных для нее биотопов и их кормовых, защитных и гнездовых качеств. Наиболее высокая численность белки в Облучен-ском районе (рис. 3а), лесные угодья которого предоставляют этим зверькам наиболее благоприятные условия существования. Ниже численность данного вида, наблюдается в Октябрьском районе (рис.3б), еще ниже – в Биробиджанском (рис.3в).
Статистический анализ учетных данных (а именно, определение средних коэффициентов эластичности) показал, что увеличение численности на 1 % от ее среднего уровня в Облученском районе приводит к увеличению на 0,73 % от среднего уровня количества белок в ЕАО, в Октябрьском районе – на 0,23 %, а в Биробиджанском – всего на 0,04 %.
В исследуемый период на территории этих районов наблюдается тенденция роста численности белки на фоне выраженных колебаний вокруг равновесного уровня, оцениваемого в Облученском районе около 21500 особей, Октябрьском – 6300 и Биробиджанском – 1550 (табл. 1). Одновременно с этим в лесных угодьях Октябрь- ского и Облученского районов происходили синхронные (по годам) колебания численности (коэффициент корреляции (r) составил 0,94 при 99 % значимости). Вероятно, эта синхронность поведения объясняется одинаковыми кормовыми условиями, повлекшими соответствующие изменения. Колебания числа белки в Биробиджанском районе несколько хуже соотносятся с изменениями численностей в Облученском (r=0,7 при 95 % значимости) и Октябрьском (r=0,72 при 95 % значимости) районах.
Некоторое несоответствие в тенденциях динамики обусловлено различиями в интенсивности плодоношения кедра в пределах районов. Действительно, в какие-то годы его урожай охватывает почти все три района области, в другие – лишь отдельные их участки, поэтому и происходят локальные, несколько дифференцированные размахи численности местных субпопуляций белки [7]. Однако в целом высокая плотность или, наоборот, депрессивное состояние популяции охватывают почти всю площадь кедрово-широколиственных лесов области. Так, например, в 2004 г. наблюдалось резкое снижение количества белки в районах области, а в 2007 г. – всплеск численности, уровень которой превзошел равновесное состояние почти в 2,5 раза (рис. 1, 3). Неурожай и, наоборот, обильное плодоношение орехов кедра корейского в 2003 и 2006 гг. соответственно также отмечались в некоторых районах Хабаровского края [4, 13].
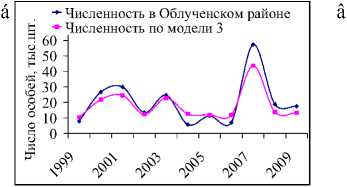
Как видно на рис. 4а–г, модели (2) и (3) вполне непло-
Численность в Октябрьском районе

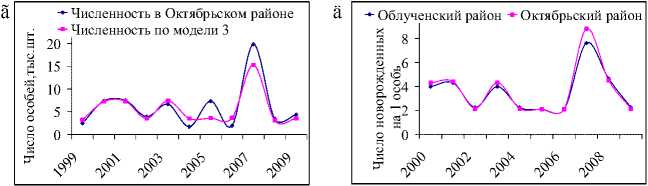
Рис. 4. Фактическая и модельная численности белок на территории Облученского по моделям 2 (а) и 3 (б) и Октябрьского районов по моделям 2 (в) и 3 (г). Динамика коэффициента рождаемости для белок, обитающих в Облученском и Октябрьском районах (д)
хо улавливают общие тенденции динамики численности белок в Облученском и Октябрьском районах от урожая кедровых орехов и описывают основные всплески наблюдаемой численности.
Согласно оценкам коэффициентов рождаемости, полученным на основе модели (3), среднегодовая плодовитость одной половозрелой самки в этих двух районах соотносится с оценками для всей области и составляет после средних урожаев кедра 8–9 особей, после слабо урожайных и неурожайных сезонов – около 4 (рис. 4д). После сезонов с обильным плодоношением кедра среднегодовая величина рождаемости в районах несколько отличается; в Облученском районе одна самка рожает в среднем 15 особей, Октябрьском – 17–18. Заметим, что максимальный коэффициент рождаемости (7,621), полученный по модели (3) в Облученском районе, близок по своему значению к величине скорости годового воспроизводства в отсутствии лимитирования (7,623). По всей вероятности, здесь отражается тот факт, что в весьма благоприятных кормовых условиях численность белки практически не лимитируется запасами пищи. В Октябрьском районе наибольший коэффициент рождаемости несколько ниже соответствующего коэффициента репродуктивного потенциала, более того, максимально возможная численность белки меньше максимальной наблюдаемой в исследуемый период, поэтому наибольший прирост популяции здесь вряд ли достижим при современных условиях беличьих местообитаний.
