Анатомическое строение побега Juniperus polycarpos С. Koch в предгорном Дагестане

Автор: Асадулаев З.М., Садыкова Г.А., Рамазанова З.Р.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 3, 2020 года.
Бесплатный доступ
Идентификация видов J. excelsa Bieb. и J. polycarpos C. Koch проблематична и требует проведения дополнительных исследований. Настоящая статья посвящена описанию и количественной оценке анатомических признаков побегов J. polycarpos, произрастающего в условиях Предгорного Дагестана, которые могут быть использованы для решения таксономических вопросов. Модельные побеги (1-3-летние) J. polycarpos собирали с северной стороны кроны на уровне 1.5 м от земли. Фиксирование, приготовление временных микропрепаратов и описание основных структурных элементов тканей побега проводили в соответствии с общепринятыми методиками. Впервые дано описание анатомического строения молодых побегов J. polycarpos. Проведена их количественная оценка, выделены характерные анатомические особенности в зависимости от возраста и формы побега. Специфическими особенностями морфологического строения можно считать наличие двух типов побегов (двухлопастных и трехлопастных); анатомического - наличие у двухлопастных побегов в паренхиме листовых подушек большого количества каменистых клеток, а также характерное строение центрального цилиндра; наличие и количество масляных клеток. Выявленные признаки могут быть использованы для сравнительно-анатомической идентификации видов рода Juniperus.
Juniperus polycarpos c. koch, двухлопасной побег, трехлопасной побег, ткани побега, цен-тральнопредгорный дагестан (губденская популяция)
Короткий адрес: https://sciup.org/147229650
IDR: 147229650 | УДК: 581.8 | DOI: 10.17072/1994-9952-2020-3-157-164
Shoot anatomical structure Juniperus polycarpos C. Koch in foothill Dagestan
Identification of the species J. excelsa L. and J. polycarpos C. Koch is problematic and requires additional research. This article is devoted to the description and quantification of the anatomical features of the shoots of J. polycarpos, which grows in the conditions of Piedmont Dagestan to solve taxonomic issues. Methods Model shoots (1-3 year old) J. polycarpos was collected from the north side of the crowns at a level of 1.5 m from the ground to study the anatomical structure, fixing, preparing temporary micropreparations and describing the main structural elements of shoot tissues was carried out in accordance with generally accepted methods. Results. The anatomical structure of young shoots of J. polycarpos is described for the first time in order to identify taxon-specific and adaptive characters when studying the anatomical structure of the stem of J. excelsa and other populations of J. polycarpos. Their quantitative assessment is given, characteristic anatomical features are identified depending on the age and shape of the shoot. Conclusion The specific features of their morphology can be considered the presence of two types of shoots (two-bladed and three-bladed); anatomy - the presence in the parenchyma of leaf pillows in two-bladed shoots of a large number of stony cells, as well as structural features of the central cylinder; the presence and quantity of oil cells. Identified characters can be used for comparative anatomical identification of species of the genus Juniperus.
Текст научной статьи Анатомическое строение побега Juniperus polycarpos С. Koch в предгорном Дагестане
Систематика рода Juniperus L. основана на признаках морфологического и анатомического строения вегетативных органов [Комаров, 1934; Hall, 1961], морфологии шишек, биохимических данных [Gadek, Quinn, 1984; Farjon, 1992, 2001;
Adams, 2008; Adams et al., 2016], экологических и ботанико-географических исследованиях [Исмаилов, 1975], и неоднократно пересматривалась.
Особенно много вопросов возникает при идентификации видов Juniperus excelsa Bieb. и J. poly-
carpos C. Koch, имеющих сходные морфологические признаки, но разные ареалы и экологию.
J. polycarpos считают ксерофитом, он встречается в Закавказье, Турции, Иране, а на территории России – в Дагестане [Кецховели, 1980]. J. excelsa – мезоксерофит с более широким ареалом, охватывающим Балканский полуостров, острова Греческого архипелага, Малую Азию, Иран, Крым (от Балаклавы до Карадага); на Северо-Западном Кавказе встречается от Анапы до Геленджика [Ареалы …, 1977].
Оба упомянутых вида древовидных можжевельников по причине их реликтовости и сокращающихся ареалов включены в разные Красные книги: J. polycarpos занесен в Красную книгу Дагестана [Красная…, 2009], J. excelsa – в Красную книгу России [Красная…, 2008].
