Анатомическое строение вегетативных органов разных половых форм будры плющевидной (Glechoma hederacea L.)
")
Автор: Анисимова А.Г.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 2, 2004 года.
Бесплатный доступ
Изучено анатомическое строение вегетативных органов обоеполых и женских растений будры плющевидной с целью установления вторичных половых признаков. Оказались достоверными различия в анатомическом строении стебля и корневища, количестве эфирномасличных железок и головчатых волосков на нижней эпидерме листа, а также в числе устьиц и размере клеток
Короткий адрес: https://sciup.org/147204340
IDR: 147204340 | УДК: 581.8:
Anatomical structure of vegetative organs of different sexual forms of Glechoma hederacea L
The anatomical structure of vegetative organs of bisexual and female plants of Glechoma hederacea was studied with the рифозе of ascertaining of secondary sexual signs. The differences in anatomical structure of stem and rhizome, in the amount of attaral glands and capitated hairs on the lower epidermis, and also in the number of stomas and the size of cells turned out to be trustworthy.
Текст научной статьи Анатомическое строение вегетативных органов разных половых форм будры плющевидной (Glechoma hederacea L.)
Пермский государственный университет, 614990, Пермь, ул. Букирева, 15
Изучено анатомическое строение вегетативных органов обоеполых и женских растений будры плющевидной с целью установления вторичных половых признаков. Оказались достоверными различия в анатомическом строении стебля и корневища, количестве эфирномасличных железок и головчатых волосков на нижней эпидерме листа, а также в числе устьиц и размере клеток.
Будра плющевидная относится к семейству Губоцветные. Это многолетнее травянистое растение, имеющее стелющийся стебель 20-50 см длиной, с многочисленными приподнимающимися побегами. Листорасположение супротивное, листья почковидной формы. Цветки собраны по 3-5 в пазухах листьев, синевато-лиловые, длиной 10-18 мм. Растет на лугах, опушках, по сорным местам, у дорог (Флора СССР. 1954. Т. 20).
Будра плющевидная является ядовитым, лекарственным и медоносным растением (Овеснов, 1997). С лечебной целью используется надземная часть в период цветения как отхаркивающее средство, при почечно-каменной болезни и заболеваниях мочевого пузыря, печени и желчного пузыря, раке печени, подагре. Наружно водный настой применяется для ванн и компрессов при язвах, ранах, фурункулёзе. В траве содержатся эфирные масла, танин, свободные аминокислоты, холин, каротин, сапонины, смола, аскорбиновая кислота (Лавренов, Лавренова,1999).
Сведения об анатомическом строении вегетативных органов в литературе отсутствуют. Цель настоящего исследования - сравнительное изучение анатомических особенностей вегетативных органов разных половых форм будры плющевидной.
Как известно, будра плющевидная является ги-нодиэцичным растением, т.е. в ее популяциях встречаются крупноцветковые обоеполые и мелкоцветковые женские особи (Knuth, 1899; Демьянова, 1997). Для исследований мы собирали растения обеих половых форм, произрастающие в одной и той же ценопопуляции на территории Пермской области (Свердловский район, окрестности пос. Сылва) в фазу цветения. Собранный материал, в количестве 10 экземпляров каждой половой формы, изучали как в свежем, так и в фиксированном в 70°спирте состоянии. Поперечные и продольные срезы изготавливали лезвием безопасной бритвы. Временные препараты готовили по общепринятой анатомической методике с применением красителей: на крахмал - раствора Люголя, на одревеснение - сернокислого анилина (Прозина, 1960; Фурст, 1975). Срезы изучались под микроскопом МБИ-1. Измерения проводились с применением окуляр-микрометра. Рисунки выполнялись с помощью рисовального аппарата РА-4.
Полученные данные обработаны методом вариационной статистики (Лакин, 1980).
Стебель
Эпидерма состоит из удлиненных параллельно оси побега толстостенных клеток с неравномерно утолщенными стенками. Стенки клеток сверху покрыты тонким слоем складчатой кутикулы. Устьица овальной формы, диацитные, приподнятые над остальными клетками эпидермы (рис. 1).
Стебель в средней части на поперечном срезе четырехгранный, с выступающими в соответствии с положением четырех сосудисто-волокнистых пучков ребрами. Пучки открытого коллатерального типа. В гранях стебля могут присутствовать небольшие также коллатеральные пучки (рис. 2).
