Анатомические особенности строения мезофилла листьев Oryza sativa и Zizania latifolia (Oryzeae, Poaceae)
")
Автор: Зверева Г.К.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 2, 2025 года.
Бесплатный доступ
Изучение пространственной организации хлоренхимы листьев растений как структурной основы фотосинтеза позволяет охарактеризовать ее более полно. Рассмотрена морфология ассимиляционных клеток и структура мезофилла листовых пластинок и листовых влагалищ генеративных побегов Oryza sativa и Zizania latifolia. Образцы листьев фиксировались в смеси Гаммалунда. Изучение ассимиляционной ткани проводилось с помощью светового микроскопа на мацерированных препаратах, а также на поперечных и продольных срезах листьев. Анатомическое строение рассматривали в средней части листовой пластинки и верхней трети влагалищной части листа. Показано, что хлоренхима листовых пластинок Oryza sativa и Zizania latifolia и листовых влагалищ Oryza sativa сложена из плоских складчатых клеток с небольшим участием клеток более сложной ячеистолопастной, или складчатоячеистой формы. Более усложненные по форме клетки имеют лопастные конфигурации на поперечных срезах и ячеистые, состоящие из двухтрех секций, в продольном направлении. В мезофилле листовых влагалищ Oryza sativa лопастная форма клеток сохраняется, хлоренхима Zizania latifolia состоит преимущественно из клеток простой формы. Строение мезофилла листовых пластинок Oryza sativa и Zizania latifolia характеризуется как упрощенное бамбузоидное. Проведено сопоставление пространственной конфигурации клеток лопастной формы в листьях злаков с бамбузоидным, арундиноидным и фестукоидным типами листовой анатомии. Показано, что эти клетки могут быть как плоскими, так и более сложными, в основном ячеистолопастными.
Типы листовой анатомии злаков, хлоренхима, клетки сложной формы, лопастные клетки
Короткий адрес: https://sciup.org/147250947
IDR: 147250947 | УДК: 581.823:582.52/.59 | DOI: 10.17072/1994-9952-2025-2-132-142
Anatomical features of the leaf mesophyll structure Oryza sativa and Zizania latifolia (Oryzeae, Poaceae)
The study of the spatial organisation of plant leaf chlorenchyma, as the structural basis of photo-synthesis, allows us to characterize it more fully. The morphology of assimilative cells and the structure of the mesophyll of leaf blades and leaf sheaths of generative shoots of Oryza sativa and Zizania latifolia are consid-ered. Leaf samples were fixed in Gammalund's mixture. The assimilative tissue was studied using a light micro-scope on macerated preparations, as well as on transverse and longitudinal sections of leaves. The anatomical structure was considered in the middle part of the leaf blade and the upper third of the sheath part of the leaf. It is shown that the chlorenchyma of the leaf blades of Oryza sativa and Zizania latifolia and the leaf sheaths of Ory-za sativa is composed of flat folded cells with a small participation of cells of a more complex cellular-lobed or folded-cellular shape. More complicated in shape, the cells have lobed configurations on the cross sections and cellular, consisting of two or three sections, in the longitudinal direction. In the mesophyll of the leaf sheaths of Oryza sativa, the lobed shape of the cells is preserved, the chlorenchyma of Zizania latifolia consists mainly of cells of simple shape. The structure of the mesophyll of the leaf blades of Oryza sativa and Zizania latifolia is characterized as simplified bambusoid. The spatial configuration of lobed cells in leaves of grasses with bam-busoid, arundinoid and festucoid types of leaf anatomy was compared. It is shown that these cells can be both flat and more complex, mainly cellular-lobed.
Текст научной статьи Анатомические особенности строения мезофилла листьев Oryza sativa и Zizania latifolia (Oryzeae, Poaceae)
Изучение пространственной организации хлоренхимы листьев у разных групп злаков позволяет более полно охарактеризовать их фототрофную ткань как структурную основу фотосинтеза.