В Конспекте флоры Кавказа [2003] J. polycarpos приводится в ранге подвида ( J. excelsa subsp. polycarpos ), тогда как другими авторами рассматривается в качестве самостоятельного вида [Галушко, 1978; Муртазалиев, 2009; Adams et al., 2016; Hojjati et al., 2018 и др.]. Результаты современных молекулярно-генетических исследований [Adams et al., 2016] также свидетельствуют об обособленности J. polycarpos от J. excelsa и подтверждают его видовой статус.
Но тем не менее, таксономическое положение J. polycarpos остается дискуссионным и требует проведения дополнительных исследований.
В связи с этим была поставлена задача сравнительной оценки J. polycarpos и J. excelsa по анатомическим признакам их вегетативных и генеративных органов.
В данной статье представлено описание и количественная оценка анатомических признаков побегов J. polycarpos , произрастающего в условиях Предгорного Дагестана .
Материал и методы исследований
Объектом исследования послужили 1–3-летние побеги J. polycarpos , собранные в 2019 г. на южных отрогах хр. Чонкатау в окрестности с. Губден Карабудахкенского р-на Предгорного Дагестана на высоте 800 м над ур. м. Модельные побеги (1–3летние) собирали с северной стороны кроны на уровне 1.5 м от земли. Их фиксировали в 70%-ном растворе спирта с глицерином. Приготовление временных микропрепаратов проводили по общепринятой методике анатомических исследований [Барыкина, 2004]. Описание основных структурных элементов тканей листа проводили в соответствии с разработками И.А. Самылиной, О.Г. Аносовой [2007].
Анатомические исследования проведены в Лаборатории интродукции и генетических ресурсов древесных растений ГорБС ДФИЦ РАН на оптиче- ском микроскопе Levenhuk D870T с использованием окуляр-микрометра. Микропрепараты фотографировали на микроскопе Ломо–АТ 054 и с помощью оптического видеоокуляра DCM 510 SCOP. Статистическая обработка полученных данных выполнена с использованием компьютерной программы Microsoft Excel.
Схемы поперечных сечений побегов J. polycarpos , представленных на рис. 1 и 2, даны авторами на основе анализа анатомических срезов.
Результаты и их обсуждение
У Juniperus polycarpos C. Koch побеги имеют супротивное и мутовчатое листорасположение, которым на поперечном срезе стебля соответствуют двух- и трехлопастное строение (рис. 1). При этом число и размеры лопастей связывают как с особенностями филлотаксиса, так и с неравномерным разрастанием разных участков стебля и оснований листьев [Лотова, 1987]. По мере утолщения стебля вследствие разрастания ксилемы к концу второго года роста выемки между лопастями распрямляются.
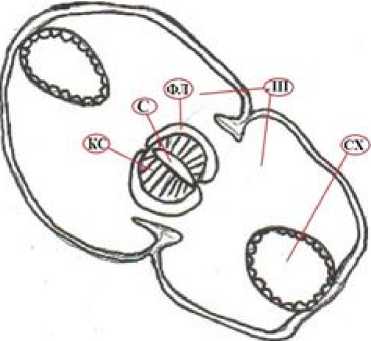
Рис. 1 . Поперечный срез двухлопастного однолетнего побега:
СХ – смоляной ход; ЛП – листовые подушки; ФЛ – флоэма; КС – ксилема; С – сердцевина; СЛ – сердцевинные лучи; ЭНД – эндодермоидный слой; ПР – перидерма
Несмотря на внешнее морфологическое различие, связанное с лопастностью (рис. 2), общее анатомическое строение и взаиморасположение тканей в стебле побегов при этом однотипное. Кутикула стебля хорошо развитая, с толщиной от 10.5 до 13.5 мкм (в зависимости от возраста). Эпидерма однослойная, сохраняется и на трехлетних побегах. Высота клеток колеблется от 7.5 мкм до 20.0 мкм, при этом стенки клеток с возрастом становятся более толстыми и более плоскими (табл. 1).
Волоски на стебле встречаются редко, они простые, серповидно изогнутые. Между клетками эпидермы четко выделяются двухэтажные устьица.
