Анатомо-морфологические особенности листа Rosa rugosa Thub. в разных климатических условиях

Автор: Сорокопудов В.Н., Кузнецова Т.А., Шлапакова С.Н., Филипповская А.О., Лукашов Е.С.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Биологические науки
Статья в выпуске: 5, 2018 года.
Бесплатный доступ
Цель исследования - оценка способности адап-тироваться R. rugosa к разным климатическим условиям; выявление наиболее информативных ксероморфных признаков, которые можно исполь-зовать в селекционной работе. Задачи исследова-ния - провести морфо-анатомическое исследова-ние адаксиального и абаксиального эпидермиса листа, исследовать водный режим и определить степень засухоустойчивости растений и способ-ность их к адаптации. Исследования проводились в Белгороде в 2009 г., в Брянске и Санкт-Петербурге - в 2014 г. в фенофазу «формирование плодов». Материал отбирался по общепринятой методике с учетом возраста растений и морфо-логического адреса, изучение эпидермиса и его структур проводилось с помощью СМ Levenhuk 320, видеокамеры Levenhuk C310 NG, окуляр-микрометра MOB-1-15×У4,2, РЭМ Quanta 200 3D, вычисляли коэффициент извилистости антикли-нальных стенок. Для исследования водного режима растений использовалась методика Никитского ботанического сада. Определена оводненность, водный дефицит и водоудерживающая способ-ность листа...
Лист, водный режим, адаптация, ксероморфные признаки, основ-ные клетки эпидермиса, устьица, трихомы
Короткий адрес: https://sciup.org/140238115
IDR: 140238115 | УДК: 635.9
Anatomical and morphological features of a leaf Rosa rugosa Thunb. in different climatic conditions
The aim of the study was to assess the ability to adapt R. rugosa to different climatic conditions and to identify the most informative xeromorphic features that can be used in the selective work. The objectives of the study were to conduct morpho-anatomical analysis of adaxial and abaxial epidermis of the leaf, to study water regime and to determine the degree of drought re-sistance of plants and their ability to be adapted. The studies were carried out in phenological stage of “fruit formation” in Belgorod in 2009, and in Bryansk and Saint-Petersburg in 2014. The material was selected according to conventional method taking into account the age of plants and morphological address, the study of the epidermis and its structures was carried out using CM Levenhuk 320, video camera Levenhuk C310 NG, eyepiece micrometer MOB-1-15×U4,2, SEM Quanta 200 3D, calculating hereby the coefficient of tortuosity of anticlinal walls. The technique of Nikitsky Botanical gar-den was used to study water regime of plants...
Текст научной статьи Анатомо-морфологические особенности листа Rosa rugosa Thub. в разных климатических условиях
Введение . Высокая устойчивость растений к различным факторам среды – залог хорошего урожая. Исследование путей адаптации растений к разным климатическим условиям позволяет оценить их потенциал приспособления, зону оптимума воздействия факторов. Выявленные признаки устойчивости можно использовать в селекционной работе для отбора наиболее перспективных растений, устойчивых к неблагоприятным факторам среды.
Rosa rugosa является интродуцированным видом в Белгородской области и Санкт-Петербурге. Ареал -Дальний Восток России (Камчатка, Приморье, Сахалин), Китай, Япония и Корея. Плоды шиповника обладают фитонцидным и мощным бактерицидным свойством, содержат большое количество антиоксидантов [6].
Цель исследования : оценка способности адаптироваться R. rugosa к разным климатическим условиям; выявление наиболее информативных ксеро-морфных признаков, которые можно использовать в селекционной работе.
Задачи исследования : провести морфоанатомическое исследование адаксиального и абаксиального эпидермиса листа, исследовать водный режим и определить степень засухоустойчивости растений и способность их к адаптации.
В 2009 г. в Белгороде наблюдались резкие колебания температуры, засуха; сумма положительных среднесуточных температур за вегетационный период – 3 460 °С, сумма осадков – 249 мм. В Санкт-Петербурге в 2014 г. сумма положительных среднесуточных температур за вегетационный период – 2 440 °С, сумма осадков – 388 мм. В Брянске в 2014 г. сумма положительных среднесуточных температур за вегетационный период – 2 648 °С, сумма осадков – 268 мм.