Oryza sativa L. и Zizania latifolia (Griseb.) Hance ex F.Muell. являются представителями трибы Oryzeae Dumort., которая по современным классификациям относится к подсемейству Oryzoideae Kunth ex Beilschm. ( Ehrhartoideae Link) семейства Poaceae Barnhart [Цвелев, 2006; Soreng et al., 2015; Цвелев, Пробатова, 2019]. В мезофилле листьев как трибы Oryzeae, так и в целом подсемейства Oryzoideae широко распространены клетки сложной лопастной формы (arm cells) с выростами и складками по краю [Tateoka, 1963; Renvoize, 1985; Ellis, 1987; Leandro, Scremin-Dias, Arruda, 2016; Watson, Macfarlane, Dallwitz, 2024 и др.], более подробно они исследованы у представителей рода Oryza L.
На примере Oryza sativa L. показано, что лопастные клетки мелкие, плотно упакованные, число выступов в них изменяется от 3 до 12 и возрастает от нижних листовых пластинок к верхним, это приводит к увеличению отношения клеточной поверхности к ее объему, на тангентальных сечениях они имеют вид тонких овалов [Chonan, 1967, 1978]. На основании особенностей проекций, числа зеленых пластид и характера их расположения у риса выделяют около 15 основных типов ассимиляционных клеток [Бурунду-кова и др., 2003]. Считается, что архитектура хлоренхимы листьев риса, связанная с высокой густотой устьиц, а также с разветвленностью и малым размером клеток в сочетании с плотно прижатыми хлоропластами к их периферии и покрытию ими более 95% клеточной поверхности, способствует компенсации высокого фотодыхательного потенциала и усилению фотосинтетической активности в более теплых условиях [Tsunoda, 1984; Evans, Loreto, 2000; Sage, Sage, 2009].
Для листьев гибридных линий риса отмечали, что усиление плотности лопастных клеток и развитости их долей приводит к увеличению проводимости мезофилла для СO 2 и возрастанию уровня фотосинтеза [Adachi et. al., 2013; He et al., 2017]. На примере листьев 50 сортов Oryza sativa также показано, что интенсивность фотосинтеза растений высоко коррелирует с индексом поверхности наружных мембран этих клеток (ИМК) и числом хлоропластов в единице листовой поверхности [Бурундукова, Холупенко, 2017]. Созданные с помощью электронной и световой микроскопии трехмерные модели клеток мезофилла Ory-za sativa выглядят как эллипсоидные диски с неровными противоположными поверхностями и несколькими лопастями по периферии [Oi et al., 2017, 2020].
Для мезофилла листьев рода Zizania L. также характерно присутствие клеток лопастной формы [Metcalfe, 1960; Watson, Macfarlane, Dallwitz, 2024]. При описании анатомического строения листьев Zizania latifolia в основном обращают внимание на толстую кутикулу на обеих поверхностях, развитие воздухоносных полостей и фузоидных клеток, а также на особенности строения сосудисто-волокнистых пучков [Yang et al., 2014].
Для подсемейства Oryzoideae характерен бамбузоидный тип листовой анатомии, отличающийся, в частности, утолщенной клеточной оболочкой паренхимной обкладки пучков с небольшим числом хлоропластов и не радиально расположенной ассимиляционной тканью, состоящей из продолговатых или равносторонних лопастных клеток [Brown, 1958; Carolin, Jacobs, Vesk, 1973].
Задачей настоящей работы было на основе сравнительного исследования листьев Oryza sativa и Zizania latifolia как представителей трибы Oryzeae более подробно рассмотреть морфологию ассимиляционных клеток и пространственную организацию мезофилла листьев.
Материалы и методы исследования
Исследована структура хлорофиллоносной паренхимы листовых пластинок и листовых влагалищ Oryza sativa и Zizania latifolia, расположенных в средней части генеративных побегов, находящихся в состоянии цветения. Основные формы ассимиляционных клеток и их расположение в листовом про- странстве изучены с помощью светового микроскопа на мацерированных препаратах, а также на поперечных и продольных срезах листьев, фиксированных в смеси Гаммалунда [Possingham, Saurer, 1969; Гродзинский, Гродзинский, 1973]. Продольные срезы осуществлялись как параллельно нижней листовой поверхности (парадермальный срез), так и перпендикулярно к ней (продольный боковой срез). Анатомическое строение рассматривали в средней части листовой пластинки и верхней трети влагалищной части листа. Высота и ширина клеток измерялись на поперечных срезах листьев, толщина – на продольных срезах. Образцы Oryza sativa отбирались в оранжерее Сибирского ботанического сада Томского государственного университета, Zizania latifolia – в пойме р. Аргунь в окрестностях с. Аргунск Нерчинско-Заводского р-на Забайкальского края.
