Анализ взаимосвязи климатических факторов и генетического разнообразия популяций дуба черешчатого в разных частях Республики Башкортостан

Автор: Янбаев Руслан Юлаевич, Бахтина Светлана Юрьевна, Садыков Айдар Харисович, Янбаев Юлай Аглямович
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 4, 2022 года.
Бесплатный доступ
Республика Башкортостан, площадью около 143 тыс. км2, расположена на Южном Урале, где геоморфологическая неоднородность территории, воздействие воздушных масс Атлантического океана, внутренних районов Арктики и Сибири формируют существенную внутри- и межгодовую изменчивость температуры и осадков. Цель работы - анализ динамики во времени и пространстве этих двух важнейших экологических факторов на фоне результатов, полученных при исследовании генетических различий популяций дуба черешчатого. Для генетического анализа растительный материал отобран в 14 естественных насаждениях региона, использованы 412 локусов однонуклеотидных полиморфизмов ядерной ДНК. Также использована информация по динамике температур и количеству осадков в разрезе месяцев на южной, центральной, северной и западной частях республики в 2004-2019 г. Показано, что температуры во всех метеорологических станциях варьируются по месяцам и годам достаточно синхронно. По режиму и количеству осадков исследованные территории различались намного больше; различия доходили до статистически достоверного уровня в 59.7% (преимущественно в месяцы вегетационных сезонов) сравнениях пар метеостанций по отдельным месяцам. При этом популяции дуба черешчатого обладают близкими уровнями генетического разнообразия и генетически относительно слабо дифференцированы, что доказывает устойчивость системы к воздействию факторов окружающей среды в условиях глобального изменения климата. Выявленный феномен может быть связан с эффективностью генетически реализованного потока пыльцы, в том числе между удаленными насаждениями.
Климат, южный урал, дуб черешчатый, генетическое разнообразие
Короткий адрес: https://sciup.org/147239682
IDR: 147239682 | УДК: 551.583.4 | DOI: 10.17072/1994-9952-2022-4-327-334
Analysis of the relationship between climatic factors and genetic diversity of pedunculate oak populations in different parts of the Republic of Bashkortostan
The geomorphological heterogeneity of the Southern Urals, the impact of air masses from the Atlantic Ocean and internal territories of the Arctic and Siberia form a significant intra- and interannual variation of temperature and precipitation in the Republic of Bashkortostan, which is a Russian region with an area of about 143 thousand square kilometres. The aim of the present study is to analyse the spatial and temporal dynamics of these two most important environmental factors in the light of the results obtained in the study of genetic differences in pedunculate oak populations. For genetic analysis, plant material was sampled in 14 natural stands of the region, 412 loci of single nucleotide polymorphisms of nuclear DNA were used. Information on the dynamics in 2004-2019 of temperatures and precipitation in the southern, central, northern and western parts of the Republic of Bashkortostan is also used in the context of months. We have shown that temperatures in all meteorological stations vary by months and years quite synchronously. The studied territories differed much more in the mode and amount of precipitation, the differences reached a statistically significant level of 59.7% (mainly in the months of the growing seasons) of monthly comparisons of pairs of the stations. At the same time, the populations of pedunculate oak have similar levels of genetic diversity and are genetically relatively weakly differentiated, which proves the system's stability to environmental factors under global climate change. It is discussed that the phenomenon revealed may be related to the effectiveness of a genetically realized pollen flow, including between remote stands.
Текст научной статьи Анализ взаимосвязи климатических факторов и генетического разнообразия популяций дуба черешчатого в разных частях Республики Башкортостан
Одним из последствий глобального изменения климата является повышение температуры на Земле. По информации Всемирной метеорологической организации, 2015–2019 гг. и 2010–2019 гг. стали самыми теплыми за всё время проведения метеонаблюдений [Доклад…, 2020]. В целом, начиная с 80-х гг. прошлого столетия, каждое новое десятилетие было более теплым. Следовательно, воздействие на живые организмы таких экологических факторов, как температура окружающей среды и тесно связанные с ней количество и режим осадков изменяются, выходя, в том числе, за пределы оптимальных значений [Исаев, 2001]. Растения, особенно виды лесных древесных растений, мало приспособлены для быстрой миграции в более благоприятные условия среды. По этой причине они вынуждены адаптироваться к меняющимся условиям преимущественно по месту своего произрастания. При этом потенциал для адаптации обеспечивается существующим в популяциях генетическим разнообразием [Degen et al., 2021]. Следовательно, его выявление, вместе с анализом динамики климата во времени, позволяют прогнозировать возможные адаптационные процессы популяционного уровня. Исследования роли экологических факторов в формировании популяционных генофондов растений стали особо популярными и результативными из-за появления генетических маркеров нового поколения [Peterson et al., 2012; Leroy et al., 2019] с использованием в качестве генетических маркеров локусов однонуклеотидных полиморфизмов [Degen et al., 2021].
