Анатомо-морфологические закономерности проводящего стволового и фотосинтетического листового аппарата древесных растений в контрастных условиях произрастания

Автор: Смирнова Н.И., Павлов И.Н.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Агролесомелиорация и лесное хозяйство
Статья в выпуске: 2, 2014 года.
Бесплатный доступ
В результате исследования выявлены высокая чувствительность устьичного аппарата и ксилемы древесных растений на условия произрастания (тип условий местопроизрастания, влажность почвы, положение в пологе и пр.) и климатические изменения (температура, осадки).
Устьица, ксилема, водный режим
Короткий адрес: https://sciup.org/14083513
IDR: 14083513 | УДК: 581.4
Anatomic-morphological regularities of the conductive stem and photosynthetic leaf apparatus of arboreal plants in the contrast growth conditions
The significant sensitiveness of the arboreal plant stomatal apparatus and xylem to the growth conditions (type of growth place, soil humidity, location in forest canopy, etc.) and climate changes (temperature, precipitation) is revealed as a result of the research.
Текст научной статьи Анатомо-морфологические закономерности проводящего стволового и фотосинтетического листового аппарата древесных растений в контрастных условиях произрастания
Многочисленными исследованиями установлено влияние условий произрастания на формирование устьичного аппарата [8, 13, 15, 20, 21] и ксилемы [1, 24]. В то же время совсем мало известно о координации двух меристем (верхушечной и боковой) в регуляции процессов роста и развития основной водопроводящей системы древесных растений – ксилемы.
Ксилема – основная водопроводящая ткань сосудистых растений. Поэтому её строение и развитие следует рассматривать в функциональной связи со всеми элементами, и в первую очередь листового аппарата. Основным процессом, обеспечивающим передвижение воды по растению, является транспирация [3]. При этом устьичная транспирация составляет 80–90 % от всего испарения листа. Следовательно, количество устьиц, их размеры и размещение могут являться фиксированным источником информации о процессах водообмена. Они в первую очередь реагируют на изменение условий произрастания в процессе роста листьев. Рост листьев и формирование устьичного аппарата происходят сопряженно с формированием водопроводящей системы у древесных растений, включая процессы отложения новых клеток флоэмы и ксилемы. Эти процессы скоординированы в сезонном цикле развития древесного растения, поскольку устьичная проводимость и транспирация воды и газов зависят от проводимости ксилемы с одной стороны, а питательные вещества и сигнальные гормоны, которые определяют формирование водопроводящих структур ксилемы, поставляются через флоэму [6].
Выявлена тесная корреляция между площадью проводящей ксилемы и массой или поверхностью хвои сосны обыкновенной (коэффициент детерминации до 0,99) [4] и вяза мелколистного (до 0,98) [10]. Установлена зависимость между состоянием дерева и строением водопроводящей системы (так, чем лучше состояние деревьев вяза, тем больше концентрически расположенных мелких сосудов отложено в поздней древесине [11]. Некоторые интересные закономерности, установленные для травянистых растений, например нетипичная реакция пшеницы (в ответ на снижение влажности воздуха, но в условиях высокой влажности почвы у некоторых сортов увеличивается устьичная проводимость, что обеспечивает рост продуктивности) [7], могут найти применение при плантационном лесовыращивании.
В то же время отсутствуют работы, в которых представлены результаты исследования связи водопроводящей системы ксилемы с морфологией листа. Особое значение поиск данных закономерностей имеет для лиственных древесных растений (кольцесосудистых, рассеяннососудистых), в частности в целях индикации природных изменений с использованием длительных древесно-кольцевых хронологий. Учитывая исключительно высокую чувствительность устьичного аппарата на изменения погоды в течение всего вегетационного периода и способность ксилемы фиксировать изменения условий произрастаний при установленных устойчивых аллометрических связях, возможно через изучение ксилемы получить информацию об условиях произрастания ретроспективно за весь период жизни дерева.
Цель и задачи исследования. Установить закономерности ответной реакции устьичного аппарата и ксилемы древесных растений на условия произрастания и климатические изменения для индикации природных аномалий по длительным древесно-кольцевым хронологиям.