Исследование проводилось в фенофазу «формирование плодов», учитывались также погодные условия. Материал отбирался по общепринятой методике с учетом возраста растений и морфологического адреса [6]. В Белгороде исследование проводили в 2009 г., в Санкт-Петербурге и Брянске – в
2014 г. Изучение эпидермиса и его структур проводилось с помощью СМ Levenhuk 320, видеокамеры Levenhuk C310 NG, окуляр-микрометра MOB-1-15×У4,2, РЭМ Quanta 200 3D. Подсчет числа устьиц, вычисление степени открытости устьиц проводилось по общепринятым методикам, вычисляли коэффициент извилистости антиклинальных стенок [5]. Для исследования водного режима растений использовалась методика Никитского ботанического сада [2].
Общее содержание воды в листовой пластинке (учитывался водный дефицит), сформированной в условиях Белгорода и Брянска – низкое, в условиях Санкт-Петербурга – высокое (табл. 1). Водоудерживающая способность растений, определенная в ходе шестичасового увядания, в условиях Санкт-Петербурга средняя и достоверно выше, чем в условиях Белгорода и Брянска, где она низкая.
Таблица 1
Характеристика водного режима листьев R. rugosa в разных климатических условиях
|
Место произрастания |
Общая вода, % |
Часы увядания |
М, % |
W, % |
D w , % |
|||
|
2 |
4 |
6 |
8 |
|||||
|
Санкт-Петербург |
71,00± 0,576 ** |
18,36± 0,666 |
26,79± 0,897 |
34,59± 0,999 |
43,41± 1,790 |
2,30± 0,559 |
95,00± 2,51 ** |
0 |
|
Белгород |
57,05± 2,33 * |
29,51± 5,35 |
47,17± 8,35 |
56,61± 8,06 |
- |
36,27± 11,84 |
79,65± 8,99 * |
5,16± 2,04 |
|
Брянск |
58,05± 1,35 ** |
29,87± 4,39 |
48,26± 5,85 |
57,55± 5,32 |
- |
35,68± 9,98 |
78,92± 7,98 * |
5,08± 3,25 |
Примечание : M – процент повреждения; W – процент восстановления; D w – водный дефицит; здесь и далее : *достоверные отличия при уровне вероятности 0,95; **при уровне вероятности 0,99 (контроль – показатели 2015 г.);
После шестичасового увядания в условиях Белгорода и Брянска наблюдается мацерация тканей листовой пластинки (до 36 % от всей площади), в условиях Санкт-Петербурга даже после восьмичасового увядания мацерация тканей не превышает 2– 3 % площади листовой пластинки. Эти данные согласуются с низким процентом восстановления оводненности тканей в условиях Белгорода и Брянска, в Санкт-Петербурге этот показатель в среднем составляет 95 %.
По данным исследования водного режима, у 12 видов рода Rosa в условиях Белгорода и 6 видов в Брянске в фенофазу «формирования плодов» R. rugosa отнесена к группе растений со средней степенью засухоустойчивости. Одним из критериев было низкое содержание связанной воды в тканях листа [4].
Анализ данных настоящего исследования позволяет сделать вывод, что к климатическим условиям Санкт-Петербурга R. rugosa более адаптирована, чем к засушливым условиям Белгорода и Брянска. Влагоудерживающая способность листьев, сформированных в условиях засухи, недостаточна, что ве- дет к необратимым изменениям в тканях листа после шестичасового увядания.
Физиологические функции растительного организма тесно связаны с его морфологическими и анатомическими признаками.
Корреляционный анализ позволил предположить, что основная роль в потере влаги при транспирации принадлежит устьицам. Коэффициент корреляции между степенью открытости устьиц (СОУ) и потерей влаги при шестичасовом увядании достоверный средней силы (r = 0,577), между числом устьиц на единицу поверхности и потерей влаги при шестичасовом увядании также достоверный средней силы (r = 0,591). Ранее была найдена сильная обратная зависимость между размерами устьиц и их числом (r = 0,874) [5].
С увеличением ксероморфности строения листовой пластинки отмечается уменьшение размеров устьиц [1]. Однако в нашем исследовании в засушливых условиях Белгорода формируются листья с более крупными устьицами (табл. 2).
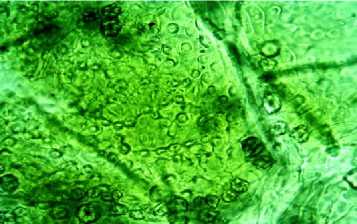
Устьица погруженные . Замыкающие клетки по периферии прикрыты основными клетками эпидермиса (рис. 1, А).