Республика Башкортостан – расположенный на Южном Урале российский регион с площадью около 143 тыс. км2, где геоморфологическая неоднородность территории, воздействие воздушных масс Атлантического океана, внутренних районов Арктики и Сибири формируют существенную и отличающуюся от других регионов внутри- и межгодовую изменчивость температуры и осадков. Это обстоятельство влияет на формирование генофонда популяций дикорастущих древесных растений Урала и их генетическое разнообразие [Пришнивская, Красильников, Боронникова, 2016]. Цель работы – анализ динамики во времени и пространстве на Южном Урале (в Башкортостане) этих двух важнейших экологических факторов в 2004–2019 гг на фоне результатов, полученных при исследовании в южной части уральских гор генетических различий популяций дуба черешчатого ( Quercus robur L.). Насаждения данного экономически и экологически важного вида сильно пострадали во время экстремально холодных зим XX в., сопровождаемых увеличением количества энтомовредителей и болезней [Царалунга, Фурменкова, Крюкова, 2015], особенно на восточной границе ареала в Башкортостане [Попов, 1980]. Однако потепление климата может дать здесь дубу черешчатому с его исключительной засухоустойчивостью возможность восстановления насаждений и смещения границы ареала на север [Albert et al., 2018].
Материалы и методы исследования
Образцы для лабораторного анализа отобраны на 14 пробных площадях дуба черешчатого. Насаждения для их закладки (рис. 1, табл. 1) выбраны, исходя из данных о распространении дубрав на территории Башкортостана [Попов, 1980]. Низкогорья западного макросклона Южного Урала (здесь представлены основные площади и запасы дуба черешчатого, вид в древостоях может доминировать, особенно на юге Башкортостана) представлены выборками ЗИЛ, АРХ-1, АРХ-2, АРХ-3 и ИГЛ. На Уфимском плато (вид входит в состав темнохвойно-широколиственных лесов) заложены пробные площади МШК и КРД.
Остальные участки представляют колочные и островные степные и лесостепные насаждения Башкирского Предуралья (ТМЗ, ИЛШ, БУР, БРС, БЛГ и БВШ) и Мясогутовской лесостепи (СЛВ). Одновременно изученные дубравы представляют южную, центральную, северную и западную части Республики Башкортостан, соответственно и разделены расстояниями в 432 (с. Зилаир – с. Аскино) и 292 км (с. Малояз – г. Туймазы) в широтном и долготном направлениях. В большинстве пробных площадей отобраны по 10 деревьев репродуктивного возраста (в Зилаирском лесничестве с 40) не менее чем в 50 м друг от друга для уменьшения эффекта семейной кластеризации. С них отобраны образцы камбия для выделения ДНК [Dumolin, Demesure, Petit, 1995].
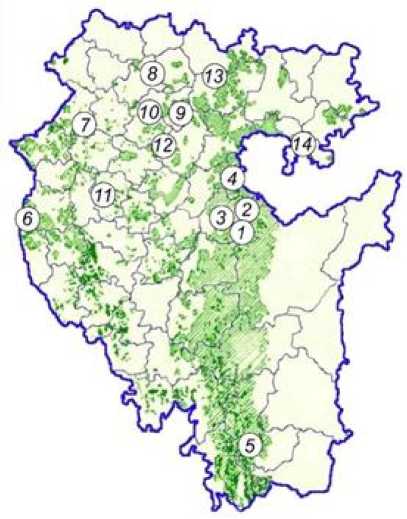
Рис. 1. Расположение исследованных выборок дуба черешчатого на схематической карте дубрав Республики Башкортостан [по Попову, 1980].
Сведения о выборках приведены в табл. 1; насаждения с доминированием и участием дуба черешчатого в составе древостоев закрашены и показаны штриховкой, соответственно
[The location of the pedunculate oak samples studied on the species schematic map in the Republic of Bashkortostan [according to Popov, 1980].