При описании формы ассимиляционных клеток использовали усовершенствованную нами классификацию [Зверева, 2009, 2011], при которой сложные клеточные проекции, характеризующиеся присутствием выраженных складок и выростов, разделялись на ячеистые (состоящие из клеточных ячеек или секций, похожих на палисадные клетки) и лопастные (с удлиненными или округлыми выростами). Ячеистые клетки расположены вдоль листа, лопастные клетки проявляются на поперечных срезах. Клетки сложной формы могут быть как плоскими (наличие выростов и складок в одной плоскости), так и более усложненными (наличие выростов и складок в двух-трех направлениях), при этом чаще встречаются ячеисто-лопастные клетки, у которых лопастные проекции проявляются на поперечных срезах, а ячеистые – на продольных сечениях.
При определении числа клеток в единице площади листа использовали подходы, предложенные Т.К. Горышиной [1989]. Количественно-анатомические признаки обработаны статистически общепринятыми методами с помощью программы Snedecor [Сорокин, 2004], сравнение данных осуществлялось с помощью t-критерия Стьюдента, в таблицах показаны средние значения и стандартные ошибки. Названия видов растений приведены по базе World Flora Online [World Flora Online: эл. ресурс].
Результаты и их обсуждение
Oryza sativa и Zizania latifolia относятся к световым гигрофитам, листья Zizania latifolia более толстые и отличаются присутствием фузоидных клеток (табл. 1). Листовые пластинки у обоих злаков амфистома-тические, с многочисленными устьицами, слабым развитием склеренхимы и крупными моторными клетками на верхней стороне, они близки по толщине эпидермы и ее наружной стенки, утолщение которой составляет 28–35% (рис. 1) . Во влагалищной части листьев имеются крупные воздухоносные полости, у Zizania latifolia они нередко заполнены бесцветными клетками с вытянутыми отростками. Верхняя треть влагалищ толще листовых пластинок в 3.2–3.8 раза и характеризуется сильно утолщенными наружными стенками абаксиальной эпидермы, особенно у Zizania latifolia (рис. 2) .
Таблица 1
Количественно-анатомическая характеристика листьев злаков трибы Oryzeae на поперечном срезе [Quantitative-anatomical characterization of the leaves of grasses of the Oryzeae tribe on a cross-section]
|
Вид |
Толщина, мкм |
||||
|
эпидермы |
наружной стенки эпидермы |
листа в области проводящих пучков |
|||
|
адаксиальнойп |
абаксиальной |
адаксиальной |
абаксиальной |
||
|
Листовая пластинка |
|||||
|
Oryza sativa |
12.2±0.42 |
11.5±0.23 |
4.3±0.20 |
4.0±0.15 |
88.1±1.61 |
|
Zizania latifolia |
11.2±0.25 |
12.5±0.43 |
3.8±0.20 |
3.5±0.18 |
120.9±1.70 |
|
Листовое влагалище |
|||||
|
Oryza sativa |
14.0±0.85 |
13.0±0.40 |
2.5±0.18 |
5.1±0.20 |
284.0±12.8 |
|
Zizania latifolia |
10.2±0.30 |
11.0±0.58 |
2.2±0.10 |
6.6±0.35 |
456.2±30.2 |
Длина замыкающих клеток устьиц у обоих злаков достоверно возрастает от пластинок к влагалищам (табл. 2). Более мелкие устьица у Oryza sativa , лишь на верхней поверхности влагалищной части листьев они близки по размерам у обоих видов. Отметим, что в листовых влагалищах устьица встречаются преимущественно на нижней стороне и значительно реже – на адаксиальной эпидерме . Вокруг проводящих пучков расположены клетки паренхимной обкладки, имеющие округлые или овальные очертания на поперечных срезах и вытянутые в виде полосок формы – в продольном направлении. У рассматриваемых злаков они достаточно короткие и отличаются отсутствием или слабым содержанием хлоропластов.
