Анатомо-топографические характеристики и цитоархитектоника тройничного узла у взрослого человека

Автор: Гайворонский И.В., Шангина Л.В., Пащенко П.C., Забродская Ю.М.
Журнал: Вестник медицинского института "РЕАВИЗ": реабилитация, врач и здоровье @vestnik-reaviz
Рубрика: Морфология. Патология
Статья в выпуске: 4 т.15, 2025 года.
Бесплатный доступ
Введение. В многочисленных отечественных и зарубежных источниках строение тройничного узла человека и животных изучалось с топографо-анатомических, гистологических, эмбриологических, хирургических, нейрофизиологических и патологоанатомических позиций. Известно, что форма тройничного узла вариабельна, большинство авторов считают, что он имеет полулунную форму. Внутреннее строение сводится в основном к характеристике клеточной составляющей узла, а также проходящих через него нервных волокон. В тройничном ганглии человека отношение нейронов к глии равняется 1:100. Считается, что Гассеров узел может содержать от 20 до 35 тыс. нейронов. Кроме того, в узле содержатся фибробласты, а также мелкие сосуды, преимущественно кровеносные капилляры. Цель исследования – уточнение анатомотопографических характеристик и структурной организации, а также морфометрических особенностей нейронов тройничного узла. Материал и методы. Объектом исследования являлись четыре тройничных узла с чувствительным корешком и ветвями, отходящими от него, с сохранённой твердой мозговой оболочкой, взятые от трупов взрослых мужчин второго периода зрелого возраста (60–65 лет), без признаков хирургических вмешательств и травм. Из отобранного материала изготовлены срезы, окрашенные по методикам Ниссля, Ван-Гизона, а также гематоксилином и эозином. Результаты. В соответствии с особенностями структурной организации вычленены структурно-функциональные зоны составляющих частей Гассерова узла (зона трифуркации, промежуточная зона тройничного узла, зона формирования чувствительного корешка). Обращает на себя внимание наибольшая плотность распределения тел нейронов, ориентированных в направлении верхнечелюстной и нижнечелюстной ветвей тройничного нерва, что, как мы предполагаем, объясняется большей площадью рецептивных полей, связанных с ними ветвей. В зоне формирования чувствительного корешка выявляются преимущественно крупные нейроны, разделённые на группы нервными волокнами, образующие пучки I, II и III порядков, а при слиянии – чувствительный корешок. Выводы. Тройничный узел характеризуется локальными особенностями нейроцито- и миелоархитектоники, исследование которых должно проводиться с учётом половых и возрастных характеристик.
Тройничный нерв [D014276], тройничный узел [D012668], Гассеров узел [D012668], морфометрия [D009055], нейроны [D009474], топографическая анатомия [D000066477], гистология [D006653], нервная система [D009420], черепные нервы [D003391], сенсорные ганглии [D00572]
Короткий адрес: https://sciup.org/143185007
IDR: 143185007 | УДК: 611.831.31:616.833.15 | DOI: 10.20340/vmi-rvz.2025.4.MORPH.6
Anatomical and topographic characteristics and cytoarchitectonics of the trigeminal node in an adult
Introduction. In numerous domestic and foreign sources, structure of the trigeminal node of human and animals have been studied from topographic, anatomical, histological, embryological, surgical, neurophysiological and pathoanatomical positions. It is known, the shape of the trigeminal node is variable, most authors believe that it has a semilunar shape. The internal structure is mainly reduced to the characteristics of the cellular component of the node, as well as the nerve fibers passing through it. In the human trigeminal ganglion, the ratio of neurons to glia is 1:100. It is believed that a Gasser node can contain from 20 to 35 thousand neurons. In addition, the node contains fibroblasts, as well as small vessels, mainly blood capillaries. The aim of the study is to clarify the anatomical and topographic characteristics and structural organization, as well as the morphometric features of trigeminal neurons. Materials and methods. The object of the study was 4 trigeminal ganglia with a sensitive root and branches extending from it, with preserved dura mater, taken from corpses of adult men of the second period of adulthood (60-65 years old), without signs of surgical interventions and injuries. Sections were made from the selected material, stained according to the Nissl, Van Gieson methods, as well as with hematoxylin and eosin. Results. In accordance with the peculiarities of the structural organization, the structural and functional zones of the constituent parts of the Gasser node (the trifurcation zone, the intermediate zone of the trigeminal node, the zone of formation of the sensitive root) are isolated. Attention is drawn to the highest density of distribution of neuron bodies oriented in the direction of the maxillary and mandibular branches of the trigeminal nerve, which, as we assume, is explained by the larger area of receptive fields associated with these branches. In the zone of formation of the sensory root are seen mainly large neurons, divided into groups by nerve fibers, forming bundles of orders I, II and III, which merge to form a sensitive root. Conclusions. The trigeminal ganglion is characterized by local features of neurocyto- and myeloarchitectonics, the study of which should be carried out taking into account gender and age characteristics.