Note: information about the samples is given in Table 1; stands with the dominance and participation of the pedunculate oak in the stands are painted over and shown with hatching, respectively]
Таблица 1
Географические координаты и генетическое разнообразие выборок дуба черешчатого
[Geographical coordinates and genetic diversity of the pedunculate oak samples]
|
Районы |
Выборки |
Долгота, широта |
Показатели генетического разнообразия |
|||
|
Ae |
Ho |
He |
Fis |
|||
|
Архангельский |
1. АРХ-1 |
57.255, 54.517 |
1.339 |
0.204 |
0.199 |
–0.038 |
|
2. АРХ-2 |
57.116, 54.449 |
1.349 |
0.215 |
0.204 |
–0.057 |
|
|
3. АРХ-3 |
56.798, 54.420 |
1.326 |
0.209 |
0.191 |
–0.077 |
|
|
Иглинский |
4. ИГЛ |
57.029, 54.838 |
1.342 |
0.222 |
0.201 |
–0.093 |
|
Зилаирский |
5. ЗИЛ |
57.320, 52.230 |
1.351 |
0.208 |
0.206 |
–0.015 |
|
Туймазинский |
6. ТМЗ |
53.512, 54.441 |
1.344 |
0.215 |
0.204 |
–0.048 |
|
Илешевский |
7. ИЛШ |
54.478, 55.362 |
1.334 |
0.205 |
0.197 |
–0.039 |
|
Бураевский |
8. БУР |
55.647, 55.862 |
1.335 |
0.214 |
0.196 |
–0.085 |
|
Мишкинский |
9. МШК |
58.318, 55.490 |
1.349 |
0.217 |
0.206 |
–0.065 |
|
Бирский |
10. БРС |
57.029, 54.838 |
1.345 |
0.216 |
0.205 |
–0.051 |
|
Благоварский |
11. БЛГ |
57.029, 54.838 |
1.331 |
0.212 |
0.196 |
–0.073 |
|
Благовещенский |
12. БВШ |
58.317, 55.489 |
1.341 |
0.202 |
0.201 |
–0.016 |
|
Караидельский |
13. КРД |
58.319, 55.491 |
1.337 |
0.204 |
0.197 |
–0.038 |
|
Салаватский |
14. СЛВ |
58.237, 55.172 |
1.330 |
0.210 |
0.197 |
–0.062 |
Примечание. Ае – эффективное число аллелей, Но и Не –наблюдаемая и ожидаемая етерозиготность, Fis – индекс фиксации Райта (коэффициент инбридинга особи относительно популяции).
Для анализа динамики климатических показателей в регионе исследования использована информация, собранная Зилаирским, Архангельским, Аскинским и Туймазинским метеостанциями Башкирского управления по гидрометеорологии и мониторингу окружающей среды. Для метеорологических данных (месячные значения температур и количества осадков) вычислены средние значения, медианы, пределы изменения показателей и коэффициент вариации. Для анализа выбран период 2004–2019 гг, который входит в период наибольшего глобального потепления [Доклад…, 2020]. Статистическая достоверность различий вариационных рядов, с учетом их непараметрического распределения, определена с применением коэффициента корреляции Спирмена. Все вычисления выполнены в программе STATISTICA 13.3. Детальное описание методов лабораторных анализов было приведено ранее [Degen et al., 2021]. Генетические сравнения популяций проведены с применением в качестве генетических маркеров однонукле-тидных полиморфизмов (SNP, Single Nucleotide Polymorphisms) из недавно разработанного нами [Degent et al., 2021] набора 412 локусов ядерной ДНК. Для каждой выборки вычислены эффективное число аллелей (Ае), наблюдаемая и ожидаемая гетерозиготность (Hо и Hе, соответственно) и коэффициент инбридинга особи относительно популяции (Fis). Уровень генетических различий между популяциями оценивался с использованием генетического расстояния Грегориуса d0 [Gregorius, 1984].Cоответствующие вычисления проведены в программе GDA_NT [Degen, 2022]. Определение числа генетических групп исследованных выборках проведено в программе STRUCTURE [Pritchard et al., 2000], а графическое представление результатов – в программе CLUMPAK [Kopelman et al., 2015].