В листовых пластинках обоих видов число слоев мезофилла составляет 6–8 в области сосудистоволокнистых пучков и 3–4 – в области моторных клеток. На поперечных срезах клетки мезофилла мелкие, плотно сомкнутые и имеют лопастную, или складчатую форму. Ассимиляционные клетки, опирающиеся на верхнюю и нижнюю эпидермы, у Oryza sativa близки к изодиаметрическим, а у Zizania latifolia их высота в среднем превышает ширину в 1.2 раза (табл. 3). Глубина рассечения этих клеток со стороны эпидерм более равномерная у Oryza sativa и составляет 16–18% от их высоты (табл. 4). У Zizania latifolia более сильно рассечены клетки под адаксиальной эпидермой. В глубине листовых пластинок между паренхимными обкладками соседних проводящих пучков у обоих злаков протягиваются в 1.4–1.8 раза более широкие клетки по сравнению с расположенными субэпидермально.
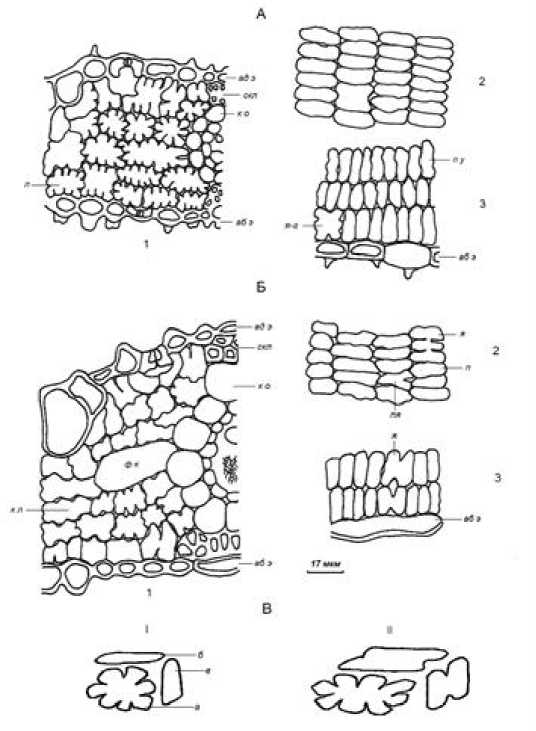
Рис. 1. Анатомическое строение ассимиляционной ткани листовых пластинок Oryza sativa (А) и Zizania latifolia (Б)
Срез: 1 – поперечный; 2 – парадермальный у абаксиальной эпидермы; 3 – продольный боковой у абаксиальной эпидермы. ад э – адаксиальная эпидерма; аб э – абаксиальная эпидерма; скл – склеренхима; м к – моторные клетки, к о – клетки паренхимной обкладки, ф к – фузоидные клетки. Форма проекций ассимиляционных клеток: л – лопастная; п у – простая удлиненная; я – ячеистая; пя – полу-ячеистая; я-г – ячеисто-губчатая. В – пространственная форма клетки: I – плоская лопастная; II – ячеисто-лопастная. Срез: а – поперечный; б – парадермальный; в – продольный боковой
[The anatomical structure of the assimilative of leaf blades of Oryza sativa (A) and Zizania latifolia (Б) Section: 1 - transverse; 2 - paradermal at the abaxial epidermis; 3 - longitudinal lateral at the abaxial epidermis. ад э - adaxial epidermis; аб э - abaxial epidermis; скл - sclerenchyma; м к - bulliform cells, к о -cells of bundle sheath; ф к - fusoid cells. The shape of the projections of assimilatory cells: л - lobed; п у - simple elongated; я - cellular; пя - semi-cellular; я-г - cellular-spongy. B - spatial shape of the cell: I -flat lobed; II - cellular-lobed. Section: а - transverse; б – paradermal; в - longitudinal lateral]
Во влагалищной части листьев хлоренхима сосредоточена у нижней поверхности, в 2–4 слоя у проводящих пучков и в 1–2 слоя между ними, и состоит преимущественно из клеток, ширина которых в 1.2– 1.5 раза больше высоты, при этом у Oryza sativa сохраняется их лопастная форма, а для клеточных проекций Zizania latifolia в подавляющем большинстве характерны простые овальные конфигурации.