Характеристика устьичного аппарата R. rugosa в разных климатических условиях
Таблица 2
|
Место произрастания |
СОУ, % |
К у |
S, мкм2 |
N, шт/мм2 |
|
Санкт-Петербург |
7,68±0,314** |
1,06±0,0184** |
303,84±5,96 ** |
164,30±5,23 ** |
|
Брянск |
20,41±0,359 * |
1,12±0,011 * |
311,11±6,16 * |
144,52±6,11 * |
|
Белгород |
21,61±0,529 * |
1,14±0,021 * |
315,06±9,13 * |
133,66±8,74 * |
Примечание: СОУ – степень открытости устьиц, %; К у – коэффициент удлиненности устьиц; S – площадь устьица, мкм2; N – число устьиц, шт/мм2.
А
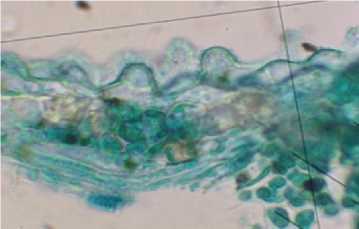
Рис. 1. Абаксиальный эпидермис листовой пластинки R. rugosa (Санкт-Петербург): А - поверхность эпидермиса; Б - срез эпидермиса
Б
В условиях Белгорода и Брянска концентрация устьиц на единицу поверхности абаксиальной поверхности листа существенно меньше, чем у листьев, сформированных в условиях Санкт-Петербурга. По литературным данным с повышением ксеро-морфности листьев число устьиц на единицу поверхности снижается [2].
Степень открытости устьиц в условиях Белгорода достоверно выше, чем у листьев, сформированных в условиях избыточного увлажнения Санкт-Петербурга, что объясняет высокую потерю влаги при шестичасовом увядании (табл. 3).
Таблица 3
|
Место произрастания |
Адаксиальный эпидермис |
Абаксиальный эпидермис |
||||
|
S ок ,мкм2 |
К и |
Т, мкм |
S ок , мкм2 |
К и |
Т, мкм |
|
|
Санкт-Петербург |
526,84± 20,51 * |
3,28± 0,749* |
18,90± 0,571 * |
266,05± 9,84* |
3,89± 0,159* |
13,60± 1,09 * |
|
Белгород |
1290,34± 28,54 * |
12,95± 0,325 * |
16,58± 0,956* |
653,12± 14,86 * |
13,99± 0,352 * |
14,38± 1,29 * |
|
Брянск |
1212,56± 25,32 * |
11,75± 0,227 * |
17,09± 0,611 * |
611,34± 12,44 * |
12,99± 0,321 * |
14,22± 1,11 * |
Примечание : S ок - площадь основных клеток эпидермиса, мкм2; К и - коэффициент извилистости антиклинальных стенок основных клеток эпидермиса, мкм; Т - толщина эпидермиса, мкм.
Морфо-анатомическая характеристика основных клеток эпидермиса
R. rugosa в разных климатических условиях
Очертания основных клеток эпидермы листа варьируют от прямолинейных до волнистых и извилистых. Степень волнистости стенок основных эпидермальных клеток зависит от условий произрастания растений [5]. В условиях Белгорода у основных клеток адаксиального эпидермиса коэффициент извилистости антиклинальных клеток существенно выше, чем у листьев, сформированных в условиях Санкт-Петербурга.
Размеры и форма основных клеток связаны между собой компенсаторными корреляциями, обеспечивающими согласованное изменение пло- щадей верхней и нижней эпидермы листовой пластинки, которое является важным условием поддержания ее плоской формы [3]. Основные клетки абаксиального эпидермиса имеют выросты, которые в условиях Санкт-Петербурга достигают 13,30±0,774 мкм (рис. 2, А).
Площадь основных клеток абаксиального и адаксиального эпидермиса достоверно ниже в условиях Санкт-Петербурга. Однако большие выросты основ- ных клеток абаксиального эпидермиса создают дополнительную поверхность для увеличения транспирации.
Лист R. rugosa в условиях Белгорода имеет на абаксиальной и адаксиальной поверхности три типа трихом: кроющие одноклеточные трихомы, железистые головчатые трихомы на ножке, пельтатные трихомы (единичные) [5].
