Анализ закономерностей распределения аллелей авенин-кодирующих локусов у сортов овса посевного отечественной селекции

Автор: Любимова А.В., Еремин Д.И.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Агрономия
Статья в выпуске: 11, 2019 года.
Бесплатный доступ
Проламин-кодирующие локусы успешно применяются в качестве маркерных систем для выявления адаптивно-значимых ассоциаций генов. Проламины овса - авенины - наследуются группами и контролируются тремя независимыми локусами: Avn A, Avn B, Avn C. Цель исследования - изучение связи между географическим распределением аллелей авенин-кодирующих локусов у образцов овса и природно-климатическими факторами для выявления аллелей-маркеров адаптивно-значимых ассоциаций генов. Материалом для исследования послужили индивидуальные зерновки 39 сортов и линий овса посевного российской селекции различного эколого-географического происхождения. Установлено, что 14 образцов были гомогенными по компонентному составу авенина. Остальные образцы содержали от 2 до 9 биотипов. Группы образцов происхождением из разных регионов отличались по частоте встречаемости аллелей АКЛ. Среди аллелей локуса Avn A самым распространенным был А2, по локусу Avn В наибольшей частотой встречаемости характеризовались аллели В1 и В4...
Овес, электрофорез, авенин-кодирующие локусы, маркерная селекция
Короткий адрес: https://sciup.org/140245600
IDR: 140245600 | УДК: 633.13:581.198(571.12.) | DOI: 10.36718/1819-4036-2019-11-30-38
The analysis of regularities of distribution of alleles of avenin-coding loci in domestic selection oat varieties
Prolamin-coding loci have been successfully used as marker systems to identify adaptively significant gene associations. Oat prolamins avenins are inherited by groups and controlled by three independent loci: Avn A, Avn B, Avn C. The aim of the research was to study the relationship between geographical distribution of alleles of avenin-coding loci in oat samples and natural and climatic factors to identify alleles-markers of adaptively significant gene associations. The materials for the study were individual grains of 39 varieties and lines of oats of Russian selection of different ecological and geographical origin. It was found out that 14 samples had been homogeneous in the component composition of avenin. The remaining samples contained from 2 to 9 biotypes. The groups of samples originating from different regions differed in the frequency of occurrence of ACL alleles. Among the alleles of Avn A locus, A2 was the most common, and B1 and B4 alleles were characterized by the highest frequency of occurrence at Avn B locus...
Текст научной статьи Анализ закономерностей распределения аллелей авенин-кодирующих локусов у сортов овса посевного отечественной селекции
Введение . Овес посевной ( Avena sativa L.) – ценная сельскохозяйственная культура, возделываемая на кормовые и зернофуражные цели, а также для питания человека.
Важнейшее направление в селекции овса – увеличение урожайности и повышение устойчивости сортов к биотическим и абиотическим факторам среды, а также улучшение их качественных характеристик. Большое значение при создании новых сортов имеет правильный под- бор исходного материала. В настоящее время в селекции все шире применяются не только традиционные способы, но и достижения генетики и биотехнологии. На большом количестве примеров показана пригодность использования разнообразных маркерных систем для выявления адаптивно-значимых ассоциаций генов. Такой анализ основан на оценке динамики частот аллелей маркерных локусов [1]. Очень эффективны для решения этих задач биохимические маркеры, в том числе спирторастворимые белки семян – проламины [2–5]. Установлена зависимость частоты встречаемости аллелей глиадин-кодирующих локусов пшеницы и гордеин-кодирующих локусов ячменя от климатических и почвенных условий [6–8]. Проламины овса называются авенинами. Компоненты авенина наследуются группами и контролируются тремя независимыми локусами: Avn A, Avn B, Avn C [9].
Известно, что сочетание аллелей авенин-кодирующих локусов (АКЛ) в генотипах носит неслучайный характер. В.А. Портянко установлена географическая зональность во встречаемости аллелей АКЛ у европейских сортов овса, а Х.А. Юмагузиной и В.В. Бахтияровой выявлено значительное сходство в спектрах авенинов сортов, обладающих идентичными биологическими и хозяйственно-ценными признаками. Это свидетельствует, что аллели АКЛ или сцепленные с ними локусы отличаются по своей адаптивной и селекционной ценности [9–11]. Однако в литературе практически отсутствуют сведения о связи между частотой встречаемости аллелей АКЛ и природно-климатическими условиями.