Материалы и методы. В качестве объектов исследования выбраны древесные растения, произрастающие в дендрариях Института леса, СибГТУ, заложенные в 50–60-х годах ХХ века. Метеорологические данные для анализа влияния условий произрастания на анатомо-морфологические параметры древесных колец в течение текущего вегетационного периода и ретроспективно получены как в Среднесибирском управлении по гидрометеорологии и мониторингу окружающей среды, так и в результате собственных измерений и мониторинга. В исследование включены кольцесосудистые, такие как дуб монгольский ( Quercus mongolica Fisch. ex Ledeb.) и дуб черешчатый ( Quercus robur L .) (виды, отличающиеся динамикой накопления фитомассы и устойчивостью к болезням и вредителям), вяз приземистый ( Ulmus pumila L.) , барбарис амурский ( Berberis amurensis Rupr.) ; рассеяннососудистые, такие как ива корзиночная ( Salix viminalis L.) , липа мелколистная ( Tilia cordata Mill.) и хвойные, такие как сосна обыкновенная ( Pinus sylvestris L .) и кедр сибирский ( Pinus sibirica Du Tour ). Ранее полученный комплекс научных результатов по кольцесосудистым древесным растениям и также факт, что в проведении воды участвует только последний годичный слой из-за затилловывания сосудов (листья данного года обслуживаются сосудами того же года), делают их интересным объектом исследования взаимоотношения в системе устьица-ксилема. В то же время хвойные растения интересны в исследовании с позиции соотношения ранней и поздней древесины в отношении к анатомии и морфологии устьиц. Дизайн эксперимента выполнен факторными планами с блоками, что обеспечивает моделирование процессов роста и формирования ксилемы с более высокой точностью посредством возможности фильтрования случайного шума с использованием смешанных моделей.
Исследование системы устьица-ксилема производится в контрастных условиях увлажнения. Для этих целей заложен оригинальный эксперимент. In vivo в идентичных условиях освещения, влажности воздуха сформированы два режима увлажнения почвы. Контроль – естественный режим увлажнения. Аридные условия – посадочные контейнера с дренажными отверстиями оторваны от уровня почвы на 25 см (рис.1). Высажены клоновые саженцы ивы корзиночной, сеянцы дуба монгольского, дуба черешчатого, кедра сибирского, липы мелколистной, барбариса амурского. Принцип лимитирования позволяет количественно описать отклик ксилемы как формирующейся ткани, контролируемой внешними и внутренними факторами, в ответ на изменение этих факторов. Отклик ксилемы на климатические условия произрастания оценен при помощи методов дендроклиматологии, таких как удаление возрастного тренда и анализ корреляционных связей и функций отклика [18].
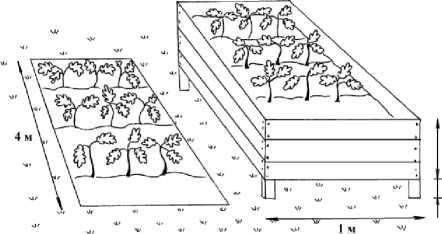
Рис.1. Схема размещения растений
Для определения биометрии устьиц (их размеров и количества) использована система анализа изображений (Image System Analyser) за исключением сложной предварительной подготовки по методам Ллойда и Молотковского. Ярусность является важным параметром, поскольку отражает информацию об условиях произрастания с высоким разрешением (до нескольких дней) в течение всего вегетационного периода, так как на распределение устьиц и их размеры, кроме внутренней архитектуры листа и расположения листьев, оказывают значительное влияние экзогенные факторы (интенсивность освещения, влажность воздуха и почвы, температура, содержание углекислого газа в атмосфере, обеспеченность элементами питания). Оценка условий произрастания также возможна благодаря наличию устьичного индекса (постоянная величина не только для листьев одного побега или всего растения, но и для всех особей какого-либо вида, произрастающего в тех же условиях). Все это позволяет проводить сопряженные исследования (ксилемы и устьичного аппарата) для оценки влияния внешних и внутренних факторов на рост древесных растений. В дальнейшем для каждого яруса листьев были выделены участки годичного прироста, сформировавшиеся одновременно с ними. Полученные при помощи санного микротома МС-2 тонкие поперечные срезы древесины в дальнейшем были окрашены водным раствором метиленового синего и помещены под покровные стекла в глицерин. Для определения биометрических показателей использовалась система анализа изображений (Image System Analyser).
Результаты и обсуждение. Учитывая то, что основная масса листового аппарата образуется в начале вегетационного периода, для того, чтобы обеспечить фитомассу достаточным количеством воды, требуется мощная проводящая система. Во второй половине вегетационного периода приоритет отдается формированию ксилемы дуба, отвечающей за обеспечение механических функций. Именно поэтому было и принято решение исследовать соотношение площади сосудов и площади ксилемы, сформировавшейся в период наиболее активной дифференциации клеток, а именно начиная со II декады мая до I декады июня. Именно в этот период у кольцесосудистых древесных растений образуется наиболее густая сеть из крупных сосудов [2]. При произрастании дуба монгольского и черешчатого на открытых хорошо освещенных участках с большой площадью питания каждого растения формируется ксилема с большей площадью сосудов. Дисперсионный анализ подтверждает вывод о значимом влиянии условий освещения на формирование сосудов ксилемы (р<0,05). Под пологом лиственного древостоя в условиях недостаточного увлажнения и освещения формируются устьица меньшего размера и уменьшается их плотность (табл. 1). Данная закономерность характерна для всех ярусов листьев. В результате проведенного дисперсионного анализа подтверждено влияние условий произрастания и индивидуальных особенностей вида растений на формирование устьичного аппарата (р<0,001).