Под эпидермой прерывистыми рядами расположена 1–2-слойная гиподерма с толстыми, сильно лигнифицированными клеточными стенками. Толщина ее варьирует в зависимости от числа слоев от 14.5 мкм до 25.3 мкм у трехлопастных и 15.3 мкм до 22.8 мкм ‒ у двухлопастных побегов. Первичная кора одно- и двулетних побегов содержит хлорофиллоносную, связанную с листовыми по- душками паренхиму, и имеет 3–4 слоя клеток. Внутренний слой первичной коры, окружающий центральный цилиндр, также четко выражен и называется эндодермоидным [Эзау, 1980]. Толщина этого слоя у трехлопасных побегов на второй год жизни увеличивается от 31.3 мкм до 39 мкм, у трехлетних побегов, наоборот, уменьшается до 32.8 мкм.
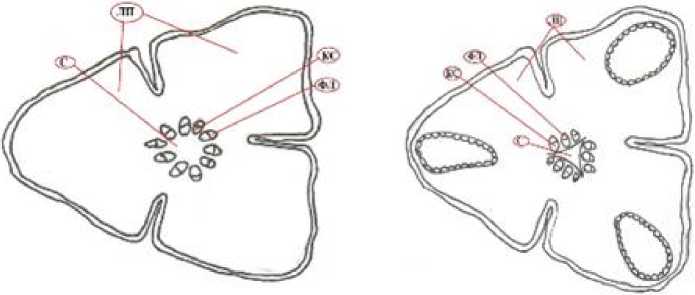
А (однолетний без смоляных ходов)
Б (однолетний со смоляными ходами)
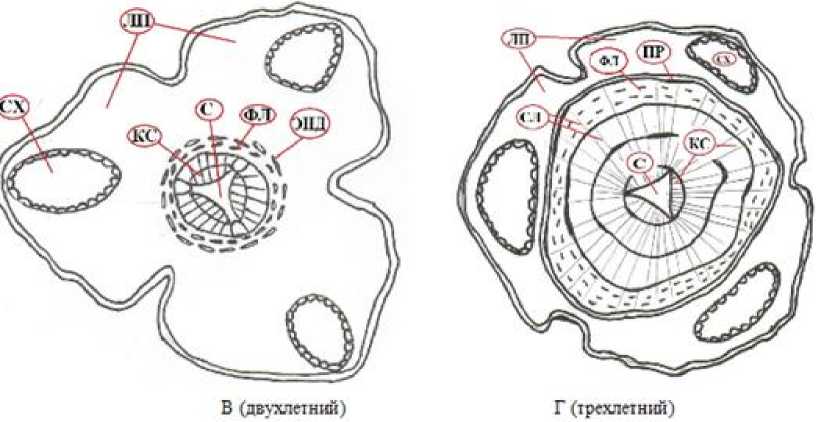
Рис. 2 . Поперечные срезы трёхлопастных разновозрастных побегов.
Обозначения см. на рис. 1
Таблица 1
Морфометрические показатели структурных элементов тканей побега Juniperus polycarpos C. Koch
|
Признак |
Двухлопастные побеги |
Трехлопастные побеги |
|||
|
x ± s x CV,% |
x ± s x CV,% |
||||
|
1-летний |
2-летний |
1-летний |
2-летний |
3-летний |
|
|
Кутикула, мкм |
10.5 ±0.82 |
12.5 ±0.64 |
12.3 ±0.78 |
12.0 ±0.82 |
13.5 ±1.00 |
|
24.6 |
16.3 |
20.3 |
21.5 |
23.4 |
|
|
Эпидерма, мкм |
13.3 ±1.12 |
12.5 ±0.91 |
14.5 ±0.62 |
13.8 ±1.13 |
13.3 ±1.06 |
|
26.8 |
23.1 |
13.6 |
26.1 |
25.2 |
|
|
Гиподерма, мкм |
15.3 ±0.69 |
22.8 ±1.80 |
14.5 ±1.22 |
20.0 ±1.05 |
25.3 ±1.60 |
|
14.4 |
25.1 |
26.7 |
16.7 |
20.1 |
|
|
Эндодермоидный слой, мкм |
33.5 ±3.42 |
22.8 ±2.59 |
31.3 ±2.14 |
39.0 ±3.78 |
32.8 ±2.09 |
|
32.3 |
36.1 |
21.7 |
30.7 |
20.2 |
|
Окончание табл. 1
|
Признак |
Двухлопастные побеги |
Трехлопастные побеги |
|||
|
x ± s x CV,% |
x ± s x CV,% |
||||
|
1-летний |
2-летний |
1-летний |
2-летний |
3-летний |
|
|
Флоэма, мкм |
39.3 ±2.11 17.0 |
65.5 ±2.66 12.8 |
41.3 ±2.39 18.3 |
69.0±4.63 21.2 |
152.0 ±4.21 8.8 |
|
Флоэма проводящая, мкм |
– |
16.3 ±1.30 25.4 |
– |
20.5 ±2.41 37.2 |
40.8 ±2.61 20.3 |
|
Ксилема, мкм |
25.5 ±1.57 19.5 |
71.0 ±2.27 10.1 |
34.8 ±1.99 18.1 |
90.8 ±3.92 13.7 |
283 ±10.48 11.7 |
|
Число трахеид в ксилемном ряду, шт. |
4.2 ±0.39 29.3 |
6.3 ±0.26 13.1 |
7.5 ±0.48 20.1 |
15.9 ±0.82 16.4 |
11.5 ±0.37 10.2 |
|
Число клеток в сердцевинном луче, шт. |