Текст научной статьи Анатомо-топографические характеристики и цитоархитектоника тройничного узла у взрослого человека
В имеющихся многочисленных отечественных и зарубежных источниках положение и строение тройничного узла человека и животных изучалось с топографо-анатомических, гистологических, эмбриологических, хирургических, нейрофизиологических и патологоанатомических позиций [1–14].
Известно, что тройничный узел (ТУ) человека располагается в тройничной (Меккелевой) полости [9], образуя нервное сплетение треугольной формы [6].
Меккелева полость (МП) находится в тройничном вдавлении между менингеальным слоем (dura propria) и твердой мозговой оболочкой, прочно прикреплённой к кости в области верхушки пирамиды [8]. Внутри МП паутинная оболочка образует карман [10].
От рострально-латерального края ТУ отходят три крупных нерва: глазной, верхнечелюстной и нижнечелюстной. Каудальный край узла продолжается в чувствительный корешок тройничного нерва, содержащий центральные отростки нейронов, которые затем входят в ствол мозга на уровне моста.
Форма ТУ вариабельна, большинство авторов считают, что он имеет полулунную форму. Другими исследованиями была показана зависимость формы и размеров данного узла от индивидуальных особенностей черепа. Для брахиоцефалического типа характерен крупный узел (короткий и высокий), для долихоцефалического – мелкий узел (длинный и узкий) [2].
Ещё в 20–40 гг. прошлого столетия возникла дискуссия в отношении развития ТУ. Одни авторы считали, что в процессе закладки узла формируется сплошное образование, связанное широкой ножкой со стволом мозга, другие признают развитие узла частями [4].
Рассмотрение дифференцировки клеток ТУ методом кариометрии позволило исследователям выявить гетерохронию в развитии офтальмического, мандибулярного и максиллярного участков узла [11]. Авторы признают дифференцировку узла на участки уже на ранних этапах эмбриогенеза. С использованием различных гистологических, гистохимических, иммунногистохимических методов направляются усилия на детальное изучение его структурных компонентов (сосудов, вегетативных сплетений и т.д.) и значительно в меньшей мере рассматриваются общие вопросы структурной организации ТУ, его отдельных участков в процессе онтогенеза [1, 11]. Что же касается функциональных зон узла, а также их структурных особенностей у людей, то в последние годы такие работы единичны [7]. Это находится в сфере запросов нейрохи- рургии, т.к. тормозит развитие новых инвазивных технологий диагностики, оказания помощи больным с патологическими изменениями ТУ, его чувствительного корешка, а также отходящих от узла нервов.
Целью работы является изучение анатомотопографических характеристик, структурной организации и морфометрических особенностей нейронов тройничного узла человека II периода зрелого возраста.
Материал и методы
Объектом исследования являлись четыре тройничных узла с корешком и ветвями, с сохранённой твердой мозговой оболочкой, взятые от трупов взрослых людей второго периода зрелого возраста (60–65 лет). В материале отсутствуют признаки хирургических вмешательств, травм.
Методика забора материала. Производили разрез ниже верхнего края пирамиды височной кости, вдоль медиального края верхней глазничной щели отслаивали твердую мозговую оболочку вместе с содержимым. Отсечение первой ветви тройничного нерва выполнено максимально близко к верхней глазничной щели, затем пересекали вторую ветвь у круглого отверстия и третью ветвь у входа в овальное отверстие. После отслоения твердой мозговой оболочки от тройничного вдавления узел извлекали с корешком и ветвями, покрытыми твёрдой мозговой оболочкой. Отобранный материал фиксировали в течение 5 суток в 10% нейтральном растворе формалина, фиксирующую жидкость периодически меняли. Материал после проводки заключали в парафиновые блоки, из которых в дальнейшем были приготовлены срезы толщиной порядка 5 мкм на микротоме Leica RM2235. Срезы после депарафинирования окрашивали по методикам Ниссля, Ван-Гизона, гематоксилином и эозином. Микрофотографии выполняли с помощью микроскопа Leica DM2500, оснащённого цветной цифровой камерой Leica DFC295. Исследование плотности распределения исследуемых структур в окрашенных срезах проводили с помощью морфометрической сетки В.С. Сидорина. Размеры (площади) тел нейронов, их ядер изучали с помощью программного обеспечения SlideViewer Version 2.6.