Характеристика устьичного аппарата (фрагмент)
Таблица 1
|
Условия произрастания |
Дуб монгольский |
Дуб черешчатый |
||||
|
1** |
2** |
3** |
1 |
2 |
3 |
|
|
а* |
445,50 |
24,33 |
18,08 |
400,61 |
26,57 |
19,78 |
|
б* |
359,62 |
22,48 |
17,41 |
276,34 |
18,31 |
13,78 |
|
Критерий существенности различия |
6,06 |
8,25 |
4,6 |
5,79 |
13,95 |
27,7 |
|
Текущ |
ий прирост |
|||||
|
а* |
485,03 |
25,00 |
18,49 |
416,62 |
27,76 |
20,17 |
|
б* |
387,60 |
23,20 |
17,67 |
290,68 |
18,82 |
13,91 |
|
Критерий существенности различия |
5,01 |
8,34 |
4,73 |
4,39 |
8,07 |
7,43 |
|
1-й ярус |
||||||
|
а* |
447,27 |
23,74 |
17,60 |
404,93 |
26,89 |
19,47 |
|
б* |
353,69 |
22,52 |
17,16 |
248,95 |
18,45 |
13,82 |
|
Критерий существенности различия |
4,5 |
6,5 |
3,2 |
12,5 |
6,3 |
4,7 |
|
2-й ярус |
||||||
|
а* |
409,16 |
24,86 |
18,36 |
361,54 |
26,70 |
20,62 |
|
б* |
336,85 |
21,95 |
17,17 |
295,74 |
18,97 |
14,55 |
|
Критерий существенности различия |
5,1 |
4,7 |
4,5 |
6,1 |
6,7 |
6,4 |
|
3-й ярус |
||||||
|
а* |
440,53 |
23,70 |
17,87 |
419,35 |
24,94 |
18,85 |
|
б* |
360,35 |
22,26 |
17,65 |
270,01 |
17,01 |
12,83 |
|
Критерий существенности различия |
6,5 |
3,4 |
3,2 |
9,3 |
10,8 |
10,4 |
|
4-й ярус |
||||||
|
а* |
428,13 |
26,76 |
18,24 |
379,43 |
24,65 |
21,45 |
|
б* |
354,77 |
23,54 |
17,34 |
313,01 |
18,87 |
15,33 |
|
Критерий существенности различия |
5,3 |
3,7 |
4,8 |
5,9 |
5,8 |
6,5 |
|
5-й ярус |
||||||
|
а* |
445,35 |
22,80 |
18,17 |
424,38 |
23,74 |
17,39 |
|
б* |
364,57 |
21,12 |
17,14 |
275,32 |
16,21 |
12,92 |
|
Критерий существенности различия |
6,4 |
3,2 |
3,4 |
8,4 |
9,8 |
10,1 |
Примечание. *: а - условия полного освещения; б - под пологом сомкнутого лиственного древостоя; **: 1 - плотность устьиц; 2 - длина устьиц; 3 - ширина устьиц.
В результате совмещения результатов исследования устьичного аппарата и ксилемы была установлена устойчивая связь между площадью проводящих сосудов в ксилеме (в процентах от общей пл о-щади, взятой для исследования), сформировавшейся в период наиболее активной дифференциации клеток – с одной стороны, и биометрическими параметрами устьичного аппарата – с другой (табл.2).
Таблица 2
Коэффициенты корреляция между биометрическими показателями устьичного аппарата и площадью сосудов ксилемы
|
Показатель |
Площадь сосудов ксилемы |
|
|
Дуб монгольский |
Дуб черешчатый |
|
|
Длина устьиц |
0,5* |
0,6* |
|
Ширина устьиц |
0,51* |
0,59* |
|
Плотность устьиц |
0,67** |
0,66* |
Примечание. * – р< 0,05; ** – р< 0,001.
В режиме искусственной засухи (на гряде) происходит снижение всех ростовых процессов (рис.2). Установлено снижение массы корней и стволика в посадочных контейнерах (в большей степени для дуба монгольского) (рис. 3).