3.7 ±0.42 30.0 |
4.6 ±0.56 38.6 |
5.6 ±0.27 15.1 |
5.8 ±0.49 26.7 |
10.3 ±0.86 26.3 |
Уменьшение толщины эндодермоидного слоя у трехлетних побегов мы объясняем тем, что клетки становятся более плоскими в связи с увеличением диаметра ксилемы при незначительном увеличении общего диаметра стебля.
В листовых подушках хлорофиллоносная паренхима дифференцирована на палисадную и губчатую, граничащую с паренхимой первичной коры. В области губчатой ткани встречаются клетки двух типов: вытянутые с ровными контурами (40.5 и 28.9 мкм) и более крупные, с извилистыми клеточными стенками. Последние к третьему году разрушаются и объединяются в полости (рис. 3).
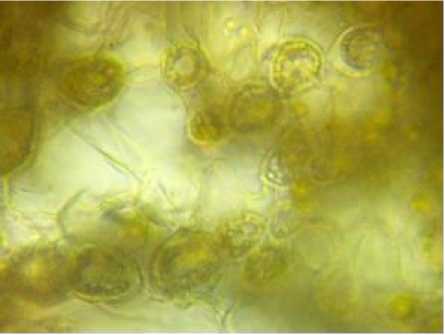
Рис. 3 . Смоляные полости
Кроме полостей, с обеих сторон листовых подушек ближе к ложбинкам побегов встречаются собранные в группы от двух (однолетние) до 14 (трехлетние) масляные клетки. На наличие в коре хвойных указанных групп клеток указывает и Л.И. Лотова [1987]. Для уточнения природы этих клеток и их принадлежности к тканям листа или стебля необходимо провести дополнительное анатомическое исследование и гистохимический анализ.
Коэффициент вариации (CV) у изученных признаков тканей стебля относительно стабильный и колеблется пределах от 8.8 у проводящей части флоэмы на третьем году жизни трехлопастного по- бега до 38.6% у числа клеток в сердцевинном луче первого года двухлопастных побегов. При этом высокая изменчивость показателей второго признака зависит от места прохождения поперечного среза и не всегда отражает реальную картину внутренней структуры поводящей системы.
У основной группы признаков изменчивость относительно невысокая, колеблется в пределах от 13 до 26%, что отражает надежность полученных результатов и, соответственно, возможность последующего их использования при сравнительной интерпретации признаков других видов и популяций. Показатели признаков как ксилемы, так и флоэмы с возрастом стабилизировались, и в связи с этим уменьшились значения CV от 17.0 до 12.8% у двулопастных и от 18.3 до 8.8% у трехлопастных по флоэме и от 19.5 до 10.1% и 18.1 до 11.7% по ксилеме соответственно. Изменчивость показателей числа трахеид в ксилемном ряду также с возрастом побега стабилизировалась. У остальных признаков выявить определенные закономерности в изменении показателей CV не удалось.