Цель исследования : выявление связи между географическим распределением аллелей авенин-кодирующих локусов у образцов овса российской селекции и природно-климатическими факторами для выявления аллелей-маркеров адаптивно-значимых ассоциаций генов.
Материалы и методы исследования. Исследование осуществляли в лаборатории сортовой идентификации семян Государственного аграрного университета Северного Зауралья. Материалом для исследований послужили 39 сортов и линий овса посевного российской селекции различного эколого-географического происхождения (табл. 1, 2).
Таблица 1
|
Сорт |
Происхождение |
Сорт |
Происхождение |
|
Аргумент |
Алтайский край |
Дедал |
Ленинградская обл. |
|
Креол |
-“- |
Борот |
-- |
|
Обыкновенный |
-“- |
Соку |
-- |
|
М 4000 |
-“- |
Песец |
-- |
|
Тулунский 19 |
Иркутская обл. |
Атула |
-- |
|
Кемеровский 90 |
Кемеровская обл. |
Гибрид |
-- |
|
Муром |
-“- |
Успех |
-- |
|
Гаврош |
-“- |
Пушкинский |
-- |
|
Чиж |
Кировская обл. |
Привет |
Московская обл. |
|
Кречет |
-“- |
Борец |
-“- |
|
Эклинс |
-“- |
Краснообский |
Новосибирская обл. |
|
Вятский голозерный |
-“- |
Мутика 556 |
Омская обл. |
|
Першерон |
-“- |
Сибирский кормовой |
-“- |
|
Вятский |
-“- |
Васехадка 298 |
Сахалинская обл. |
|
Траверс |
Красноярский край |
Универсал 1 |
Свердловская обл. |
|
Л 1004-6(479) |
-“- |
Памяти Балавина |
-“- |
|
Голец |
-“- |
Спринт 2 |
-“- |
|
Льговский 9 |
Курская обл. |
Отрада |
Тюменская обл. |
|
Льговский |
-“- |
Талисман |
-“- |
Исследованные образцы овса посевного
Растительный материал предоставлен из коллекции Федерального исследовательского центра Всероссийского института генетических ресурсов растений им. Н.И. Вавилова и НИИСХ Северного Зауралья – филиалом ФИЦ ТюмНЦ СО РАН.
Таблица 2
Характеристика климатических условий в регионах происхождения образцов овса
|
Регион |
СК s zr __ s c[ 05 O_ ^ IS 2 О |
co co л 2 О |
1— о ф ф о о 05 со |
ф в V X СК CD 10 X 05 ф о |
ф 05 . й 05 05 о S 05 Ф о |
05 О О_ о Л с |
5 х 05 > I СЕ 00 И о |
5 X S О о )S CL 2 2 О 5 |
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
|
Кировская обл. |
3775 |
2000 |
105 |
18 |
1 |
31 |
55 |
162,5 |
|
Красноярский край |
3575 |
1400 |
100 |
18 |
-5 |
31 |
60 |
212,5 |
|
Ленинградская обл. |
3775 |
2000 |
105 |
18 |
3 |
29 |
65 |
137,5 |
|
Московская обл. |
3775 |
2000 |
105 |
20 |
3 |
31 |
65 |
137,5 |
Окончание табл. 2
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
|
Свердловская обл. |
3575 |
1400 |
100 |
18 |
0 |
32 |
55 |
162,5 |
|
Тюменская обл. |
3575 |
1400 |
100 |
20 |
0 |
34 |
55 |
162,5 |
|
Алтайский край |
4625 |
2100 |
-250 |
20 |
1 |
34 |
55 |
162,5 |
|
Кемеровская обл. |
3575 |
1400 |
100 |
20 |
-1 |
33 |
60 |
187,5 |
|
Новосибирская обл. |
4625 |
2100 |
-250 |
20 |
1 |
34 |
55 |
162,5 |
|
Омская обл. |
4625 |
2100 |
-250 |
20 |
1 |
34 |
50 |
162,5 |
|
Сахалинская обл. |
4175 |
1500 |
100 |
14 |
-1 |
25 |
75 |
187,5 |
|
Иркутская обл. |
3975 |
950 |
0 |
19 |
-4 |
31 |
55 |
212,5 |
|
Курская обл. |
3775 |
2000 |
105 |
20 |
7 |
33 |
65 |
112,5 |
Примечание: таблица составлена с использованием данных Национального атласа России (URL: .