Результаты и их обсуждение
Рассмотрение препаратов, окрашенных гистологическими методиками для световой микроскопии, показало, что МП представляет собой полое пространство, нижняя стенка (дно) которого имеет костную основу в виде тройничного вдавления на передней поверхности каменистой части височной кости. С медиальной стороны вдавление ограничено верхушкой пирамиды височной кости, а лате-рально оно достигает дугообразного возвышения. Указанная особенность топографии тройничного узла имеет исключительно важное значение в пункционном доступе, поскольку дугообразное возвышение является опознавательным образованием при оперативном вмешательстве и вскрытии его капсулы [5]. Вдавление покрыто приросшей периостальной пластинкой твёрдой мозговой оболочки, выполняющей в этом месте функции надкостницы. Кнутри от неё обнаруживается тонкий слой паутинной оболочки (рис. 1).
В непосредственной близости от тройничного вдавления, а значит, от ТУ, находятся важнейшие анатомические образования:
-
- впереди и латерально - пещеристый синус и внутренняя сонная артерия;
-
- медиально и позади - верхний каменистый синус;
-
- латерально и сзади - средняя оболочечная артерия, принимающая участие в кровоснабжении ТУ;
-
- у верхушки пирамиды - задний межпещеристый синус.
Указанные топографо-анатомические отношения ТУ имеют клиническое значение как для понимания возможности перехода патологического процесса на узел и его ветви, так и опасности проведения хирургических вмешательств в достаточно узкой области, насыщенной перечисленными анатомическими образованиями.
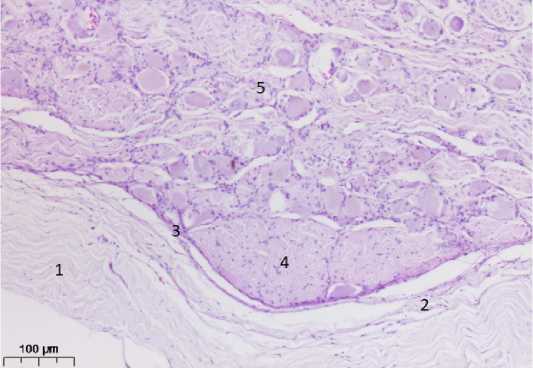
Рисунок 1. Каудальная часть тройничного узла. Окраска гематоксилином и эозином, ув. 100: 1. Твердая мозговая оболочка. 2. Паутинная оболочка. 3. Капсула узла. 4. Фрагмент внутри-узловой части чувствительного корешка. 5. Паренхима узла
Figure 1. Caudal part of the trigeminal node. H&E, x100: 1. Dura mater. 2. The spider web. 3. Node capsule. 4. Fragment of the internode part of the sensitive root. 5. Parenchyma of the node
Нами предпринята попытка определить участки (зоны) узла, исходя из их функциональных и морфологических особенностей.
Так, периферический участок узла, нейроны которого ориентированы в направлении I, II, III ветвей V пары, можно рассматривать в качестве мест вхождения нервных импульсов в мононейронную сенсорную систему, представленную псевдоунипо-лярными нервными клетками (сенсорный вход, рис. 2).
Противоположный (каудальный) участок ТУ является местом конвергенции импульсов, распространяющихся по центральным отросткам псевдо-униполярных нейронов в направлении формирующегося чувствительного нервного корешка.
Между этими участками находится промежуточная зона нейронов, которая выполняет функцию приёма импульсов, их первичный анализ, а также распределение по центральным отросткам псев-доуниполярных нейронов (сенсорнораспределительная зона), ориентированных в сторону формирующегося чувствительного корешка тройничного нерва (рис. 3).
Рассмотренные с функциональных позиций участки узла имеют также свои особенности организации. Так, в участке, непосредственно примыкающем к переднему полюсу с входящими в узел ветвями (место «сенсорного входа»), было выявлено разделение нейронов на три группы (трифуркация). Будучи отделёнными между собой соединительной тканью и пучками нервных волокон, каждая из групп оказалась ориентированной в направлении одной из входящих в узел ветвей (нервов).
Группы нервных клеток ТУ, направленных к I, II, III ветвям тройничного нерва, обозначены нами как глазная (гр. I), верхнечелюстная (гр. II) и нижнечелюстная (гр. III) посттрифуркационные зоны.
Нервные волокна в указанных зонах образуют пучки, пронизывающие скопления нейронов в каждой отдельной зоне. Следовательно, есть основание полагать, что описанные в раннем эмбриогенезе участки в закладке ТУ, заселённые нейробластами, в постанатальном онтогенезе человека содержат группы псевдоуниполярных нейронов, периферические отростки которых в процессе развития приобрели связи с соответствующими рецептивными полями. Значение разделения данных участков узла на зоны заключается в возможности дифференцировать группы нервных клеток и волокон, из которых построена начальная часть отдельно взятой ветви (чувствительного нерва) V пары ЧН.