На продольных сечениях листовых пластинок обоих злаков и влагалищной части листьев риса ассимиляционные клетки выглядят в виде плотно расположенных удлиненных узких овалов, часто с небольшой волнистостью боковых стенок, особенно это проявляется у Oryza sativa. На парадермальных срезах они перпендикулярны к клеткам паренхимной обкладки, на продольных боковых – к эпидермам. Такие клетки можно описать как плоские лопастные, или плоские складчатые. Но наряду с простыми вытянутыми проекциями в одном ряду встречаются и более усложненные, имеющие выемчатую, полуячеистую, слабо ячеистую или реже хорошо выраженную ячеистую форму, состоящую из двух или очень редко трех секций. Эти проекции более толстые, их рассеченность на тангентальных срезах листьев Oryza sativa составила 10–15% от высоты. В листовых пластинках Zizania latifolia более часты простые или выемчатые утолщенные проекции. В целом, такие клетки имеют более сложное пространственные формы и их можно охарактеризовать как выемчато-лопастные или ячеисто-лопастные.
Хлорофиллоносная паренхима листовых влагалищ Zizania latifolia представлена клетками простой формы, среди которых можно выделить плоские и более утолщенные.
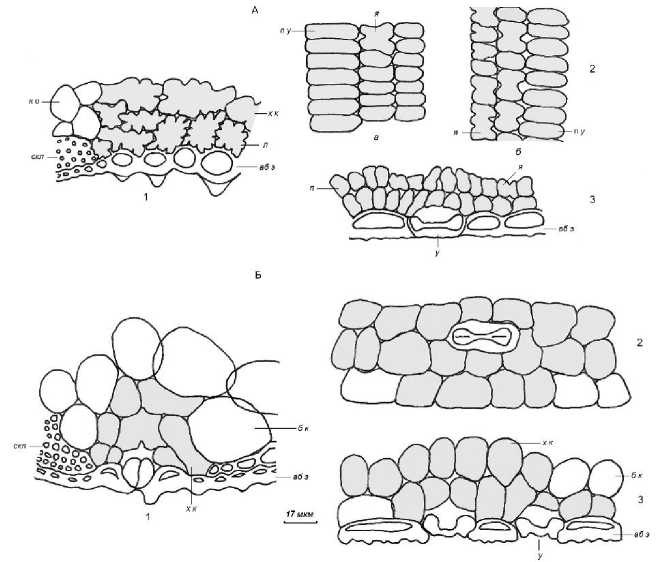
Рис. 2. Расположение ассимиляционных клеток в листовых влагалищах Oryza sativa (А) и Zizania latifolia (Б)
Срез у абаксиальной эпидермы: 1 – поперечный; 2 – парадермальный ( а – у абаксиальной эпидермы; б – в середине листа); 3 – продольный боковой. Клетки мезофилла: х к – хлорофиллоносные, б к – бесцветные. у – устьице, п – простая форма проекций. Остальные обозначения см. рис. 1
[The location of assimilative cells in leaf sheaths Oryza sativa (А) и Zizania latifolia (В) Section at the abaxial epidermis: 1 – transverse; 2 – paradermal; 3 – longitudinal lateral. Paradermal section: a – at the abaxial epidermis; б – in the middle of the leaf. Mesophyll cells: х к – chlorophyll-bearing cells, б к – colourless; у – the stoma. Other designations see Fig. 1]
Таблица 2
Количественно-анатомическая характеристика листьев злаков трибы Oryzeae на продольном срезе, мкм [Quantitative-anatomical characterization of the leaves of grasses of the Oryzeae tribe on a longitudinal section, µm]
|
Вид |
Длина замыкающих клеток устьиц на эпидерме |
Размеры клеток паренхимной обкладки |
||
|
адаксиальной |
абаксиальной |
длина |
ширина |
|
|
Листовая пластинка |
||||
|
Oryza sativa |
21.8±0.30 |
21.7±0.20 |
32.4±1.77 |
11.9±0.33 |
|
Zizania latifolia |
23.5±0.26 |
24.5±0.26 |
45.8±2.63 |
20.9±0.58 |
|
Листовое влагалище |
||||
|
Oryza sativa |
29.6±0.85 |
24.9±0.28 |
31.1±1.27 |
19.5±0.43 |
|
Zizania latifolia |
28.4±0.47 |
27.6±0.28 |
42.0±3.85 |
13.7±0.46 |
Уплощенная форма ассимиляционных клеток, упакованных в стопки, приводит к высокой насыщенности ими единицы площади листа. Так, в листовых пластинках Oryza sativa содержится 4.04 млн/см2 клеток, что близко к данным, полученными ранее [Бурундукова, 1993; Бурундукова и др., 1993], в листовых влагалищах их в 4.5 раза меньше. Плотность ассимиляционных клеток в листовых пластинках Zizania latifolia – 4.56 млн/см2, во влагалищной части листьев ввиду укрупнения клеток она резко снижается и составляет 0.89 млн/см2. Считается, что при небольшой толщине листьев подобное расположение ассимиляционных клеток с многочисленными боковыми выростами создает большую ассимиляционную поверхность и систему воздухоносных каналов, необходимую для интенсивного газообмена [Бурундукова, 1993].