В выступающих частях стебля под эпидермисом располагается многослойная уголковая колленхима. Первичная кора представлена клетками ассимиляционной паренхимы, расположенными в 4-5 рядов. Обширная сердцевина занята крупными паренхимными клетками, в центре располагается полость.
На поверхности стебля имеются железистые волоски с 2-клеточной головкой, эфирномасличные железки, состоящие из 8 клеток, простые 1-, 2-, 3-, 4-, 5-, 6-, 7-клеточные волоски, не-
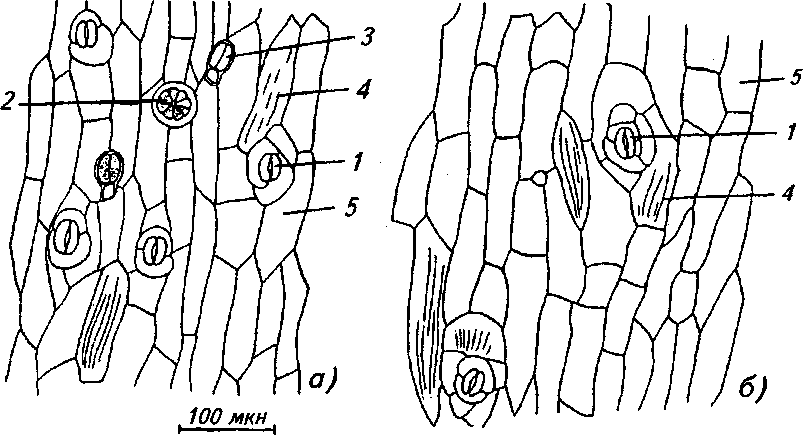
Рис. 1. Эпидерма стебля: а) обоеполая форма, б) женская форма:
1 - устьице, 2 - эфирно-масличная железка, 3 - железистый волосок, 4 - складчатая кутикула, 5 - клетки эпидермы
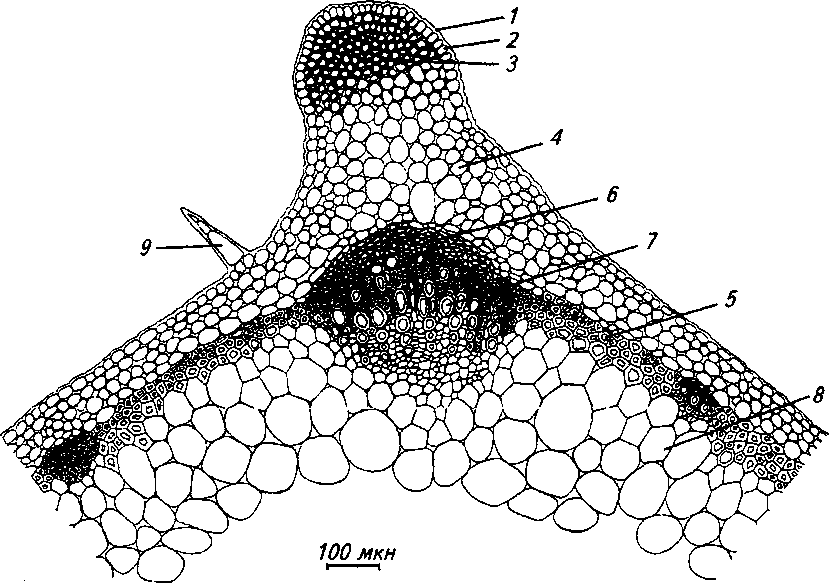
Рис. 2. Стебель в средней части (обоеполая форма):
1 - складчатая кутикула, 2 - эпидерма, 3 - уголковая колленхима, 4 - ассимиляционная паренхима, 5 - склеренхима, 6 - флоэма. 7 - ксилема, 8 - паренхима, 9 - простой 3-клеточный волосок
которые волоски со спавшимися клетками (рис. 3).
Результаты сравнения различных признаков в строении стебля женской и обоеполой половой формы будры плющевидной показали, что по количеству головчатых волосков, эфирно-масличных железок на 1мм2 поверхности женские и обоеполые особи не отличаются (табл. 1). Количество простых волосков преобладает у женских особей, причем строение последних более разнообразно.