На поперечных срезах листовых подушек у побегов разных возрастов имеется по одному схизо-генному смоляному ходу, на некоторых срезах они не обнаруживаются, что может быть связано с местом прохождения среза. Форма смоляных ходов первоначально округлая или овальная радиально вытянутая (одно-, двулетние побеги), с возрастом при утолщении стебля их форма становится тан-гентально-вытянутой (трехлетние побеги), края их ровные. Размеры в зависимости от возраста и формы побега различны, у трехлопасных побегов крупнее (табл. 2). Смоляные ходы имеют многослойную (3–5 слоев клеток) обкладку из паренхимных клеток.
В паренхиме листовых подушек и первичной коре стебля встречаются склерефицированные (каменистые) клетки разных размеров (от 57.5 мкм до 107.5 мкм) и формы. Расположены они группами или по одному. Особенно много каменистых клеток в парен- химе двулопастных побегов независимо от возраста, трехлопастных побегов (рис. 4).
и они крупнее по сравнению с такими же клетками
Таблица 2
Морфометрические показатели признаков клеток тканей побегов Juniperus polycarpos C. Koch (поперечный срез)
|
Типы клеток и их признаки, в мкм |
Двухлопастные побеги x ± s x |
Трехлопастные побеги x ± s x |
||||
|
1-летний |
2-летний |
1-летний |
2-летний |
3-летний |
||
|
Эпидермальные |
д |
13.3 ±1.12 |
12.5 ±0.91 |
14.5 ±0.62 |
13.8 ±1.13 |
13.3 ±1.06 |
|
в |
9.0 ±0.41 |
10.0 ±0.53 |
12.3 ±1.08 |
10.8 ±0.75 |
9.5 ±0.50 |
|
|
Гиподермальные |
д |
15.3 ±0.82 |
14.5 ±0.97 |
14.5 ±1.22 |
20.0 ±1.05 |
14.8 ±0.58 |
|
ш |
13.0 ±0.82 |
12.8 ±0.95 |
12.0 ±0.89 |
15.8 ±0.75 |
13.0 ±0.73 |
|
|
Сердцевинные |
д |
14.5 ±1.22 |
13.5 ±0.85 |
16.3 ±1.07 |
17.8 ±1.02 |
18.8 ±1.50 |
|
ш |
8.5 ±0.93 |
10.0 ±0.75 |
13.3 ±0.99 |
12.0 ±0.73 |
12.8 ±0.69 |
|
|
Трахеиды |
д |
7.0 ±0.50 |
11.0 ±0.67 |
9.0 ±0.55 |
10.0 ±0.64 |
10.5 ±0.33 |
|
ш |
5.8 ±0.38 |
8.5 ±0.50 |
7.5 ±0.55 |
9.0 ±0.76 |
9.8 ±0.58 |
|
|
Флоэмные во локна |
д |
– |
14.5 ±0.75 |
– |
10.5 ±0.82 |
14.5 ±0.89 |
|
ш |
– |
5.3 ±0.45 |
– |
5.3 ±0.25 |
7.8 ±0.78 |
|
|
Сердцевинные лучи |
д |
13.5 ±1.06 |
16.5 ±1.07 |
13.8 ±1.0 |
17.8 ±1.02 |
22.5 ±1.75 |
|
ш |
8.0 ±0.50 |
6.5 ±0.41 |
7.0 ±0.50 |
6.8 ±0.38 |
7.5 ±0.37 |
|
|
Каменистые |
д |
единичн. |
71.1 ±7.39 |
– |
единичн. |
59.4 ±8.38 |
|
ш |
единичн. |
56.4±4.75 |
– |
единичн. |
44.4 ±4.71 |
|
|
Масляные |
д |
35.3 ±1.46 |
33.3 ±2.44 |
25.0 ±1.70 |
23,3 ±1,29 |
лизирован. |
|
ш |
25.0 ±1.05 |
26.3 ±1.67 |
18.8 ±1.0 |
17.8 ±0,95 |
лизирован. |
|
|
Смоляные ходы |
д |
267.5±34.03 |
305 ±0.0 |
335.8± 22.1 |
350 ±12.83 |
402.5 ±80.47 |
|
ш |
174.2 ±19.7 |
145 ±0.0 |
230 ±6.29 |
228,3±10.63 |
199.2 ±36.32 |
|
|
Обкладочные смоляных ходов |
д |
26.8 ±2.97 |
21.1 ±1.88 |
23,0 ±1.70 |
28,0 ±1.10 |
35.3 ±2.25 |
|
ш |
17.1 ±1.15 |
13.2 ±0.89 |
12,3 ±0.95 |
16,8 ±1.24 |
13.0 ±0.89 |
|
Примечание. Здесь и в табл. 3 – Д – длина, Ш – ширина, В – высота клеток. Радиальный диаметр трахеид, фло- эмных волокон и смоляных ходов обозначен так же буквами «Д – длинная ось», «Ш» - короткая ось.