Для анализа методом случайной выборки отбирали по 100 зерновок каждого сорта. Электрофорез проводили по ранее описанной методике [12]. Идентификацию аллельных вариантов авенин-кодирующих локусов осуществляли по каталогу, описанному В.А. Портянко [9]. В качестве стандарта использовали зерновки овса посевного сорта Астор ( Avn A2 B4 C2 ).
Для поиска групп образцов со схожим аллельным составом применяли кластеризацию методом Варда и методом k-средних. Связь между частотой встречаемости аллелей авенин-кодирующих локусов и природно-климатическими факторами выявляли с применением коэффициента ранговой корреляции Спирмена.
Результаты исследования и их обсуждение. В результате проведения электрофоретического анализа установлено, что 14 образцов были гомогенными по компонентному составу авенина. Остальные образцы содержали от 2 до 9 биотипов. Общее число обнаруженных биотипов составило 114 шт. Для выявления связи между аллелями АКЛ и природно-климатическими факторами необходимо идентифицировать аллели по всем трем локусам для каждого из исследованных биотипов. Однако некоторые из обнаруженных нами блоков компонентов проламина отсутствовали в каталоге генетической номенклатуры, что не позволило идентифицировать контролирующие их аллели АКЛ. Биотипы, для которых не удалось полностью описать генетические формулы авенина, были исключены из дальнейшего исследования. В результате все последующие расчеты проводились на основании данных об аллельном составе 64 биотипов (табл. 3).
Таблица 3
|
Регион происхождения |
Кол-во биотипов |
Регион происхождения |
Кол-во биотипов |
|
Алтайский край |
4 |
Московская обл. |
2 |
|
Иркутская обл. |
3 |
Новосибирская обл. |
2 |
|
Кемеровская обл. |
5 |
Омская обл. |
2 |
|
Кировская обл. |
11 |
Сахалинская обл. |
1 |
|
Красноярский край |
5 |
Свердловская обл. |
6 |
|
Курская обл. |
4 |
Тюменская обл. |
3 |
|
Ленинградская обл. |
16 |
Количество исследованных биотипов с известными генетическими формулами авенина
Селекционная работа с овсом направлена на создание высокоурожайных сортов с высоким содержанием белка в зерне, устойчивых к полеганию, засухе, болезням и обладающих экологиче- ской пластичностью. Некоторые из ассоциаций генов, контролирующих проявление этих признаков, могут быть сцеплены с аллелями АКЛ. Для того чтобы выявить группы регионов, где в про- цессе селекции выводят сорта со схожим аллельным составом АКЛ, нами была проведена кластеризация методом k-средних. Искомое число конечных кластеров (k) было определено проведе- нием предварительной кластеризации методом Варда и равнялось шести. Результаты кластеризации представлены в таблице 4.
Таблица 4
|
Кластер |
Состав (регион происхождения образцов) |
|
1 |
Сахалинская обл. |
|
2 |
Тюменская обл. |
|
3 |
Кировская обл., Красноярский край, Курская обл., Ленинградская обл., Омская обл., Свердловская обл. |
|
4 |
Алтайский край, Новосибирская обл. |
|
5 |
Иркутская обл., Кемеровская обл. |
|
6 |
Московская обл. |
Кластеры совокупностей образцов овса различного происхождения, выделенные методом k-средних
Самую большую группу сформировали образцы овса происхождением из Кировской, Курской, Ленинградской, Омской, Свердловской областей и Красноярского края. Эти регионы находятся на большом расстоянии друг от друга и существенно отличаются по природно-климатическим условиям. Преобладание в сортах овса, созданных в их селекционных центрах, одних и тех же аллелей АКЛ может быть вызвано рядом причин. Во-первых, вовлечением в селекционные программы одних и тех же генотипов. Во-вторых, отбором особей с одинаковым или близким аллельным составом АКЛ. На примере пшеницы доказано, что существуют аллели проламин-кодирующих локусов, маркирующие хозяйственно-ценные признаки, такие как высокие качество зерна и технологические свойства муки. Некоторые из таких аллелей-маркеров достаточно широко распространены и могут с высокой частотой встречаться в сортах, созданных в разных селекционных центрах [13, 14]. По нашему мнению, аллельные варианты АКЛ, встречающиеся с высокой частотой в сортах овса с разным эколого-географическим происхождением, также могут быть маркерами хозяйственно-ценных признаков и представляют интерес для дальнейшего изучения.