т.е. кроме 1-, 2-, 3-, 4-, 5-клеточных волосков, характерных для обоеполых растений, присутствуют 6-, 7-клеточные с одной или двумя спавшимися клетками (рис. 3). Остальные признаки, вероятно, также могут являться диагностическими, т.к. разница между их значениями является достоверной (табл. 1), за исключением толщины флоэмы и кол-ленхимы, одинаковой у обеих половых форм.
Корневище клетками, сверху покрыта складчатой кутикулой.
п _ На поверхности очень редки простые волоски.
Покровная ткань эпидерма с толстостенными r r г
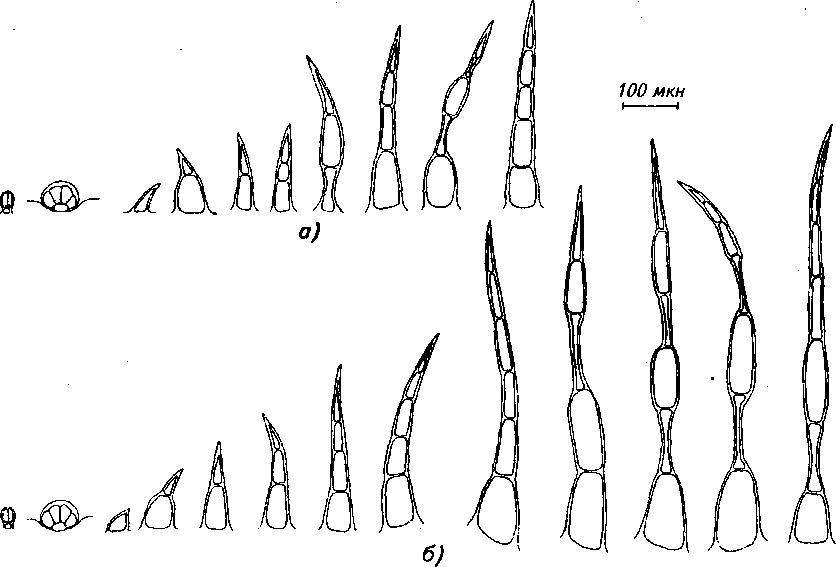
Рис. 3. Трихомы стебля: а) обоеполая форма, б) женская форма
Таблица 1
Анатомические признаки стебля обоеполой и женской формы Glechoma hederacea L.
|
Признак |
Женская форма |
Обоеполая форма |
td |
||
|
М±т |
Cv, % |
М±т |
Cv, % |
||
|
Диаметр стебля, мкн: шах |
2450143,51 |
7,94 |
3507135,48 |
4,52 |
18,83 |
|
min |
1678132,46 |
8,65 |
2592146,21 |
7,97 |
16,19 |
|
Толщина первичной коры, мкн |
98,513,35 |
15,19 |
12813,24 |
11,33 |
6,33 |
|
Число рядов клеток первичной коры |
4,3510,11 |
11,25 |
5,2010,14 |
11,85 |
4,77 |
|
Толщина флоэмы, мкн |
41,711,09 |
11,68 |
40,4011,12 |
12,40 |
0,26 |
|
Соотношение толщины флоэмы и радиуса стебля, % |
3,40 |
- |
3,29 |
- |
- - |
|
Толщина ксилемы, мкн |
190+5,58 |
13,12 |
27217,35 |
12,80 |
8,89 |
|
Соотношение толщины ксилемы и радиуса стебля, % |
15,51 |
- |
22,20 |
- |
- |
|
Число рядов сосудов в проводящем пучке |
7,30+0,21 |
12,65 |
9,75+0,18 |
8,07 |
8,86 |
|
Диаметр сердцевины, мкн |
1312+17,61 |
8,48 |
1972,60120,5 |
4,65 |
24,44 |
|
Соотношение радиуса сердцевины и радиуса стебля, % |
53,55 |
- |
80,51 |
- |
- |
|
Толщина колленхимы в ребре, мкн |
16914,75 |
12,57 |
157,8014,13 |
11,70 |
1,78 |
|
Толщина эпидермы, мкн |
20+0,41 |
9,20 |
24,7011,42 |
25,79 |
3,18 |
|
Наличие трихом и железок на эпидерме (шт. на 1 мм2 поверхности): а) головчатые волоски |
11,6011,17 |
10,4011,37 |
0,67 |
||
|
б) эфирно-масличные железки |
1,73+0,46 |
- |
1,8010,46 |
- |
0,12 |
|
в) простые волоски |
4,2710,88 |
- |
1,4010,44 |
- |
2,92 |
Примечание; Cv - коэффициент вариации, свидетельствующий о небольшом (0-10%), среднем (11-20%), большом (20%) варьировании признака; td - критерий достоверности разности, если он равен или больше 3, разница считается достоверной.