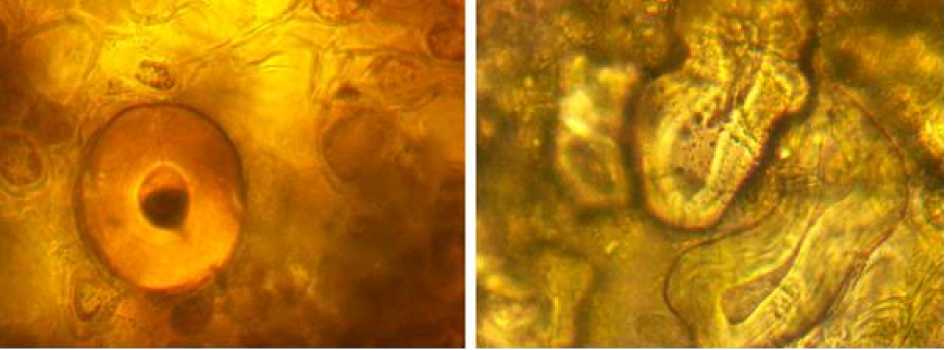
трехлопастной двухлопастной
Рис. 4. Каменистые клетки в паренхиме двухлетних побегов разной лопастности
Двух- и трехлопастные побеги первого и второго года жизни зеленого цвета, трехлетние ‒ бурого цвета и покрыты пробкой. Феллоген закладывается под листовыми подушками в эндодермоидном слое первичной коры. В середине лета феллема имеет серографитовый цвет и толщину 8–13 мкм, к осени цвет стебля приобретает характерную красно-бурую окраску.
Сердцевина стебля выраженная, у трехлопастных треугольной формы, в перимедуллярной части кото- рой расположены пучками тяжи первичной ксилемы. По мере роста в результате функционирования камбия у побегов в середине лета первого года формируется сплошное ксилемное кольцо. У однолетних двухлопастных побегов ксилема представлена двумя частями полулунной формы (по 4 пучка в каждой) (см. рис. 1), которые разделены листовыми лакунами, соединенными с сердцевиной стебля. У трехлопастных побегов ксилема разделена на три части, соответственно тремя листовыми лакунами (см. рис. 2).
Вторичная ксилема представлена трахеидами, расположенными радиальными рядами, число которых в зависимости от возраста побега (1–3-летние) изменяется у двухлопастных в среднем от 4.2 до 6.3 шт., у трехлопастных ‒ от 3.7 до 15.9 шт. Ксилемные ряды объединены в группы (максимальное число рядов в группах 11), их разделяют однорядные сердцевинные лучи. Размеры клеток сердцевинных лучей с возрастом увеличиваются от 13.5 до 16.5 мкм у двухлопастных побегов и от 13.8 до 22.5 мкм ‒ у трехлопастных. Во флоэме у трехлетних побегов J. polycar-pos , кроме проводящей системы и паренхимных клеток, встречаются и смолоносные полости. У трехлетнего трехлопастного побега толщина флоэмы достигает 152 мкм, что составляет пятую часть от толщины ксилемы.
Флоэма представляет собой два тангентальных слоя (проводящий и запасающий). Часть флоэмы с проводящей функцией на поперечных срезах трехлетних трехлопастных побегов имеет более светлую окраску (толщина 40.8 мкм) и состоит из четырехпяти рядов клеток. По бокам замыкают эту часть флоэмы крупные клетки тяжевой паренхимы. Размеры клеток тяжевой паренхимы увеличиваются в непроводящей части флоэмы и примыкают к фелло-дерме. Перидерма на третий год жизни побега уже формирует вокруг проводящей системы сплошное кольцо. Сердцевинные лучи во внешней части флоэмы приобретают более извилистую форму, видимо из-за внутреннего давления утолщающейся ксилемы и наружного сопротивления этому давлению уже сформированного сплошного слоя феллемы.