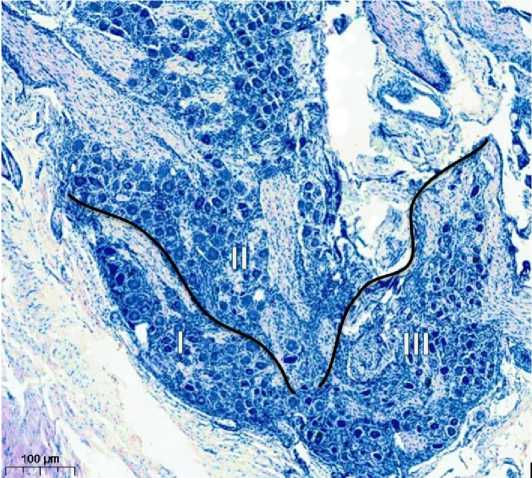
Рисунок 2. Периферическая часть тройничного узла. Окраска по Нисслю, ув. 100. Группы нейронов, разделенные между собой соединительной тканью и пучками нервных волокон, ориентированные в направлении вхождения в узел ветвей (глазного – I, верхнечелюстного – II, нижнечелюстного нервов – III)
Figure 2. Peripheral part of the trigeminal node. Nissl staining, ×100. Groups of neurons, separated by connective tissue and bundles of nerve fibers, oriented in the direction of entry into the node of the branches (ocular – I, maxillary – II, mandibular nerves – III)
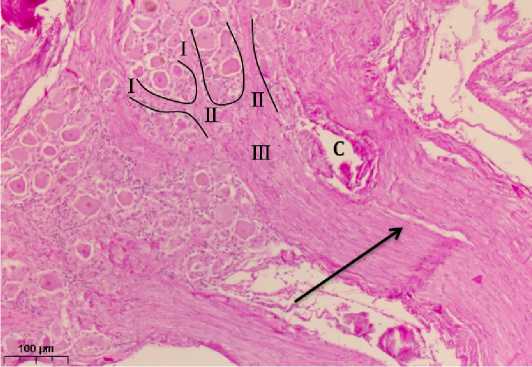
Рисунок 3. Каудальный участок тройничного узла в месте интра-ганглионарного формирования чувствительного корешка тройничного нерва. Окраска по Ван-Гизону, ув. 500. I – пучки первого порядка; II – пучки второго порядка, образующиеся путем слияния пучков первого порядка; III – пучок третьего порядка, формирующийся из пучков второго порядка. Фрагмент чувствительного корешка (стрелка). С – сосуд
Figure 3. Caudal section of the trigeminal node at the site of intra-ganglionic formation of the sensitive root of the trigeminal nerve. Van Gieson staining, ×500. I – first–order beams; II – second–order beams formed by merging first-order beams; III – third-order beam formed from second-order beams. A fragment of the sensitive root (arrow). C is a vessel
В указанных группах плотность распределения тел нейронов не одинакова. Так, если в гр. I относительное содержание нервных клеток составляет 6,5% от площади данной зоны, то в гр. II этот показатель составил 24,4%, а в гр. III – 17,5%. Значительный процент площади, занимаемой телами нейронов в гр. II и гр. III, может объясняться большей площадью их рецептивных полей по сравнению с рецептивным полем, связанным с телами нейронов при помощи их периферических отростков в гр. I.
Промежуточная зона ТУ, расположенная между трифуркацией и началом формирующегося внутри неё чувствительным корешком, состоит из тел псевдоуниполярных нейронов, окружённых глиальной капсулой, их центральных и, связанных с рецептивными полями, периферических отростков. Следовательно, в зоне обеспечивается как приём, так и передача импульсов в направлении чувствительных ядер V пары.
Нейроны этой зоны преимущественно крупные, они образуют структурно-функциональные группы, внутри которых формируются соразмерные по толщине пучки нервных волокон – пучки I порядка, поперечный размер которых находится в диапазоне от 20,1 до 42,9 мкм.
Конвергируя между собой они составляют более толстые пучки (II порядка) с поперечным размером от 47,8 до 54,1 мкм. Как следует из рисунка 4, максимальная площадь, занимаемая телами нервных клеток и глии находится в начальном отделе промежуточной зоны, между трифуркацией и началом формирования нервных пучков I порядка. По мере формирования пучков I-го, а затем и II-го порядков, величина этого показателя постепенно уменьшается, при этом относительная площадь нервных волокон, наоборот, увеличивается за счёт прибавления центральных отростков псевдоуни-полярных нейронов из заднего отдела ТУ.