На поперечном срезе корневище слабо четы- ставлена запасающей паренхимой, расположенной рехгранное, почти округлое. Первичная кора пред- в 7'8 РЯД°В- в РебРах находятся открытые колла- теральные пучки, иногда пучковое строение пере- сающей паренхимы (рис. 4). ходит в непучковое. Сердцевина состоит из запа- .
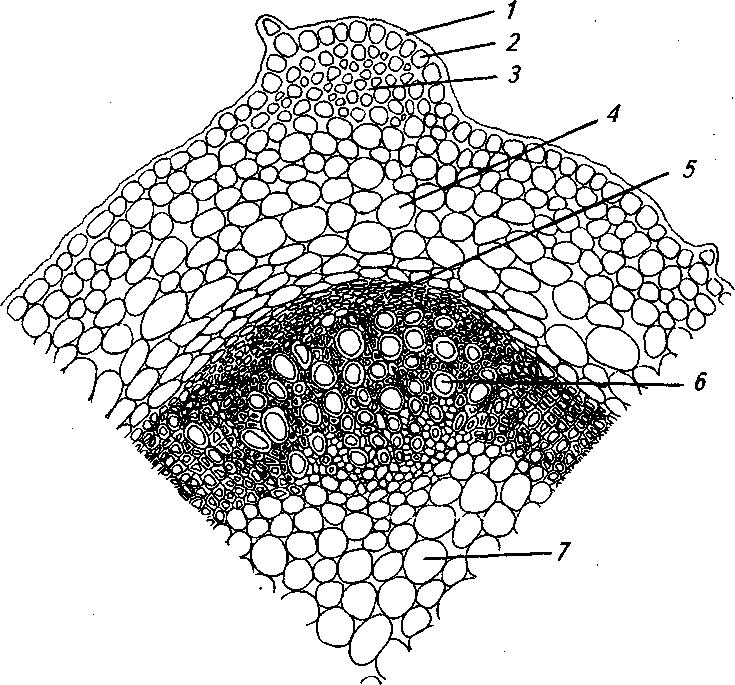
Рис. 4. Корневище в средней части (обоеполая форма):
1 - складчатая кутикула, 2 - эпидерма, 3 - уголковая колленхима, 4 - паренхима первичной коры, 5 - флоэма, 6 -ксилема, 7 - сердцевина
При сравнении полученных данных (табл. 2) можно сделать вывод, что диаметр корневища, толщина флоэмы и ксилемы больше у женских
особей (причем разница между их значениями является достоверной), что может рассматриваться в качестве диагностического признака.
Таблица 2
Анатомические признаки корневища обоеполой и женской формы Glechoma hederacea L.