Большинство показателей размерных признаков клеток тканей побегов имеют незначительный (4.7– 15.1%) или средний (15.7–34.6%) коэффициент вариации (табл. 3). Какой-либо общей тенденции в изменении вариабельности изученных признаков у разновозрастных двух- и трехлопастных побегов выявить не удалось. CV одних признаков с возрастом увеличивается, других – уменьшается. Причем, если изменчивость признака «длина клеток эпидермы» у трехлопастных побегов с возрастом увеличивается в 1.9 раза, то признак «высота клеток эпидермы» наоборот, в 1.7 раз уменьшается. Коэффициенты вариации показателей клеток гиподермы у двухлопастных побегов имеют средний уровень вариации и с возрастом практически не изменяются (19.7–23.5%), в то же время у трехлопастных побегов наблюдается уменьшение вариабельности в 2.1 раза. Стабилизация CV показателей клеток гиподермы объясняется прекращением роста клеток и их растяжения вследствие раннего одревеснения (на первом году жизни). Наиболее вариабельными оказались размерные признаки смоляных ходов у трехлопастных побегов от 4.7% у одно- и двухлетних побегов до 34.6% ‒ у трехлетних. Низкий CV, возможно, связан с ранней закладкой и дифференциацией эпителиальных клеток будущих смоляных ходов однолетних побегов [Лотова, 1987], а к третьему году жизни побег утолщается в 1.4 раза за счет активного неравномерного прироста ксилемы, что приводит к неравномерному растягиванию смоляных ходов.
Таблица 3
Коэффициенты вариации показателей признаков клеток тканей побегов Juniperus polycarpos C. Koch (CV, %)
|
Типы клеток и их признаки, в мкм |
Двухлопастные побеги |
Трехлопастные побеги |
||||
|
1-летний |
2-летний |
1-летний |
2-летний |
3-летний |
||
|
Эпидермальные |
д |
26.8 |
23.1 |
13.6 |
26.1 |
25.2 |
|
в |
14.3 |
16.7 |
27.9 |
22.1 |
16.6 |
|
|
Гиподермальные |
д |
19.7 |
21.2 |
26.7 |
16.7 |
12.5 |
|
ш |
19.9 |
23.5 |
23.7 |
15.1 |
17.7 |
|
|
Сердцевинные |
д |
26.7 |
19.9 |
20.8 |
18.1 |
25.3 |
|
ш |
34.5 |
23.6 |
23.6 |
19.1 |
17.2 |
|
|
Трахеиды |
д |
22.6 |
19.2 |
19.4 |
20.4 |
10.0 |
|
ш |
21.0 |
19.7 |
22.2 |
26.8 |
18.9 |
|
|
Флоэмные волокон |
д |
– |
15.8 |
– |
24.6 |
19.6 |
|
ш |
– |
27.0 |
– |
15.1 |
32.1 |
|
|
Сердцевинные лучи |
д |
25.0 |
20.5 |
23.1 |
18.1 |
24.6 |
|
ш |
19.7 |
38.6 |
22.6 |
17.9 |
15.7 |
|
|
Каменистые |
д |
единич. |
31.2 |
– |
единич. |
28.2 |
|
ш |
единич. |
25.3 |
– |
единич. |
21.3 |
|
|
Масляные |
д |
13.1 |
23.3 |
21.6 |
17.6 |
лизиров. |
|
ш |
13.3 |
20.2 |
16.9 |
16.9 |
лизиров. |
|
|
Смоляные ходы |
д |
22.0 |
– |
11.4 |
6.3 |
34.6 |
|
ш |
19.6 |
– |
4.7 |
8.1 |
31.6 |
|
|
Обкладочные смоляных ходов |
д |
29.4 |
23.6 |
23.4 |
23.7 |
20.2 |
|
ш |
17.7 |
18.0 |
24.4 |
23.4 |
21.8 |
|
Заключение
В работе впервые дано описание анатомического строения молодых побегов Juniperus polycarpos , произрастающего в условиях Предгорного Дагестана. Цель – выявление при изучении анатомического строения стебля таксоноспецифичных и адаптивных признаков. На основе полученных результатов и сравнительного анализа литературных источников показано, что в целом побег J. polycar-pos имеет типичное для рода анатомическое строение [Лотова, 1987]. Специфическими особенностями его морфологии можно считать наличие двух типов побегов (двух- и трехлопастных). При этом у двухлопастных побегов, в отличие от трехлопастных, выявлено наличие в паренхиме листовых подушек первых большего числа каменистых клеток. Отличаются указанные побеги и строением центрального цилиндра. У трехлопастных побегов сердцевина имеет в целом треугольную форму с извилистым контуром. Он формируется благодаря наличию в перимедуллярной зоне множества (до десяти) тяжей первичной ксилемы. К концу первого года, с формированием камбия, ксилема формирует сплошное годичное кольцо. У двухлопастных побегов проводящие пучки к концу первого года полностью не сливаются и сохраняют автономность благодаря наличию более крупных листовых лакун.