В целом, во всех исследованных участках узла нервные волокна занимают большую площадь, чем тела нейронов. Этот факт, вероятно, можно объяснить тем, что кроме нервных волокон, являющихся отростками нейронов узла, в ганглии присутствуют афферентные волокна центрального происхождения, а именно периферические отростки псевдо-униполярных нейронов среднемозгового ядра [12].
Пучки II порядка соединяются между собой, образуя толстые тяжи, разделённые и окружённые соединительной тканью. Их поперечный размер составляет от 149,3 до 284 мкм. После их слияния образуется чувствительный корешок шириной более 300 мкм, формирование которого как единого образования происходит внутри каудального полюса МП. Нервные клетки в таких тяжах отсутствуют или же они единичны.
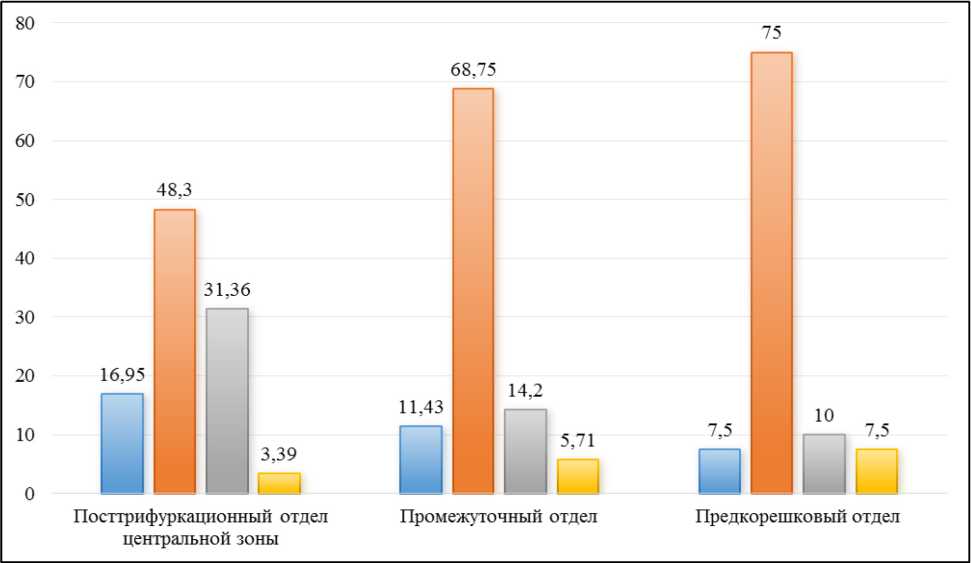
Рисунок 4. Распределение структурных компонентов тройничного узла на протяжении промежуточной зоны узла, %
Figure 4. Distribution of the structural components of the trigeminal node throughout the central zone of the node, %
Тела нейронов тройничного узла в исследованных зонах имеют округлую или овальную форму. Все они одноядерные. Ядро клетки находится в центре цитоплазмы или близко к нему. Оно сферической формы, в кариоплазме контурируется одно (реже два) ядрышка, расположенное в центре ядра.
Диапазон размеров (площадей) тел нейронов достаточно широк – от 297,7 до 3697,3 мкм2. Нами выделены три группы нейронов по их размерам: малые, средние и крупные (табл. 1).
Исследование ядерно-цитоплазматического отношения выявило близкие значения данного показателя в больших (0,11) и средних (0,13) нейроци-тах, что может быть свидетельством однородности этих клеток. В сумме крупные и средние нейроны составили 66,8% всей нервно-клеточной совокупности тройничного узла. В мелких нейронах ядер-но-цитоплазматического отношение оказалось более высоким, составив при этом 0,18 (табл. 2).
Выявлены отдельные клетки, у которых площадь ядра превалировала над площадью цитоплазмы, в результате показатель ядерно-цитоплазматического отношения достигал 3,9.
Следует отметить, что в описанных выше зонах (I, II, III группах) распределение крупных, средних и мелких нейронов имеет различные количественные характеристики (табл. 3).
Таблица 1. Морфометрические показатели тел нейронов тройничного ганглия у людей пожилого возраста
Table 1. Morphometric parameters of trigeminal ganglion neuron bodies in the elderly
|
Размеры нейронов |
S*, µм2, X±mx |
S min – S max, µм2 |
Медиана |
|
Крупные |
2225,7±73,3 |
1557,2–3697,3 |
2169,3 |
|
Средние |
1147,3±26,5 |
905,2–1553,4 |
1146,2 |
|
Мелкие |
717,5±22,6 |
297,7–901,9 |
749 |
Примечание: *S – площадь тела нейрона.