Таблица 3
Размеры клеток мезофилла в листьях злаков трибы Oryzeae
[The sizes of mesophyll cells in the leaves of grasses of the Oryzeae tribe]
|
Вид |
Размеры клеток мезофилла, мкм |
|||
|
высота |
ширина |
толщина |
||
|
плоских \ |
утолщенных |
|||
|
Листовая пластинка |
||||
|
Первый ряд у адаксиальной эпидермы |
||||
|
Oryza sativa |
17.4±0.62 |
15.5±0.58 |
6.7±0.51 |
13.4±0.25 |
|
Zizania latifolia |
17.4±0.38 |
14.0±0.51 |
9.8±0.23 |
16.7±0.91 |
|
В центре у паренхимной обкладки |
||||
|
Oryza sativa |
16.9±0.60 |
24.0±0.95 |
7.7±0.62 |
14.4±0.91 |
|
Zizania latifolia |
18.7±0.92 |
21.6±0.77 |
8.8±0.28 |
15.9±0.93 |
|
Первый ряд у абаксиальной эпидермы |
||||
|
Oryza sativa |
15.7±0.36 |
16.2±0.51 |
7.2±0.25 |
12.0±0.43 |
|
Zizania latifolia |
16.2±0.60 |
13.8±0.86 |
8.4±0.32 |
15.7±0.95 |
|
Листовое влагалище |
||||
|
В центре у паренхимной обкладки |
||||
|
Oryza sativa |
18.4±0.78 |
29.6±2.24 |
13.9±1.38 |
18.4±1.67 |
|
Zizania latifolia |
32.1±0.85 |
41.3±1.02 |
20.9±0.61 |
36.4±2.32 |
|
Первый ряд у абаксиальной эпидермы |
||||
|
Oryza sativa |
15.0±0.36 |
22.6±0.85 |
9.8±0.29 |
17.3±0.28 |
|
Zizania latifolia |
21.9±0.60 |
26.4±1.56 |
15.9±0.92 |
30.2±1.75 |
|
Примечание: высота и ширина определены на поперечных срезах, толщина – на продольных срезах. |
||||
Таблица 4
Размеры лопастей клеток мезофилла первого ряда на поперечных срезах листовых пластинок злаков трибы Oryzeae , мкм
[Sizes of the lobes of the first row mesophyll cells on cross sections of leaf blades of grasses of the Oryzeae tribe, µm]
|
Вид |
Клетки у эпидермы (первый ряд) |
|||
|
адаксиальной |
абаксиальной |
|||
|
глубина рассечения со стороны эпидермы |
ширина выступов у эпидермы |
глубина рассечения со стороны эпидермы |
ширина выступов у эпидермы |
|
|
Oryza sativa |
2.9±0.21 |
6.3±0.28 |
2.8±0.27 |
5.5±0.50 |
|
Zizania latifolia |
3.7±0.33 |
7.5±0.25 |
2.5±0.23 |
7.2±0.26 |
Таким образом, хлоренхима листовых пластинок Oryza sativa и Zizania latifolia и листовых влагалищ Oryza sativa состоит из плоских складчатых клеток с небольшим участием клеток более сложной ячеисто-лопастной, или складчато-ячеистой формы. В листовых влагалищах Zizania latifolia мезофилл сложен из крупных клеток простой овальной формы.