Установлено, что некоторые из групп образцов происхождением из разных регионов отличались по частоте встречаемости аллелей АКЛ (рис. 1, 2).
Среди аллелей локуса Avn A самым распространенным был А2 – биотипы с таким вариантом встречались во всех регионах, кроме Сахалинской и Тюменской областей. Аллели А5 и А6 были самыми редкими и обнаружены нами только в образцах происхождением из Новосибирской и Свердловской областей соответственно.
По локусу Avn В наибольшей частотой встречаемости характеризовались аллели В1 и В4 . Вариант В5 встречался реже всего. Он выявлен нами в группах образцов происхождением из Московской и Тюменской областей.
Из аллелей локуса Avn С максимальную частоту встречаемости имели С2 (26,6 %), С3 (20,3 %) и С1 (18,7 %). Реже всего встречались аллели С6a (Ленинградская и Сахалинская обл.) и C6b (Кемеровская обл.).
Одной из причин таких отличий может быть неодинаковая адаптивная ценность аллелей аве-нин-кодирующих локусов. Ряд авторов при изучении проламинов овса отмечали аллели, характерные только для сортов происхождением из конкретных географических регионов [9, 11]. Это указывает на то, что некоторые из аллелей АКЛ могут быть сцеплены с определенными адаптивнозначимыми генными комплексами. В этом случае при отборе генотипов, наиболее приспособленных к определенным природно-климатическим условиям, одновременно происходит и отбор маркирующих их аллелей проламин-кодирующих локусов. В результате частота встречаемости аллелей АКЛ в образцах овса происхождением из разных регионов может существенно различаться.
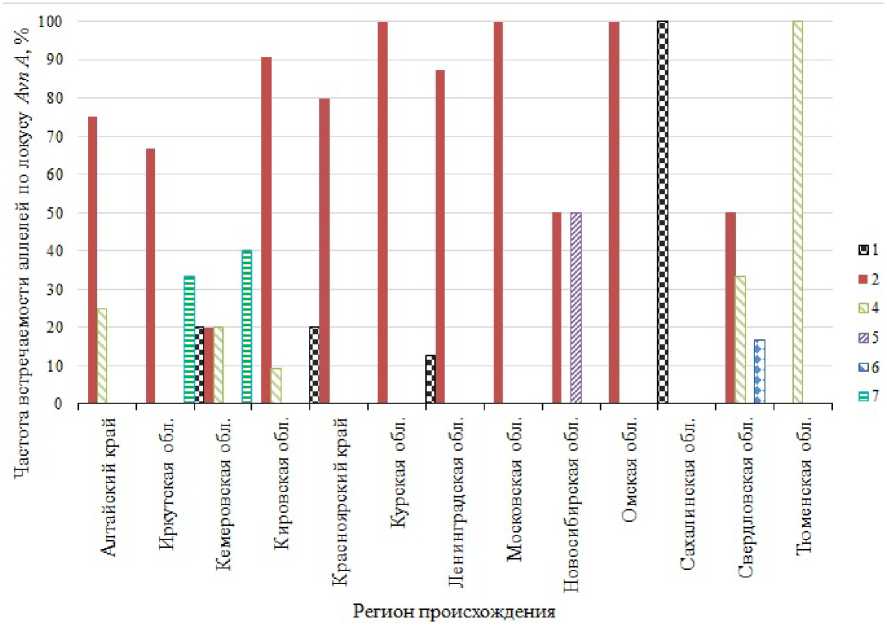
Рис. 1. Частота встречаемости, %, аллелей локуса Avn A в биотипах овса посевного происхождением из различных регионов: 1–7 – номера аллелей
Частота встречаемости аллелей по локусу Avn В, %
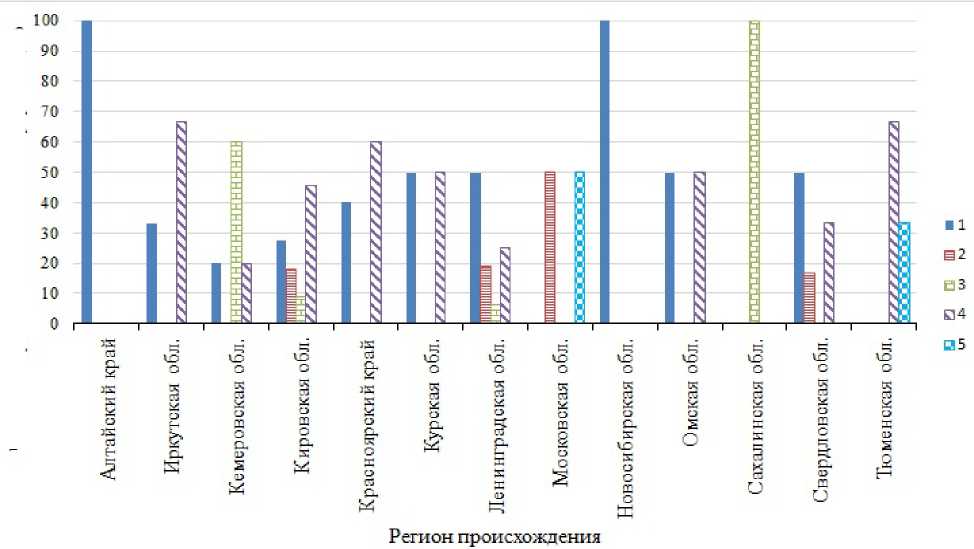
Рис. 2. Частота встречаемости, %, аллелей локуса Avn B в биотипах овса посевного происхождением из различных регионов: 1–5 – номера аллелей