Таблица 2. Усредненное отношение площади ядер к площади цитоплазмы в крупных, средних и мелких нейронах тройничного узла Table 2. The average ratio of the area of the nuclei to the area of the cytoplasm in large, medium and small trigeminal neurons
|
Размеры нейронов |
S цитоплазмы, µм2 X±mx |
S ядра, µм2 X±mx |
ЯЦО*, отн. ед. X±mx |
|
Крупные |
2017,1±70,4 |
220,6±8,5 |
0,11±0,01 |
|
Средние |
993±30,6 |
154,3±16,9 |
0,13±0,07 |
|
Мелкие |
609,2±19,9 |
110,3±4,6 |
0,18±0,007 |
Примечание: *ЯЦО – ядерно-цитоплазматическое отношение (nuclear–cytoplasmic ratio).
Таблица 3. Распределение мелких, средних и крупных нейронов в посттрифуркационной зоне тройничного узла, %
Table 3. Distribution of small, medium, and large neurons in the posttrifurcation zone of the trigeminal node, %
|
Размер нейрона |
Глазной нерв |
Верхнечелюстной нерв |
Нижнечелюстной нерв |
|
Крупные |
5,5 |
69,6 |
33,3 |
|
Средние |
38,9 |
17,4 |
40,0 |
|
Мелкие |
55,6 |
13 |
26,7 |
Исходя из структурных особенностей цитоплазмы и ядра нам представилось возможным выделить четыре группы нервных клеток:
-
1. Нейроны, в которых не выявлено структурных проявлений реактивных и патологических изменений (рис. 5А).
-
2. Нервные клетки с признаками центрального и периферического хроматолиза.
-
3. Нейроны, содержащие в цитоплазме липофусцин (рис. 5Б).
-
4. Нейроциты с выраженными признаками дегенерации цитоплазмы и ядра (рис. 5В и 5Г).
Вторая группа нейронов отличается от предыдущей наличием в периферических или центральных (иногда в центральных и периферических) отделах цитоплазмы разукрупнение базофильных гранул (зерен), их распыление. В этих участках цитоплазма приобретает более бледную окраску, что является признаком хроматолиза (хроматолизиса) – периферического и (или) центрального – структур- ного признака повышенной активности нервной клетки. Следовательно, нейроны I и II групп имеют все морфологические признаки зрелых, функционально полноценных клеток.
Для нейронов третьей группы характерным является развитие в них липофусциногенеза, проявляющегося накоплением в центральных и периферических участках цитоплазмы пигмента липофусцина: в одних нейронах пигмент рассеян между гранулами базофильного вещества в небольшом количестве (это первая – начальная, диффузная стадия липофусциногенеза). Она превалирует в цитоплазме нейронов крупных и средних размеров.
В других нервных клетках липофусцин в виде небольших скоплений (очагов) наблюдается в око-лоядерной зоне (рис. 5Б) – это стадия концентрации липофусцина. Некоторые авторы считают, что процесс липофусциногенеза в виде накопления пигмента начинается с околоядерной зоны [5, 7].
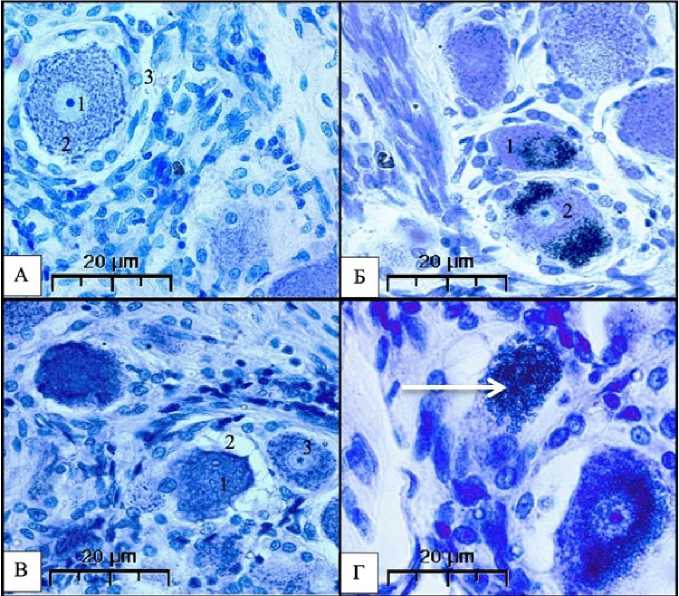
Рисунок 5. Различные формы нейронов и клеток глии в тройничном узле мужчин 60–65 лет. Окраска по Нисслю. А – Тело нейрона с обычным строением ядра и цитоплазмы (ув. 500) (1 – ядро с ядрышком; 2 – цитоплазма, заполненная равномерно расположенными базофильными гранулами; 3 – клетки перинейрональной глии); Б – Нейрон со скоплениями липофусцина в цитоплазме (ув. 500) (1 – околоядерная форма; 2 – двухполюсная форма); В – вакуольная дистрофия (ув. 500) (1 – нейрон со светлыми вакуолями на периферии цитоплазмы; 2 – зона глиоза; 3 – нейрон с обычным строением ядра и цитоплазмы); Г – Дегенерация нейрона, отсутствие ядра, тотальное заполнение клетки липофусцином (стрелка) (ув. 600)
Figure 5. Different shapes of neurons and glial cells in the trigeminal ganglion of 60-65 year old men. Nissl staining. A is the body of a neuron with the usual structure of the nucleus and cytoplasm (500x) (1 is the nucleus with a nucleolus; 2 is the cytoplasm filled with evenly spaced basophilic granules; 3 are perineuronal glial cells); B is a neuron with accumulations of lipofuscin in the cytoplasm (500x) (1 is the perinuclear form; 2 is bipolar form); V – vacuolar dystrophy (500x) (1 – neuron with light vacuoles on the periphery of the cytoplasm; 2 – gliosis zone; 3 – neuron with the usual structure of the nucleus and cytoplasm); G – Degeneration of a neuron, absence of a nucleus, total filling of a cell with lipofuscin (arrow) (×600)
Встречаются нейроны, у которых значительные скопления липофусцина занимают один из полюсов цитоплазмы. Авторы называют эту стадию полярной [14].