Цвет стебля первого и второго года жизни зеленый, на третий год стебель полностью теряет зеленую окраску и приобретает бурый оттенок.
Межпопуляционные и межвидовые сравнительно-анатомические исследования вегетативных и генеративных органов Juniperus polycarpos C. Koch и J. excelsa Bieb. будут продолжены с целью выявления диагностических и адаптивных признаков этих видов и представлены в последующих сообщениях. При этом соответствующая обозначенным выше целям интерпретация количественных признаков может быть проведена только при получении сравнительных данных.
Список литературы Анатомическое строение побега Juniperus polycarpos С. Koch в предгорном Дагестане
- Ареалы деревьев и кустарников СССР. Л.: Наука, 1977. Т. 1. С. 36.
- Барыкина Р.П. Справочник по ботанической микротехнике. М.: Изд-во МГУ, 2004. 312 с.
- Галушко А.И. Флора Северного Кавказа. Ростов, 1978. Т. 1. 320 с.
- Исмаилов М.И. Можжевельники СССР: автореф. дис. ... д-ра биол. наук. Ташкент, 1975. 31 с.
- Кецховели Н.Н. Ксерофитные (аридные) редколесья // Растительность европейской части СССР. Л.: Наука, 1980. С. 273-276.
- Комаров В.Л. Род 42. Можжевельник, Верес - Juniperus L. // Флора СССР. Л.: Изд-во АН СССР, 1934. Т. 1. С. 174-191.
- Конспект флоры Кавказа. СПб.: Изд-во С.-Петерб. ун-та, 2003. 204 с.
- Красная книга Республики Дагестан. Махачкала, 2009. С. 53-250.
- Красная книга Российской Федерации (растения и грибы). М., 2008. 855с. Лотова Л.И. Анатомия коры хвойных. М.: Наука, 1987. 152 с.
- Муртазалиев Р.А. Конспект флоры Дагестана. Т. 1. (Lycopodiaceae - Urticaceae). Махачкала, 2009. 252 с.
- Самылина И.А., Аносова О.Г. Фармакогнозия. Атлас: учеб. пособие. М.: ГЭОТАР - Медиа, 2007. Т. 1. 192 с.
- Эзау К. Анатомия семенных растений. М.: Мир, 1980. Т. 1. 218 с.
- Adams R.P. Investigation of Juniperus species of the United States for new Sources of Cedarwood Oil. 2008. DOI: 10.1007/BF02859346.
- Adams R.P. et al. Evidence of relictual introgression or incomplete lineage sorting in nrDNA of Juniperus excelsa and J. polycarpos in Asia Minor // Phytologia. 2016. Vol. 98, № 2. P. 146-155.
- Farjon A. The taxonomy of multiseed junipers (Juniperus sect. Sabina) in Southwest Asia and East Africa (Taxonomic notes on Cupressaceae I) // Edinburgh Journal of Botany. 1992. Vol. 49. P. 251-283.
- Farjon A. Word checklist and bibliography of conifers. Kew: Royal Botanic Gardens, 2001. 309 p. Gadek P.A., Quinn C.J. Biflavones of the subfamily Cupressoideae, Cupressaceae // Phytochemistry. 1984. Vol. 24, № 2. P. 267-272.
- Hall M.T. Notes on cultivated junipers // ZZButler University Botanical Studies. 1961. № 14. P. 73-90.
- Hojjatti F. et al. Molecular phylogeny of Junirerus in Iran with special reference to the J. excelsa complex, focusing on J. seravschanica // Phytotaxa. 2018. Vol. 375, № 2. P. 135-157.