А
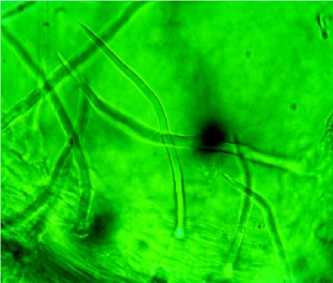
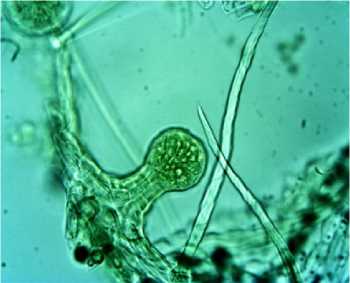
Рис. 2. Классификация трихом R. rugosa на поверхности листовой пластинки (Санкт-Петербург): А – простые кроющие трихомы; Б – головчатые трихомы
Б
В условиях Санкт-Петербурга трихомы обнаружены только на абаксиальной поверхности, пель-татные железки не обнаружены.
В условиях Белгорода листовые пластинки R. rugosa более опушенные, число одноклеточных кроющих трихом в условиях Белгорода – 1378– 1550 шт. на 1 мм2, их длина – 58,60±4,44 мкм (рис. 2). Основные клетки эпидермиса, прилегающие к основанию одноклеточного волоска, имеют трапециевидную форму.
Таблица 4
|
Место произрастания |
Длина ножки, мкм |
Ширина ножки, мкм |
Диаметр головки, мкм |
N гт , шт/мм2 |
|
Санкт-Петербург |
5,49±0,376 ** |
4,12±0,176 |
5,89±0,198 (8–10 клеток) |
8,397±0,924** |
|
Белгород |
19,61±2,69 * |
– |
(9–10 клеток) |
199,80±21,67 * |
|
Брянск |
18,23±2,14 * |
– |
– |
178,46±12,31 * |
Примечание: N гт - число головчатых трихом на 1 мм2 площади листовой пластинки, шт/мм2.
Характеристика головчатых трихом R. rugosa в разных климатических условиях
В условиях Санкт-Петербурга на абаксиальной поверхности обнаружены головчатые и простые кроющие трихомы, концентрирующиеся на жилках. Головчатые трихомы имеют в головке 8–10 клеток, в ножке – 2 клетки в ширину и 2 клетки в длину. В условиях Белгорода головчатые трихомы достоверно более крупные, их число на единицу поверхности больше, чем в Санкт-Петербурге.
Выводы
R. rugosa в большей степени адаптирована к условиям Санкт-Петербурга, чем Белгорода и Брян- ска. Влагоудерживающая способность листьев, сформированных в условиях засухи, недостаточна, что ведет к необратимым изменениям в тканях листа после шестичасового увядания. Увеличение СОУ в условиях Белгорода (в Брянске он немного меньше) способствует повышению транспирации и уменьшению влагоудерживающей способности.
В условиях Белгорода наблюдается проявление ксероморфности в строении листовой пластинки: уменьшение числа устьиц на единицу поверхности, увеличение извилистости антиклинальных стенок основных клеток эпидермиса, увеличение опушен-ности листовой пластинки, более крупные трихомы.
Список литературы Анатомо-морфологические особенности листа Rosa rugosa Thub. в разных климатических условиях
- Васильев Б.Р. Строение листа древесных растений различных климатических зон/под ред. В.М. Шмидта. -Л.: Изд-во ЛГУ, 1988. -208 с.
- Кушниренко М.Д. Физиология водообмена и засухоустойчивости растений. -Кишинев: Штиинца, 1991. -307 с.
- Паутов А.А., Васильева В.А. Роль формы основных клеток эпидермы в морфогенезе листа представителей Hamamelidaceae//Ботанический журнал. -2010. -Т. 95. -№ 3. -С. 338-345.
- Резанова Т.А., Сорокопудов В.Н., Свинарев Е.Н. и др. Адаптация видов рода Rosa L. в условиях Белгородской области//Фундаментальные исследования. -2012. -№ 11-2. -С. 309-312.
- Резанова Т.А. Сорокопудов В.Н., Свинарев Е.Н. Особенности анатомического строения листа у видов рода Rósa L. (Rosaceae Juss.)//Научные ведомости Белгородского государственного университета. Сер. «Естественные науки». -2011. -№ 9 (104), Вып. 15/1. -С. 337-347.
- Сорокопудов В.Н., Нетребенко Н.Н., Евтухова М.В. и др. Морфо-анатомические аспекты некоторых представителей рода Rosa L.//Вестн. КрасГАУ. -2009. -№ 11. -С. 50-54.