Результаты и их обсуждение
Среднемесячная температура 2004–2019 гг. изменяется по данным четырех метеостанций от 3.0±0.9 до 4.7±0.9; она выше в Архангельском (4.6±0.9ºС) и Туймазинском р-нах (4.7±0.9ºС) с более благоприятными для дуба черешчатого [Попов, 1980] лесорастительными условиями. Как в северной части исследованной территории (метеостанция в с. Аскино), так и на границе со степной зоной (с. Зилаир) показатель ниже – 3.0±0.9, и 3.2±0.9ºС, соответственно. Тем не менее, эти различия не столь выражены по сравнению с данными по среднемесячному количеству осадков. Экстремально низких температур ниже –40ºС, обусловивших массовое усыхание дубрав во второй половине прошлого века [Попов, 1980], не наблюдалось. Анализ динамики температур показал, что они в данных метеостанций изменяются по месяцам в целом синхронно. Из всех теоретически возможных 72 случаев попарный коэффициент корреляции Спирмена был статистически достоверным на разных уровнях значимости. Другая картина наблюдается при сравнении вариационных рядов с данными по месячному количеству осадков за 2004–2019 гг. В среднем за месяц рассмотренных 16 лет выпадало осадков 51.1±2.2 (с. Архангельское), 48.9±2.2 (с. Аски-но), 41.0±1.9 (с. Зилаир) и 39.8±1.9 мм (г. Туймазы). Однако средние величины не дают представления о размахе изменчивости признака – он варьируется в пределах отдельных месяцев в широких пределах – в пределах 0.5–170.8 мм, 2.7–156.7 мм, 1.2–110.8 мм и 0.8–170.4 мм, соответственно. Более того, часто месяцы с обильными осадками или малыми их количествами в разных районах не совпадают. В таблице 2 приведена матрица, показывающая 43 из 72 изученных месяцев (59.7 %), по которым динамика количества осадков в парах метеостанций не является статистически достоверной – то есть не синхронной.
Таблица 2
Месяцы со статистически достоверными различиями количества осадков пар метеостанций Башкортостана
[Months with statistically significant differences in precipitation of pairs of weather stations of Bashkortostan]
|
Метеостанции |
Аскино |
Зилаир |
Туймазы |
|
Архангельское |
Март, сентябрь, октябрь, ноябрь, декабрь |
Апрель, июнь, июль, август, сентябрь, октябрь, ноябрь, декабрь |
Март, июль, сентябрь, октябрь, ноябрь, декабрь |
|
Аскино |
- |
Март, июнь, июль, август, сентябрь, октябрь, ноябрь, декабрь |
Март, апрель, июль, август, сентябрь, октябрь, ноябрь, декабрь |
|
Зилаир |
- |
- |
Апрель, июнь, июль, август, сентябрь, октябрь, ноябрь, декабрь |
Наиболее отличается от других по этому показателю южная, Зилаирская метеостанция, расположенная на границе со степной зоной. Количество осадков в разных метеостанциях статистически значимо коррелировало лишь в январе и феврале. И, наоборот, с апреля по октябрь (в вегетационный период) дифференциация изученных частей Башкортостана по ним становились намного более выраженной. Таким образом, количество и время выпадения осадков может быть для дуба черешчатого сравнительно более критическим явлением, воздействующим на состояние дубрав через действие отбора, чем месячные температуры. В то же время, влияние температуры, другого важного экологического фактора, на популяции может оказываться опосредованно через его воздействие на режим и количество осадков [Исаев, 2001]. В среднем для 14 изученных популяций эффективное число аллелей составило значение Ае = 1.339±0.002, наблюдаемая и ожидаемая гетерозиготность, Но = 0.211±0.002 и Не = 0.200±0.001, коэффициент инбридинга Fis = –0.054 ± 0.06. Данные табл. 1 и вычисление коэффициентов вариации (0.6, 2.8 и 2.3% для Ае, Но и Не, соответственно) показывают существенное сходство уровней генетического разнообразия как в соседствующих насаждениях, так и в удаленных друг от друга дубравах. Генетическое расстояние между выборками в среднем по 412 локусам SNP составило значение d0 = 0.069±0.001 с изменениями между парами от d0 = 0.054 до d0 = 0.084. При этом генетические различия популяций в пределах центральной, западной и северной частей Башкортостана выражены на уровне, сопоставимым с генетической дифференциацией более удаленных друг от друга выборок, иногда превышая ее. Во всех выборках отмечен небольшой эксцесс наблюдаемой гетерозиготности. Результаты работы в программах STRUCTURE и CLUMPAK подтвердили слабую генетическую подразделенность дубрав Башкортостана. На рисунке 2 для наиболее информативных К = 2 и К = 3 показано, что исследованные выборки не формируют разные генетические группы и не образуют отличающиеся кластеры. Таким образом, близкая динамика месячных температур в исследованных частях Башкортостана в 2004–2019 гг. совпадает с результатами генетических исследований дуба черешчатого, демонстрируя относительно слабую дифференциацию и близкие уровни генетического разнообразия популяций. И наоборот, сходство популяционных генофондов на этой территории наблюдается на фоне выраженных и статистически значимых различий южной, центральной, западной и северных частях региона по режиму и количеству осадков.