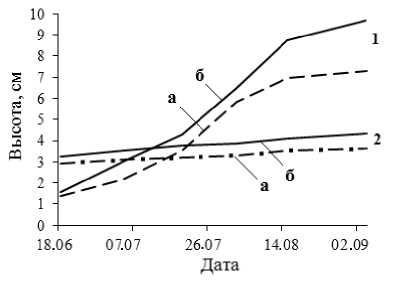
Рис. 2. Динамика роста по высоте стволика саженцев липы мелколистной (1) и дуба монгольского (2) в условиях искусственной засухи (а) и в контроле (б)
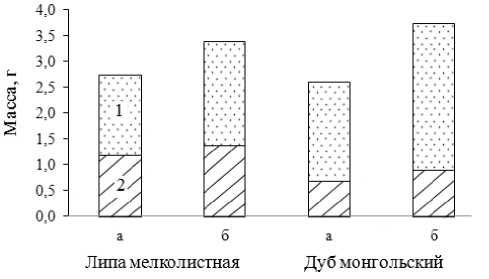
Рис. 3. Масса сеянцев в воздушно-сухом состоянии (1 – стволика; 2 – корней) в условиях искусственной засухи (а) и в контроле (б)
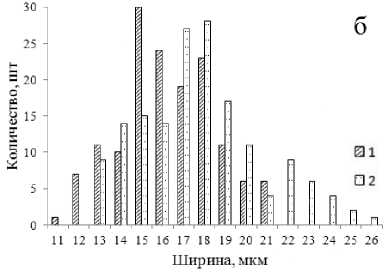
Дефицит влаги привел к изменению биометрических параметров устьичного аппарата. Произошла его ксерофитизация – уменьшился размер устьиц, увеличилось их количество (табл. 3). На рисунках 4–5 показана гистограмма распределения устьиц по длине и ширине. Для распределения устьиц контрольных саженцев (в оптимальных условиях увлажнения) характерна выраженная положительная асимметрия.
Талица 3
Биометрическая характеристика устьиц в разных условиях произрастания
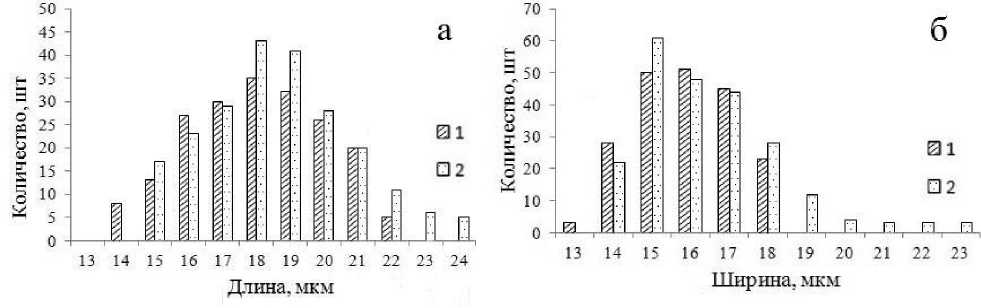
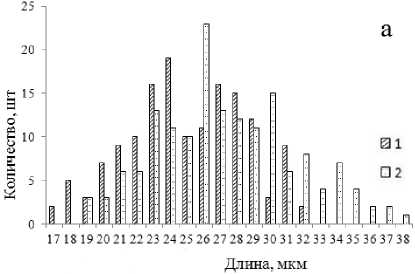
Рис. 5. Распределение устьиц липы мелколистной по длине (а) и ширине (б) (1 – условия искусственной засухи; 2 – контроль)
Рис. 4. Распределение устьиц дуба монгольского по длине (а) и ширине (б) (1 – условия искусственной засухи; 2 – контроль)
|
Условия произрастания |
Длина, мкм |
Ширина, мкм |
Количество, шт/мм |
|||
|
1 |
2 |
1 |
2 |
1 |
2 |
|
|
Контроль |
18,1±0,14 |
26,9±0,33 |
16,1±0,13 |
17,2±0,22 |
659±21,4 |
343±19,8 |
|
Условия искусственной засухи |
17,5±0,13 |
24,5±0,29 |
15,3±0,08 |
15,8±0,18 |
766±28,7 |
422±25,6 |
Примечание. 1 – дуб монгольский; 2 – липа мелколистная.
Заключение. Установлена высокая чувствительность устьичного аппарата древесных растений к условиям произрастания (тип условий местопроизрастания, положение в пологе и пр.) и климатическим изменениям (температура, осадки). В условиях искусственной засухи снижается не только биомасса саженцев, но и происходит ксерофитизация устьичного аппарата. Выявлен важный методический прием: для оценки условий произрастания (в частности, освещения) целесообразно использовать прирост ксилемы в период активной дифференциации клеток (вторая декада мая – первая декада июня). Установлена устойчивая связь между площадью проводящих сосудов в ксилеме, сформировавшейся в период наиболее активной дифференциации клеток, и биометрическими параметрами устьичного аппарата (длина и ширина устьиц, плотность).