Тенденция увеличения количества белки в Облучен-ском районе определяется ростом и стабилизацией ее численности в лесных угодьях ООО «Сутара» и
ООО «Ирбис» (рис. 5а), ООиР (рис. 5б) Тенденция роста популяции в Октябрьском районе, возможно, обусловлена стабилизацией и даже некоторым увеличением количества белки в лесных угодьях ООО «Сутара» (здесь расположены 33 % угодий), ОХ ОРО «Диана» и заказнике «Журавлиный». Достаточно устойчивый подъем численности наблюдается в Биробиджанском районе, который отчасти вызван увеличением числа белки в заказнике «Шухи-Поктой» (рис. 5г).
Вероятно, на рост численности белки в Облученском и Биробиджанском районах повлияло также увеличение их количества в заповеднике «Бастак», лесные земли которого на 9 % покрыты лесными насаждениями с преобладанием кедра, на 8 % – ели, на 13 % – пихты и на 14 % дуба [15]. В случае большого прироста популяции не исключена миграция зверьков на сопредельные территории, поскольку объем кормовых ресурсов заповедника ограничен (максимально возможная модельная численность ниже максимальной учетной численности).
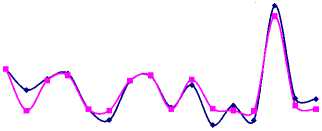
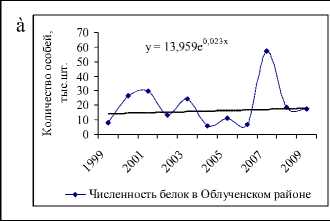
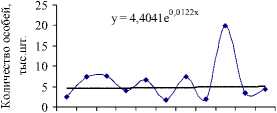
Значительный рост числа белок был отмечен на территории охотничьего хозяйства «Ирбис» (рис. 5а), площадь которого на 94 % покрыта лесными угодьями, из них 5 % составляют кедровые леса, 9 % – кедрово-широколиственные, 15 % – елово-пихтовые и 18 % – дубовые леса, составляющие в целом неплохую кормовую базу для популяции. Более того, согласно оценкам модели (1), экологическая ниша белки находится здесь в удовлетворительном состоянии. Колебания численности белок с 2004 по 2009 гг. неплохо коррелируют с изменениями интенсивности плодоношения кедра (рис. 6а), а модели (2) и (3) хорошо описывают оценки их численности (рис. 6б, 6в).
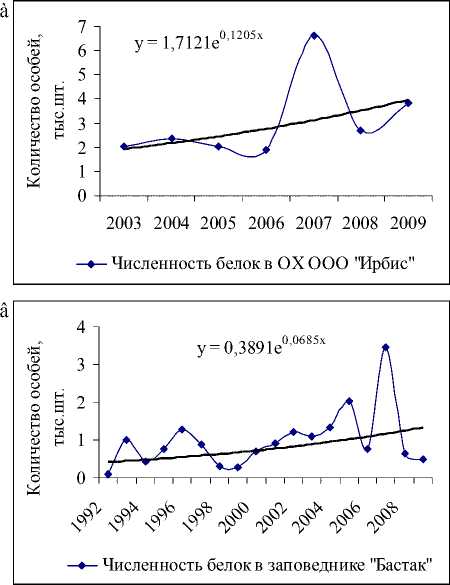
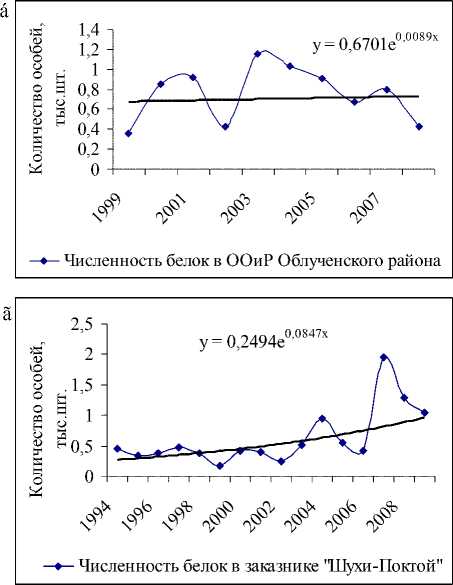
Рис. 5. Данные учета численности белки на территории ООО «Ирбис» и соответствующие им экспоненциальные тренды: а) с 2003 по 2009 гг.; б) ООиР Облученского района с 1999 по 2008 гг.; в) заповедника «Бастак» с 1992 по 2009 гг.; г) заказника Шухи-Поктой с 1994 по 2009 гг.