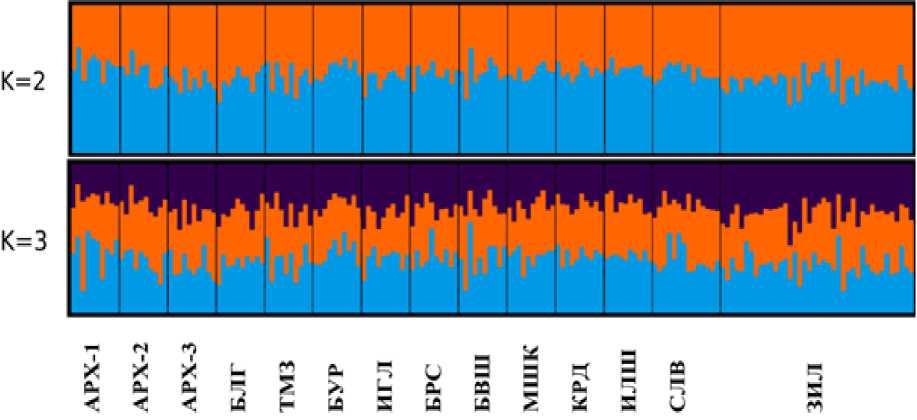
Рис. 2. Популяционная структура дуба черешчатого по результатам анализа в программе STRUCTURE. Вертикальные полосы показывают особей выборок, представленных разными цветами пропорционально принадлежности к тому или иному кластеру
[The population structure of the pedunculate oak according to the results of the analysis in the STRUCTURE program. Note: vertical bars represent by different colors individuals of samples in proportion according to belonging to a particular cluster]]
Этот феномен требует дополнительного исследования и, по-видимому, имеет под собой комплекс причин, в том числе генетических, связанных с эффективностью генетически реализуемого потока пыльцы. Нами, по данным находящегося в открытом доступе (сайт Министерства лесного хозяйства Башкортостана, URL: forest.bashkortostan.ru) Лесного плана 2016 г., был проведен анализ соотношения площадей насаждений 24 лесничеств, где представлены широколиственные древесные растения региона. Установлено (табл. 3), что во всех хозяйствах доминирует Tilia cordata , а доля других видов существенно варьируется. Доля насаждений с доминированием Q. robur сравнительно мала во всех лесничествах – в 45.8% из них занимают 0.6–3.0% лесопокрытой площади. Тем не менее, он во всех случаях является непременным спутником других представителей широколиственных лесов. Возможно, эти деревья представляют «генные тропы», которые дают возможность потоку генов выравнивать частоты аллелей и сближать генофонды и уровни генетического разнообразия популяций всего Южного Урала. Вряд ли может этот процесс быть реализован путем распространения тяжелых желудей дуба черешчатого.
Таблица 3
Доли насаждений широколиственных древесных растений в лесничествах Республики Башкортостан (в %)
[The proportion of broad-leaved stands in forestry enterprises of the Republic of Bashkortostan (in %)]
|
Виды |
Средняя |
Пределы изменений |
Медиана |
Коэффициент вариации, % |
|
Quercus robur |
4.2±0.6 |
0.6-10.4 |
3.7 |
68.1 |
|
Tilia cordata |
86.7±2.2 |
61.4-98.4 |
89.3 |
12.3 |
|
Acer platanoides |
6.9+1.7 |
0.1-30.4 |
2.7 |
128.6 |
|
Ulmus glabra , U. laevis |
2.6+0.5 |
0.1-7.9 |
1.8 |
92.8 |
Нами ранее было показано [Bushbom, Yanbaev, Degen, 2011), что успешный генетический поток может обеспечиваться за счет эффективности потока «отцовских» аллелей на дальние расстояния при помощи пыльцы. Реконструкция генотипов зародышей при помощи микросателлитных генетических маркеров в удаленном на много десятков километров от других дубрав малой популяции выявила, что по крайней мере 35% пыльцы попали в насаждение из дальних источников. В настоящее время насаждения широколиственных лесов Башкортостана существенно фрагментированы в пространстве, в первую очередь, при сельскохозяйственном освоении территории. Тем не менее, уменьшение лесистости почти в два раза произошло лишь за несколько последних веков [Попов, 1980]. Это, вероятно, еще не привело у долгоживущего дуба черешчатого к заметному обеднению генофондов популяций, оказавшихся в пространственной изоляции безлесными территориями, и к увеличению генетической дифференциации между ними. Эти сценарии могут лежать в основе показанного нами феномена близости генофондов удаленных популяций региона с относительно большой изменчивостью климата – выраженной динамикой в пространстве и во времени количества осадков.