Для того чтобы определить, существует ли авенин-кодирующих локусов и климатическими связь между частотой встречаемости аллелей условиями, описанными в таблице 2, нами был
применен коэффициент ранговой корреляции мости шести аллелей связана определенными Спирмена. Установлено, что частота встречае- природно-климатическими факторами (табл. 5).
Таблица 5
Коэффициент корреляции частоты встречаемости аллелей АКЛ и природно-климатических факторов (связь существенна при p < 0,05)
|
Аллель АКЛ |
СК .4 го к Р 1 s п" ГО ГО 2 >, р сг О ^^ |
1 ^ ^ ф о о 1 X ф 1— |
ф £ i О „ Ф -О Ю 1- О 2 2 § 05 со |
СК 05 05 т >о к ° О 05 - <15 О- СК 05 2 s s 05 ^ о |
ск О аз ° со « § 1 2 ь ® S. d ф CL 2 О ф |
® й 1 2:0 £ н ° | о с[ В 1 2 | н “ |
СК Ого from О |
Ф Z g Ц Т 2 ° О о с о |
|
А1 |
- |
- |
- |
- |
- |
- |
0,71 |
- |
|
В2 |
- |
- |
-0,67 |
- |
- |
- |
- |
- |
|
С3 |
- |
- |
- |
- |
0,67 |
- |
- |
- |
|
С5 |
- |
0,60 |
- |
- |
- |
- |
- |
- |
|
С6 |
- |
- |
-0,74 |
- |
- |
- |
-0,56 |
-0,69 |
|
С6a |
- |
- |
- |
-0,61 |
- |
-0,65 |
0,61 |
- |
Выявлено, что аллель А1 может выступать маркером влаголюбивого генотипа, В2 и С6 – засухоустойчивого. Аллели С3 и С5 перспективны в качестве маркеров теплолюбивых генотипов, а аллель С6а – генотипа, предпочитающего прохладный и влажный климат. Большая часть аллелей, связанных с климатическими факторами, принадлежит локусу Avn C. Как известно, характер белкового полиморфизма неоднороден даже внутри одной маркерной системы. По нашему мнению, аллели локуса Avn C играют наибольшую роль в процессе приспособляемости особей и маркировании генетических систем, детерминирующих выраженность адаптивно-значимых признаков.
Выводы
-
1. Кластеризацией методом k-средних выделено 6 кластеров, объединяющих группы образцов овса посевного разного географического происхождения со схожим аллельным составом АКЛ. Аллели, встречающиеся с высокой частотой в группах сортов, вошедших в один кластер, могут быть маркерами хозяйственно-ценных признаков и представляют интерес для дальнейшего изучения.