à
-■- Запасы орехов кедра корейского
-♦- Численность белок в "Ирбис"
á
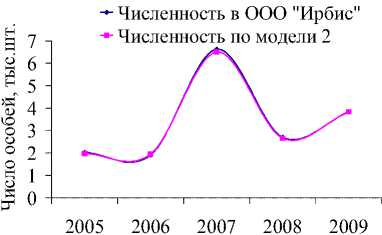
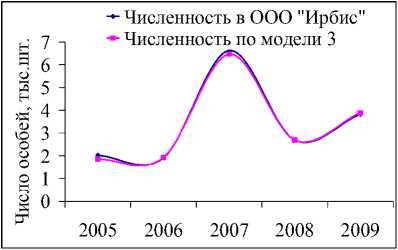
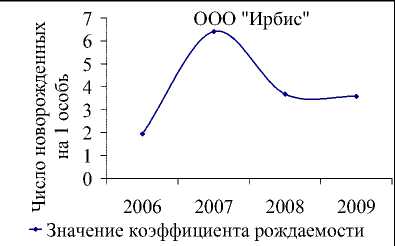
Рис. 6. Динамика численности белки в ООО «Ирбис» в зависимости от запасов орехов кедра корейского (ряд данных урожайности смещен на 1 год вперед) (а). Фактическая и модельная численность, оцененная по модели 2 (б) и 3 (в). Динамика коэффициента рождаемости для белок, обитающих в «Ирбисе» (д)
Значение репродуктивного потенциала, вычисленного по модели (2), несколько меньше, чем оценки для всей области и ее районов, и составляют 7 новорожденных особей на одну взрослую самку. Это значение соотносится со среднегодовой плодовитостью белки, определенной по модели (3) при средней интенсивности урожая (рис. 6д). Видимо, высокая оценка репродуктивных возможностей белки, составляющая 13 особей в 2007 г., включает в себя рост численности в связи с миграцией зверьков с сопредельных территорий.
На другой территории лесных угодий области, где произрастают кедрово-широколиственные леса, численность белок невелика. В заказнике «Журавлином», некогда достаточно большая популяция (3 тыс. особей) заметно сократилась и в последние годы составляет в среднем – 350 белок. Тем не менее, в последнее десятилетие наметилась некоторая тенденция роста численности, и в отдельные годы их количество достигало 700–950 особей. В пределах границ охотничьего хозяйства «Диана» насчитывается в среднем 750 особей.
Выводы
Таким образом, в последнее десятилетие наблюдается стабилизация численности белки в ЕАО, вызванная ростом ее количества почти на всех учетных территориях. Наиболее заметная тенденция роста числа белок отмечается в Биробиджанском районе. Динамика численности популяции в ЕАО в значительной мере определяется ее изменением в лесных угодьях Облученского района.
Согласно модельным расчетам максимально возможная модельная численность белок сопоставима с максимальной учетной численностью, наблюдаемой за рассматриваемый период. По-видимому, экологическая ниша этого вида на большинстве территорий его обита- ния находится в удовлетворительном состоянии: потребности популяции вполне удовлетворены ресурсами (запасом корма, размером ареала, наличием мест для размножения и подрастания потомства и т.п.). Наименее заполнена экологическая ниша белки в лесных угодьях Облученского и Биробиджанского районов, наиболее – Октябрьского.
Показано, что ежегодные колебания численности белки определяются урожайностью орехов кедра корейского в годы, предшествующие размножению. Неурожай основных видов кормов неблагоприятно отражается на сохранении популяции, ее численность резко падает, животные мигрируют в другие биотопы в поисках лучших кормовых условий. Согласно модельным оценкам, среднегодовая плодовитость самки белки на территории области составляет после сезонов обильного плодоношения кедра 20 особей, после средних урожаев – 8–10 и в малоурожайные сезоны – около 4–5.