Наличие двух форм клеток мезофилла в листьях Oryza sativa было описано ранее [Chonan, 1970], первая из которых, более многочисленная, имеет лопастные контуры на поперечных срезах и удлиненные овалы – на продольных сечениях, во второй, редко встречающейся, в этих направлениях сочетаются лопастные и слабо ячеистые очертания. Но в дальнейшем при описании хлоренхимы и детальном составлении моделей клеток и мезофилла трехмерно более сложные клетки не отмечались [Oi et al., 2017; Ouk, Oi, Taniguchi, 2020; Ouk et al., 2022]. Клетки хлоренхимы видов Oryza в основном описывают как вытянутые или сплюснутые сфероиды с неровными вогнуто-выпуклыми поверхностями [Sca-faro et al., 2011; Oi et al., 2020; Ouk et al., 2022; Yamane, Oi, Taniguchi, 2022]. Расщепление лопастной клетки на два сегмента в продольном направлении показано лишь в одной из многочисленных представленных 3D-моделей для Oryza sativa [Oi, 2023]. При изучении анатомии листьев у 24 видов Oryza было выделено два типа клеток мезофилла по степени развитости лопастей: без долек на клеточной стенке и с их наличием, и на этой основе построены модели их ассимиляционной ткани [Chatterjee et al., 2016].
Из клеток лопастной формы (arm cells) сложен также мезофилл листьев злаков подсемейства Bam-busoideae [Calderon, Soderstrom, 1973; Soderstrom, Ellis, 1988; Vieira et аl., 2002 и др.]. На примере листьев Phyllostachys reticulata (Rupr.) K.Koch и Pseudosasa japonica (Siebold & Zucc. ex Steud.) Makino ex Nakai нами показано, что лопастные клетки, выявляющиеся на поперечных срезах, могут быть как плоскими, так и утолщенными, последние отличаются преимущественно ячеистыми проекциями в продольном направлении [Зверева, 2011]. У Phragmites australis (Cav.) Steud., как представителя арундиноидного строения, в клетках хлоренхимы, занимающих внутреннее пространство листа между сосудистоволокнистыми пучками, сочетаются разнообразные лопастные очертания на поперечных срезах и овальные или реже ячеистые конфигурации из двух-трех секций на тангентальных сечениях [Зверева, 2023]. Хорошо выраженные или слабо лопастные ассимиляционные клетки нередко встречаются и в листьях фестукоидных злаков, особенно увлажненных местообитаний [Зверева, 2009, 2011], при этом они могут быть как плоскими, так и ячеисто-лопастными в основном с 2–4 секциями на тангентальных срезах. По сравнению с фестукоидными злаками-гигрофитами плоские лопастные клетки Oryza sativa и Zizania lati-folia, расположенные в глубине листа около проводящих пучков, близки по высоте, но меньше по ширине в 1.2–1.6 раза, а по толщине – в 1.8–2.6 раза.
Таким образом, группа клеток лопастной формы, более или менее многочисленная, проявляется в листьях злаков с фестукоидным, арундиноидным и бамбузоидным типами листовой анатомии, при этом выявляются как плоские, так и более усложненные ячеисто-лопастные клетки, что может свидетельствовать об общих чертах построения их хлоренхимы.
Заключение
Ассимиляционная паренхима листовых пластинок Oryza sativa и Zizania latifolia состоит из мелких плоских складчатых клеток с небольшим участием утолщенных выемчато-лопастных и ячеистолопастных. По сравнению с бамбуками, хлоренхима листьев которых сложена из более разнообразных по трехмерной форме складчатых клеток, строение мезофилла у Oryza sativa и Zizania latifolia можно рассматривать как упрощенное бамбузоидное, что, вероятно, способствует усилению специализации листа для функционирования в условиях повышенной влажности и часто интенсивной инсоляции.
В листовых влагалищах Oryza sativa однообразное и упрощенное строение мезофилла сохраняется, а у Zizania latifolia в этой части листа хлоренхима состоит преимущественно из клеток простой формы.
Группа лопастных клеток, проявляющаяся своими складчатыми проекциями на поперечных срезах, в той или иной степени развитости наблюдается в листьях фестукоидных злаков, более выраженные и многочисленные их формы характерны для представителей арундиноидного и бамбузоидного типов листовой анатомии. Как в листьях Oryza sativa и Zizania latifolia, так и в целом в листьях С 3 -злаков наряду с плоскими формами лопастных клеток выявляются и более сложные, в основном, ячеисто-лопастные.