-
2. Установлено, что частота встречаемости шести аллелей авенин-кодирующих локусов связана с определенными климатическими условия-
- ми. Аллель А1 может выступать маркером влаголюбивого генотипа, В2 и С6 – засухоустойчивого. Аллели С3 и С5 перспективны в качестве маркеров теплолюбивых генотипов, а аллель С6а – генотипа, предпочитающего прохладный и влажный климат.
-
3. Аллели локуса Avn C, вероятно, играют наибольшую роль в процессе приспособляемости особей и маркировании генетических систем, детерминирующих выраженность адаптивно-значимых признаков.
Список литературы Анализ закономерностей распределения аллелей авенин-кодирующих локусов у сортов овса посевного отечественной селекции
- Романова Ю.А., Губарева Н.К., Конарев А.В. Исследование коллекции вида пшеницы Triticum spelta L. по полиморфизму глиадинов // Генетика. - 2001. - Т. 37, № 9. - С. 1258-1265.
- Перчук И.Н., Конарев А.В., Лоскутов И.Г., Блинова Е.В. и др. Белковые маркеры, морфологические и селекционные признаки в идентификации дублетных образцов культурного овса в коллекциях ВИР (Россия) и нордического генного банка (Nordgen, Швеция) // Тр. по прикладной ботанике, генетике и селекции. - 2016. - Т. 177. - С. 82-93.
- Любимова А.В., Ярова Э.Т., Еремин Д.И. Изменение биотипного состава сортов яровой тритикале в процессе возделывания // Вестн. КрасГАУ. - 2018. - № 5 (140). - С. 3-8.
- Любимова А.В., Еремин Д.И. Изучение генетического разнообразия сортов овса Сибирской селекции по авенин-кодирующим локусам // Агропродовольственная политика России. - 2017. - № 9 (69). - С. 70-74.
- Тоболова Г.В. Изменение биотипного состава сорта мягкой пшеницы Тюменская 80 в процессе семеноводства // Аграрный вестник Урала. - 2009. - № 10. - С. 12-14.
- Новосельская-Драгович А.Ю., Беспалова Л.А., Шишкина А.А., Мельник В.А. и др. Изучение генетического разнообразия сортов мягкой озимой пшеницы по глиадинкодирующим локусам // Генетика. - 2015. - Т. 51, № 3. - С. 324-334.
- Поморцев А.А., Лялина Е.В., Калабушкин Б.А. Гордеин-кодирующие локусы как генетические маркеры у ячменя // Тр. по прикладной ботанике, генетике и селекции. - 2009. - Т. 165. - С. 32-36.
- Якубышина Л.И., Казак А.А., Логинов Ю.П. Использование метода электрофореза в семеноводстве ячменя сорта Одесский 100 // Изв. Оренбург. гос. аграр. ун-та. - 2017. - № 5 (67). - С. 56-59.
- Портянко В.А. Генетический контроль и полиморфизм проламина овса: автореф. дис. … канд. биол. наук. - М., 1987. - 16 с.
- Юмагузина Х.А., Бахтиярова В.В. Электрофоретические спектры спирторастворимых белков зерна видов и подвидов рода Avena // V съезд Всесоюзного общества генетиков и селекционеров им. Н.И. Вавилова (Москва, 24-28 ноября 1987 г.): тез. докл. - М., 1987. - Т. IV. - Ч. 4. - С. 322.
- Souza E., Sorrells M.E. Inheritance and distribution of variation at four avenin loci in North American oat germ plasm // Genome. - 1990. - V. 33, № 3. - P. 416-424.
- Любимова А.В., Еремин Д.И. Региональные особенности полиморфизма авенинов овса, возделываемого в Западной Сибири // Тр. по прикладной ботанике, генетике и селекции. - 2018. - Т. 179, № 2. - С. 85-95.
- Летяго Ю.В., Тоболова Г.В., Белкина Р.И. Оценка сортов мягкой яровой пшеницы по технологическим свойствам и биохимическим признакам // Агропродовольственная политика России. - 2015. - № 5. - С. 64-67.
- Новосельская-Драгович А.Ю., Фисенко А.В., Имашева А.Г. и др. Сравнительный анализ динамики генетического разнообразия по глиадинкодирующим локусам среди сортов озимой мягкой пшеницы Triticum aestivum L., созданных за 40-летний период научной селекции в Сербии и Италии // Генетика. - 2007. - № 43 (11). - С. 1478-1485.