|
Признак |
Женская форма |
Обоеполая форма |
td |
||
|
М±т |
Cv, % |
М±т |
Cv, % |
||
|
Диаметр корневища, мкн: max |
1697+32,73 |
8,62 |
1268,35+41,78 |
14,72 |
8,08 |
|
min |
1351+22,46 |
7,43 |
972,05+16,06 |
7,39 |
13,72 |
|
Толщина эпидермы, мкн |
20,40+0,34 |
7,55 |
23,05+1,23 |
23,95 |
2,08 |
|
Толщина первичной коры, мкн |
231+8,77 |
16,96 |
207,30+5,71 |
12,31 |
2,26 |
|
Соотношение толщины первичной коры и радиуса корневища, % |
27,22 |
_ |
32,69 |
- |
- |
|
Число рядов клеток первичной коры |
7,25+0,14 |
8,82 |
7,10+0,16 |
10,14 |
0,71 |
|
Толщина флоэмы, мкн |
33,50+1,78 |
23,70 |
17,50+1,32 |
33,83 |
7,22 |
|
Соотношение толщины флоэмы и радиуса корневища, % |
3,95 |
- |
2,76 |
- |
- |
|
Толщина ксилемы, мкн |
167,90+4,17 |
11,10 |
137,70+5,97 |
19,38 |
4,15 |
|
Соотношение толщины ксилемы и радиуса корневища, % |
19,79 |
- |
21,71 |
- |
- |
|
Максимальный диаметр сосудов, мкн |
31,70+1,28 |
। 18,11 |
36,05+1,46 |
18,06 |
2,24 |
Окончание табл. 2
|
Признак |
Женская форма |
Обоеполая форма |
td • |
||
|
М±т |
Cv, % |
М±т |
Cv, % |
||
|
Минимальный диаметр сосудов, мкн |
14,9010,72 |
21,54 |
11,0510,75 |
30,14 |
3,70 |
|
Диаметр сердцевины, мкн |
612,75139,26 |
28,64 |
319,15122,85 |
32 |
6,46 |
|
Толщина колленхимы в ребре, мкн |
102,5013,47 |
15,14 |
80,5513,14 |
17,43 |
4,69 |
Таблица 3
|
Анатомические признаки листа обоеполой и женской формы Glechoma hederacea L. |
|||||
|
Признак |
Женская |
форма |
Обоеполая форма |
td |
|
|
М±ш |
Cv, % |
М±ш |
Cv, % |
||
|
Толщина листовой пластинки, мкн |
144,4512,57 |
7,95 |
141,1512,63 |
8,33 |
0,89 |
|
Высота клеток верхней эпидермы, мкн |
26,4010,73 |
12,31 |
25,4010,64 |
11,30 |
1,07 |
|
Высота клеток нижней эпидермы, мкн |
18,20+0,64 |
15,71 |
18,8010,79 |
18,88 |
0,59 |
|
Толщина мезофилла, мкн |
97,9012,64 |
12,03 |
90,9512,20 |
10,83 |
2,02 |
|
Толщина палисадного мезофилла, мкн |
35,60+2,21 |
27,75 |
35,2010,88 |
11,11 |
0,17 |
|
Отношение палисадного мезофилла к толщине всего мезофилла, % |
36,36 |
- |
38,70 |
- |
- |
|
Толщина губчатого мезофилла, мкн |
62,4512,95 |
21,15 |
55,8511,92 |
15,34 |
1,88 |
|
Отношение губчатого мезофилла к толщине всего мезофилла, % |
63,79 |
- |
61,41 |
- |
- |
|
Верхняя эпидерма |
|||||
|
Длина клеток эпидермы, мкн |
85,1012,86 |
15,01 |
63,5014,14 |
29,15 |
4,29 |
|
Ширина клеток эпидермы, мкн |
42,1012,13 |
22,59 |
35,7511,21 |
15,19 |
2,59 |
Лист
Эпидермис однорядный, с тонкой складчатой кутикулой. Клетки верхней эпидермы более крупные, толщина кутикулы больше, чем у клеток
нижней эпидермы, стенки их слабо извилистые, практически прямые (рис. 5). Количество клеток на 1 мм2 поверхности эпидермы больше у обоеполых особей (табл. 3).