В некоторых клетках очаги пигмента были отмечены на противоположных полюсах цитоплазмы нейронов (в крупных и мелких нейронах), что свидетельствует о его дальнейшем распространении (биполярная стадия). В отдельных клетках липофусцин занимает пространство между ядром и одним из полюсов цитоплазмы или же выполняют цитоплазму и ядро.
Распределение липофусцина в нейронах различных размеров является мозаичным (табл. 4).
Таблица 4. Формы распределения липофусцина в клетке, % Table 4. Forms of lipofuscin distribution in the cell, %
|
Нейроны |
Форма распределения |
||||
|
рассеянная |
около-ядерная |
полярная |
двухполярная |
тоталь ная |
|
|
Крупные |
23,1 |
38,5 |
23,1 |
15,4 |
0 |
|
Средние |
5,56 |
66,7 |
27,8 |
0 |
0 |
|
Мелкие |
2,7 |
35,7 |
28,6 |
7,1 |
25,9 |
Так, среди мелких нейронов максимальное количество клеток в состоянии полного насыщения липофусцином достигает 28,6%, что свидетельствует об их низкой резистентности к липофусциноге-незу. В нейронах крупных и средних размеров процесс липофусцинообразования не достигает подобного распространения и завершается конденсацией пигмента в виде отдельных очагов на одном из полюсов цитоплазмы (средние нейроны) или же стадией двухполюсного (биполярного) расположения липофусцина в клетке (крупные и мелкие нейроны, см. табл. 3).
Авторы, исследовавшие липофусциногенез в нервных клетках других чувствительных узлов, приходят к выводу, что в цитоплазме мелких чувствительных нейронов липофусцин встречается чаще, чем в средних и крупных нейроцитах [9]. В то же время, попыток объяснения причин таких различий в литературе встретить не удалось. По нашему мнению, это связано с отсутствием единой теории, объясняющей механизм образования липофусцина в постмитотических клетках, и, в частности, в нейронах.
Мы склонны предположить, что в активно функционирующих крупных и средних нейроцитах, имеющих близкие показатели ядерно-цитоплазматического отношения и обладающие большей площадью цитоплазмы по сравнению с ядром, существует более значимый ресурс для выведения пигмента из клетки, и, как следствие, предотвращение развития терминальной стадии тотального липофусциногенеза.
Мы сходимся во мнении с авторами, считающими, что мелкие нейроны, у которых площадь цитоплазмы существенно уступает таковым в крупных и средних нервных клетках, обладают пониженной способностью выводить метаболиты (гранулы пигмента) по сравнению с крупными [14].
Учитывая тот факт, что липофусцин накапливается в ущерб синтетической функции клеток, появление в нервно-клеточной популяции узла стареющих и старых нейронов следует признать неблагоприятным признаком, после которого может развиваться терминальная стадия липофусциноге-неза – липофусциноз.
Многочисленные исследования показывают неоднозначность мнений авторов о природе липофусцина и его функционального предназначения в клетках тканей организма (от компенсаторноприспособительного продукта до «клеточного мусора»).
Анализируя материалы наших исследований о содержании липофусцина в нейронах ТУ у лиц в возрасте 60–65 лет, мы сопоставили их с данными, полученными при исследовании этого пигмента в нейронах других чувствительных, в частности, поясничных спинномозговых узлов людей преклонного возраста [9]. Авторы отметили, что липофусцин является характерной особенность нервных клеток спинномозговых узлов в пожилом возрасте.