Исследования проведены при частичной финансовой поддержке РФФИ (проект № 09-04-00146-а) и ДВО РАН (проект № 09-I-Р15-01, 09-I-ОБН-12, 09-II-СО-06-006, 09-III-А-09-498, 09-I-П23-13).
Список литературы Анализ влияния запасов корма на динамику численности популяции белки (на примере Еврейской автономной области)
- Ашихмина Е.В., Скалецкая Е.И., Фрисман Е.Я., Куликов А.М. Математическая модель динамики величины заготовок на примере локальной популяции маньчжурской белки//Журнал общей биологии. 1982. Т 43, № 5. С. 246-257.
- Доклад о состоянии и об охране окружающей природной среды Еврейской автономной области в 2000 г. Биробиджан: Комитет природных ресурсов по Еврейской автономной области. 2001. 100 с.
- Доклад о состоянии и об охране окружающей природной среды Еврейской автономной области в 2002 г. Биробиджан: Управление природных ресурсов и охраны окружающей среды МПР РФ по Еврейской автономной области. 2003.1 65 с.
- Дунишенко Ю.М. Мероприятия по восстановлению и увеличению численности диких копытных животных. Хабаровск: Хабаровский фонд диких животных, 2004. 40 с.
- Колосов А.М. Зоогеография Дальнего Востока. М.: Мысль, 1980. 254 с.
- Кормовая база в охотничьих хозяйствах/Е.А. Даниленко, В.А. Кузьмин, И.Ф. Кузьмин и др. М.: Лесная промышленность, 1979. 96 с.
- Костенко В.А. Грызуны (Rodentia) Дальнего Востока России. Владивосток: Дальнаука, 2000. 210 с.
- Куренцова Г.Э. Очерк растительности Еврейской автономной области. Владивосток: Дальневосточ. кн. изд-во, 1967. 63 с.
- Лесной план Еврейской автономной области на 2009-2018 годы. Разработчик -исполнитель: Филиал ФГУ «ВНИИЛМ» «ДальНИИЛХ». Биробиджан, 2008. Кн. 1. 23 с.
- Медико-экологический атлас Хабаровского края и Еврейской автономной области/автор-сост. атласа В.И. Волков. Хабаровск, 2005. 112 с.
- Михайловский Б.А., Залесов А.С., Рычкова Н.Н. Промысловая оценка и освоение биологических ресурсов: сб. науч. тр. Киров, 1988. С. 43-59.
- Наумов С.П. Зоология позвоночных: учебник для студентов пед. вузов по биол. спец. 4-е изд., перераб. М.: Просвещение, 1982. 464 с.
- Отчет «Состояние ресурсов охотничьих животных, их численность и добыча по регионам России в сезон 2006-2007 гг.». Киров: ГНУ ВНИИОЗ им. проф. Б.М. Житкова, 2007. 77 с. . URL: http://www.vniioz.kirov.ru/inst/structure/ohotres/otchet_2008.pdf
- Природные ресурсы Еврейской автономной области//В.И. Журнист, Р.М. Коган, Т.Е. Кодякова, Т.М. Комарова, Т.А. Рубцова и др. Биробиджан: ИКАРП ДВО РАН, 2004. 112 с.
- Проект организации и ведения лесного хозяйства Государственного природного заповедника «Бастак». Хабаровск: Министерство природных ресурсов РФ, ФГУП «Дальневосточное государственное лесоустроительное предприятие». 2002. Т. I. С. 22-40.
- Рубцова Т.А. Флора Малого Хингана. Владивосток: Дальнаука, 2002. 194 с.
- Русанов Я.С. Охота и охрана фауны. М.: Лесная промышленность, 1973. 144 с.
- Фрисман Е.Я., Ревуцкая О.Л., Неверова Г.П. Анализ популяционной динамики промысловых млекопитающих Среднего Приамурья России: математическое моделирование и оценка ресурсного потенциала//Биологические ресурсы Дальнего Востока: комплексный региональный проект ДВО РАН. М.: Товарищество научных изданий КМК, 2007.С. 184-202.
- Фрисман Е.Я. Математические модели динамики численности локальной однородной популяции. Владивосток: Дальрыбвтуз, 1996. 58 с.
- Фрисман Е.Я. Математическое моделирование и анализ механизмов популяционной динамики промысловых видов животных//Региональные проблемы. 2005. № 6-7. С. 149-153.