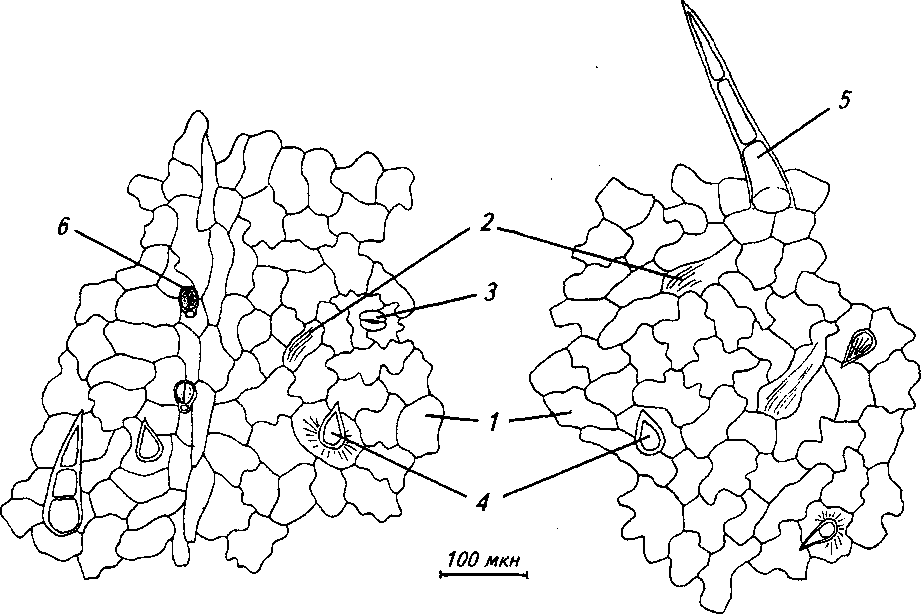
Рис. 5. Верхняя эпидерма листа: а) обоеполая форма, б) женская форма:
I - клетки эпидермы, 2 - складчатая кутикула, 3 - устьице, 4 - простой одноклеточный волосок, 5 - простой 3-клеточный волосок, 6 - железистый волосок
Окончание табл. 3
|
Признак Количество устьиц на 1 мм2 |
Женская форма |
Обоеполая форма |
td |
||
|
М±т |
Су, % |
М±т 3,75±2,05 |
Су, % 244 |
||
|
Длина устьица, мкн |
- |
- |
32,67+0,33 |
1,78 |
— |
|
Ширина устьица, мкн |
- |
- |
24,33±0,67 |
4,73 |
— |
|
Количество эфирно-масличных железок на 1 мм2 |
0 |
- |
0 |
- |
- |
|
Количество железистых волосков на 1 мм2:
|
5,00±1,12 2,40+0,89 0 |
- |
7,20±1,56 2,40±0,65 1,20+0,61 |
1,15 0 1,97 |
|
|
Количество простых волосков на 1 мм2: а) вдоль главной жилки 61 на листовой пластинке в) по краю листа |
6,50±0,94 6,40+1,07 3,00+0,00 |
- |
9,20±1,47 4,80±0,80 7,60±1,26 |
- |
1,55 1,20 3,65 |
|
Количество клеток на 1 мм2 |
367,50±9,27 |
11,28 |
. 545+16,87 |
13,85 |
9,22 |
Нижняя эпидерма
Клетки нижней эпидермы несколько меньше, разных половых форм, с преобладанием последних стенки их более извилистые (рис. 6). Количество у женских особей (табл. ЗУ *
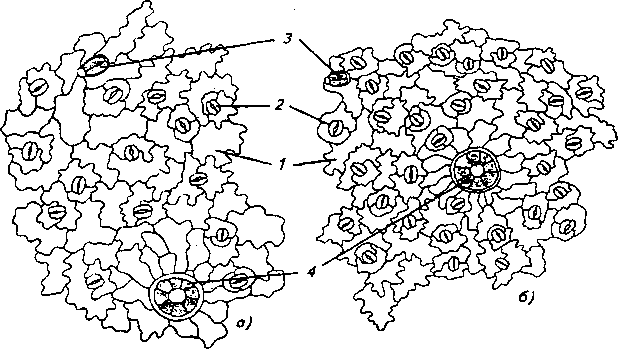
Рис. 6. Нижняя эпидерма листа: а) обоеполая форма, б) женская форма: 1 - клетки эпидермы, 2 - устьице, 3 - железистый волосок, 4 - эфирно-масличная железка
Листья у женских особей гипостоматические, т.е. устьица находятся на нижней стороне листа. У обоеполых особей небольшое количество устьиц было отмечено и на верхней эпидерме, т.е. у них листья амфистоматические. У обеих половых форм устьица овальной формы, диацитные.
Количество устьиц на 1мм2 поверхности нижней эпидермы также сильно отличается у разных половых форм: у женских форм их значительно больше, что может свидетельствовать о предрасположенности женских форм к более высокой интенсивности транспирации в сравнении с обоеполыми.
Литературные данные свидетельствуют о том, что у двудомных растений мужские и женские экземпляры различаются по уровню водного обмена, с преобладанием последнего у женских форм (Шереметьев, 1983). Можно высказать предположение, что подобная закономерность свойственна не только двудомным растениям, но и гинодиэцич-ным. Ранее к подобному выводу пришла Е.И. Демьянова при изучении половой структуры популяций у сексуально-полиморфных растений (Демьянова, 1990). Автором обнаружена большая требовательность к условиям (прежде всего, лучшего почвенного увлажнения) женских форм двудомных, гинодиэцичных и функционально двудомных растений.