Вероятно, появление и накопление липофусцина может происходить при естественных процессах старения.
Появление липофусцина в нейронах следует считать его надежным биомаркером старения этих клеток и организма в целом.
Наряду с пигментной дистрофией дегенеративно изменённые нейроны по типу вакуольной дистрофии, выраженной дисхромии, пикноза и нейро-нофагии, были отмечены нами в 21,5% случаев. Вероятно, у людей старше 60 лет возникает возрастная гипоксия смешанного генеза, она является универсальным стимулятором не только липофус-циногенеза, но также беспигментной программированной гибели нервных клеток.
Приступая к обсуждению материалов собственных исследований, необходимо отметить особенности топографии тройничного узла. Узел находится в Меккелевой полости, являющейся для него вместилищем и защитой, он покрыт твердой и паутинной оболочками. Стенки Меккелевой полости оказывают на тройничный узел формообразующее воздействие. Форма и размеры узла зависят также от индивидуальных особенностей черепа [6, 9]. Нахождение в непосредственной близости к узлу анатомических образований (артерий, венозных синусов и твердой мозговой оболочки) подчерки- вает необходимость соблюдения мер предосторожности при выполнении нейрохирургических операций как на самом тройничном узле по поводу прозопалгий (лицевых болей), так и в окружающих тройничный узел узких пространствах, из-за опасности повреждения находящихся рядом важнейших сосудистых образований (внутренней сонной и средней менингеальной артерий, пещеристого, а также заднего межпещеристого синусов).
При рассмотрении строения узла с морфофункциональных позиций, нам представилось целесообразным выделить (конечно, без строгих границ) в тройничном узле как едином анатомическом образовании отдельные части, каждая из которых имеет свои структурные и функциональные особенности. Отличия между зонами заключаются в значениях показателей относительного содержания нейронов, распределением последних в зависимости от размеров их тел, и выраженностью в них липофус-циногенеза.
При этом в глазной зоне преобладают мелкие нейроны (более 50%), обнаружившие низкую резистентность к липофусциногенезу. Более высокая устойчивость к накоплению пигмента была установлена в телах нейронов, заполняющих две другие зоны. Несмотря на то, что по существующим представлениям, часть крупных нейронов с полярным и биполярным распределением липофусцина следует отнести к стареющим и старым, большинство из них сохраняет обычную структуру ядра и значительную часть цитоплазмы, не занятую липофусцином. Это даёт основания полагать о функциональной способности этих клеток. Данный факт является доказательством зависимости устойчивости нейронов к липофусциногенезу от их размеров, а также подчёркивает различия в функциональных возможностях и резервах нервных клеток описанных зон.
Наряду с прогрессирующим липофусциногене-зом в тройничном узле существует другая стратегия дегенеративных возрастных изменений – программированной гибели нейронов, в которых отсутствуют признаки наличия в их цитоплазме данного пигмента или же он находится в начальной стадии своего формирования (диффузная форма).
Такого рода изменения представлены дисхро-мией, гидропией цитоплазмы в виде обильной вакуолизации, сморщивания и, наконец, нейронофа-гией – финальной стадии онтогенеза клетки – её гибелью.
Рассматривая с позиций теории функциональных систем П.К. Анохина обе стратегии гибели нейронов тройничного узла (липофусциногенез и программированную – без липофусцина в клетке), следует считать, что системообразующим фактором, объединяющим эти процессы, является возраст, а непосредственным пусковым механизмом – возрастная гипоксия смешанного генеза, ведущая к повреждению внутриклеточных структур.
Выводы
Тройничный (Гассеров) узел является сложным нервным образованием, обеспечивающим приём, распределение и передачу нервных импульсов к нервным центрам тройничного нерва, расположенным в стволе головного мозга и спинном мозге. Паренхима ганглия представлена нейронами крупных, средних и малых размеров, окружённых клетками перинейрональной глии, а также нервными волокнами, суммарная площадь которых в 4 раза превышает аналогичный показатель тел нейроцитов.
В тройничном узле выделены три основные части, характеризующиеся своими функциональными и морфологическими особенностями:
-
- периферическая часть, содержащая три скопления нейронов, ориентированных в направлении отходящих от узла глазного, верхнечелюстного и нижнечелюстного нервов (первой, второй и третьей ветвей ТУ), разделённых между собой соединительной тканью и проходящими волокнами (сенсорная зона);
-
- промежуточная часть ТУ, расположенная между сенсорной зоной и участком интраганглионар-ного формирования чувствительного корешка V пары черепных нервов, является сенсорнораспределительной зоной;
-
- начальная часть чувствительного корешка тройничного нерва, расположенная интраганглио-нарно и состоящая из центральных отростков псевдоуниполярных нейронов.