Выводы
-
1. Температура в южной, центральной, северной и западной частях Республики Башкортостан варьируется по месяцам и годам в целом синхронно, что доказывается положительной статистически достоверной корреляцией ее значений.
-
2. Количество осадков по месяцам и годам является более изменчивым показателем, асинхронность (статистически достоверная на разных уровнях значимости в 59.7% сравнений пар метеостанций) выражена преимущественно в месяцы вегетационного периода. Таким образом, данный показатель потенциально является более критическим экологическим фактором для популяций дуба черешчатого.
-
3. Применение генетических маркеров 412 локусов однонуклеотидных полиморфизмов ядерной ДНК позволило выявить сравнительно низкую генетическую дифференциацию популяций дуба черешчатого региона и близкие значения показателей их генетического разнообразия, несмотря на относительно большую динамику в пространстве и во времени количества осадков. Этот феномен демонстрирует устойчивость популяционной системы к воздействию этого климатического фактора.
Список литературы Анализ взаимосвязи климатических факторов и генетического разнообразия популяций дуба черешчатого в разных частях Республики Башкортостан
- Доклад о научно-методических основах для разработки стратегий адаптации к изменениям климата в Российской Федерации (в области компетенции Росгидромета). СПб.; Саратов: Амирит, 2020. 120 с. URL: http://cc.voeikovmgo.ru/images/ dokumenty/2G2G /dokladRGM.pdf.
- Исаев А.А. Экологическая климатология. M.: Науч. мир, 2001. 456 с.
- Попов Г.В. Леса Башкирии. Уфа, 1980. 144 с.
- Пришнивская Я.В., Красильников В.П., Боронникова С.В. Mолекулярно-генетическая идентификация популяций Pinus sylvestris L. на востоке Русской равнины на основании полиморфизма ISSR-маркеров // Вестник Пермского университета. Сер. Биология. 2016. Вып. 2. С. 171-176.
- Царалунга В.В., Фурменкова Е.С., Крюкова А.А. Внешние признаки патологии дуба черешчатого. Воронеж, 2Gi5. 231 с.
- Albert M. et al. Quantifying the effect of persistent dryer climates on forest productivity and implications for forest planning: a case study in northern Germany // Forest Ecosystems. 2Gi8. Vol. 5 (33). DOl: https://doi.org/iG.ii86/s4G663-Gi8-Gi52-G.
- Bushbom J., Yanbaev Yu., Degen B. Efficient long-distance gene flow into an isolated relict oak stand // Journal of Heredity. 2011. Vol. 102, № 4. P. 464-472.
- Degen B. GDA-NT 2G2i a computer program for population genetic data analysis and assignment // Conservation Genetics Resources. 2G22. DOl: iG.iGG7/si2686-G22-Gi283-2.
- Degen B. et al. Applying targeted genotyping by sequencing with a new set of nuclear and plastid SNP and indel loci for Quercus robur and Quercus petraea // Conservation Genetics Resources. 2021. Vol. 13 (4). P. 345-347.
- Dumolin S., Demesure B., Petit R.J. Inheritance of chloroplast and mitochondrial genomes in pedunculate oak investigated with an efficient PCR method // Theoretical and Applied Genetics. 1995. Vol. 91. P. 1253 -1256.
- Gregorius H.R. A unique genetic distance // Biometrical Journal. 1984. Vol. 26, № 1. P. 13-18.
- Kopelman N.M. et al. Clumpak: a program for identifying clustering modes and packaging population structure inferences across K // Molecular Ecology Resources. 2015. Vol. 15. P. 1179-1191.
- Leroy T. et al. Adaptive introgression as a driver of local adaptation to climate in European white oaks // New Phytologist. 2019. doi:10.1111/nph.16095.
- Peterson B.K. et al. Double Digest RADseq: An inexpensive method for de novo SNP discovery and genotyping in model and non-model species // Plos One. 2012. Vol. 7, № 5. https://doi.org. 10.1371/journal.pone.0037 135.
- Pritchard J.K., Stephens M., Donnelly P. Inference of population structure using multilocus genotype data // Genetics. 2000. Vol. 155. P. 945-959.