На верхней эпидерме листа у обеих половых форм встречаются головчатые волоски с 1- и 2клеточной головкой, они располагаются в основном вдоль главной и отчасти боковых жилок. Эфирно-масличные железки отсутствуют как у женских, так и у обоеполых особей. Простые во-
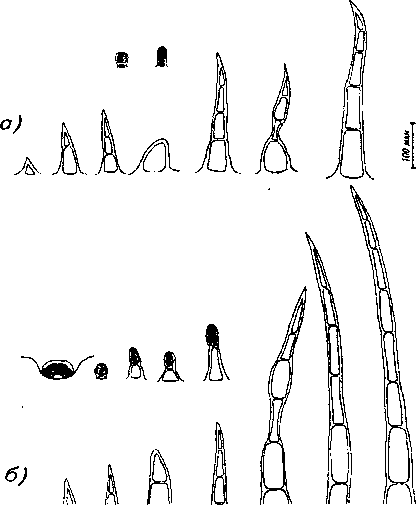
Рис. 7. Трихомы листа обоеполых растений: а) верхняя эпидерма, б) нижняя эпидерма лоски 1-, 2-, 3-, 4-, 5-, 6-клеточные в основном находятся вдоль жилок и по краю листа. Строение простых волосков разнообразнее у женских особей (рис. 7, 8). Их количество также несколько больше у обоеполых форм (табл. 3).
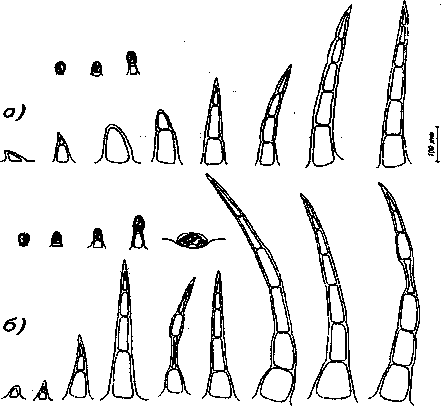
Рис. 8. Трихомы листа женских растений: а) верхняя эпидерма, б) нижняя эпидерма
На нижней эпидерме листа имеются железистые волоски с 1- и 2-клеточной головкой, на короткой или длинной ножке (рис. 7, 8). Располагаются они вдоль жилок и количество их на 1 мм2 больше у обоеполых особей (табл. 3). Эфирномасличные железки находятся на листовой пластинке между жилками, причем их количество, приходящееся на 1 мм2, больше у женских форм. Простые волоски более разнообразны по строению у женских особей, располагаются в основном вдоль жилок. Их количество на 1 мм2 поверхности несколько больше также у женских особей.
На поперечном срезе листа обнаруживается дорсовентральное строение. Палисадная ткань однорядная, клетки её прерываются в области жилок. Губчатая ткань рыхлая, с довольно крупными межклетниками, представлена 3-4 рядами клеток (рис. 9). Толщина мезофилла в целом несколько больше у женских особей (табл. 3). Главная жилка сильно выпирает на нижней стороне листа, имеет один коллатеральный пучок (рис. 10). Механическая ткань главной жилки представлена уголковой колленхимой, образованной 4-5 рядами клеток, с боков переходящих в клетки хлоренхимы. Черешок листа с верхней стороны несколько вогнутый. Сосудисто-волокнистый пучок лежит в центре черешка. Кроме центрального пучка имеется по одному небольшому пучку в выступах черешка. Клетки паренхимы крупные, почти без межклетников. В выступах черешка и в его нижней части имеется уголковая колленхима. Эпидермис черешка представлен одним рядом клеток, окружен тонкой кутикулой, покрыт железистыми волосками, эфирно-масличными железками и простыми волосками.
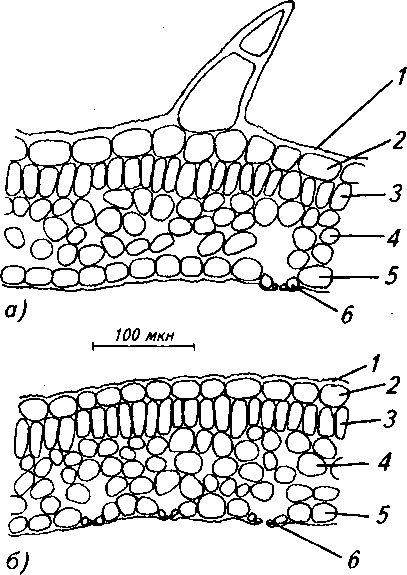
Рис. 9. Анатомическое строение листовой пластинки: а) обоеполого растения, б) женского растения:
1 - складчатая кутикула, 2 - верхняя эпидерма, 3 - палисадная паренхима, 4 - губчатая паренхима, 5 - нижняя эпидерма, 6 - устьице
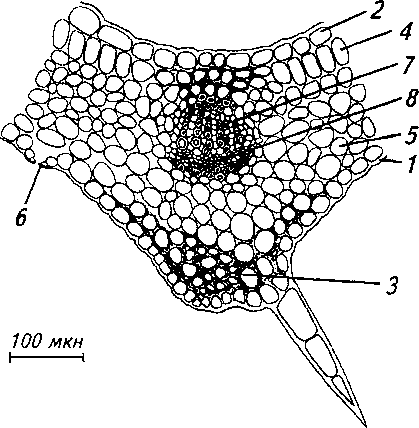
Рис. 10. Анатомическое строение главной жилки листа обоеполого растения:
1 - складчатая кутикула, 2 - верхняя эпидерма, 3 -уголковая колленхима, 4 - палисадная паренхима, 5 - губчатая паренхима, 6 - устьице, 7 - ксилема, 8 - флоэма
По анатомическому строению листовой пластинки, жилки листа и черешка обоеполые и женские особи не отличаются.
Выводы
-
1. При сравнении строения стебля у обоеполых и женских особей диагностическими могут являться все исследуемые признаки, кроме толщины флоэмы и колленхимы. В строении корневища у обоеполых и женских особей достоверными оказались: диаметр корневища, толщина флоэмы и ксилемы, диаметр сердцевины, толщина колленхимы.
-
2. Верхняя эпидерма листа характеризуется большим количеством клеток и простых волосков у обоеполых растений. Клетки нижней эпидермы имеют более крупные размеры у обоеполых растений по сравнению с женскими.
-
3. Число эфирно-масличных железок и головчатых волосков на эпидерме стебля у обоеполых и женских особей практически одинаково. На нижней эпидерме листа у обоеполых растений преобладают головчатые волоски, у женских растений -эфирно-масличные железки.
-
4. Количество устьиц и клеток на нижней эпидерме больше у женских особей, что, вероятно, свидетельствует о большей их предрасположенности к более высокой интенсивности транспирации в сравнении с обоеполыми растениями.
3. Биология. С. 9-20.
Список литературы Анатомическое строение вегетативных органов разных половых форм будры плющевидной (Glechoma hederacea L.)
- Демьянова Е.И. Половой полиморфизм цветковых растений: Автореф. дис... д-ра биол. наук. М., 1990.
- Демьянова Е.И. Половая структура природных популяций сексуально-полиморфных растений Предуралья//Вестник Перм. ун-та. 1997. Вып. 3. Биология. С. 9-20.
- Лавренов В.К., Лавренова Г.В. Полная энциклопедия лекарственных растений. СПб.: Нева; М.: ОЛМА-ПРЕСС, 1999. Том I.
- Лакин В.Ф. Биометрия. М.: Высш. шк., 1990.
- Овеснов С.А. Конспект флоры Пермской области. Пермь: Изд-во Перм. ун-та, 1997.
- Прозина М.Н. Ботаническая микротехника. М., I960.
- Фурст Г.Г. Методы анатомо-гистохимического исследования растительных тканей. М.: Наука, 1975.
- Шевченко СВ., Денисова Г.А. Данные по морфологии и анатомии шалфея мускатного//Растительные ресурсы. 1970. Т. 6, вып. 4, С. 610-615.
- Шереметьев С.Н. О приспособительном значении полового диморфизма цветковых растений//Бот. журн. 1983. Т. 68, № 5. С. 561-570.
- Шишкин Б.К. Labiatae//Флора СССР. М.;Л.: Изд-во АН СССР, 1954. Т. 20.
- Knuth P. Handbuch der Blutenbiologie. Leipzig: Verlag von Wilhelm Engelmann, 1899. Bd II, T. II. 705 S.
