Апробация методов оценки популяционного разнообразия в естественных насаждениях сосны обыкновенной

Автор: Царев Анатолий Петрович
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Сельскохозяйственные науки
Статья в выпуске: 2 (92), 2008 года.
Бесплатный доступ
Исследованы методы К. Шеннона и Л. А. Животовского для оценки популяционного разнообразия сосны обыкновенной в черничниковом типе леса средней тайги. Показано, что эти методы можно использовать для оценки качественных и количественных признаков. Однако в последнем случае требуется соблюдение одинаковой градации признаков при их разделении на классы.
Биоразнообразие, сосна обыкновенная, методы оценки, внутривидовое разнообразие
Короткий адрес: https://sciup.org/14749406
IDR: 14749406 | УДК: 630.18
Текст научной статьи Апробация методов оценки популяционного разнообразия в естественных насаждениях сосны обыкновенной
При генетико-селекционных мероприятиях, проводимых в популяциях, существует опасность снижения уровня генетического разнообразия. Поэтому необходим постоянный или периодический его мониторинг. Для оценки разнообразия необходимы объективные и сравнительно простые методы. Разработка таких методов – сложная и длительная задача, особенно для варьирующих количественных признаков.
Изменчивость количественных признаков является, как правило, непрерывной, что затрудняет дифференциацию совокупностей на классы. В большинстве случаев классификации являются или субъективными, или «по договоренности». Недостатки таких классификаций особенно четко проявились при исследовании внутривидового разнообразия живых организмов. Довольно устоявшимися являются принципы классификации экосистем и надвидовых таксонов, равно как и внутривидовых форм, различающихся по качественным признакам [1, 2, 3]. Однако классификация внутривидовых таксонов по количественным признакам является довольно спорной проблемой.
Для более объективной оценки внутривидового разнообразия, и в частности внутрипопу-ляционного, в генетике предложен ряд методов, в том числе и по количественным признакам [4, 5, 6 и др.]. В некоторых исследованиях эти подходы было предложено использовать для оценки внутрипопуляционного разнообразия лесных древесных пород [7, 8, 9]. Первые опыты по апробации этих методов в Карелии были освещены ранее [3].
Среди трех направлений сохранения биоразнообразия (внутривидового, межвидового и экосистемного), отмеченных в Международной конвенции по сохранению биологического разнообразия [10], наиболее существенное значение в эволюционном и биологическом плане имеет внутривидовое разнообразие. Оно же является и наиболее сложным для исследования.
Так, если нарушение экосистемы или видового состава обнаруживается сравнительно просто, то потеря какой-либо части внутривидового разнообразия не всегда столь очевидна. Например, если в какой-то части ареала сосны обык-
новенной систематически вырубать лучшие деревья, то можно прийти к такому положению, что сосна как вид будет существовать (и в этом смысле с видовым рангом будет все в порядке), но вид этот будет уже, в сущности, другим, обедненным. И при том за счет лучшей части генофонда. Так исчезают мачтовые сосны, не пораженные сердцевинной гнилью осины, карельская береза и другие внутривидовые комплексы. На подобные последствия указывали многие исследователи [11, 12 и др.].
Поэтому очень важно, особенно при селекции, сохранить внутривидовое разнообразие. Этой проблемой в последние годы активно занималась шведская школа лесных генетиков под руководством Дага Линдгрена.
Между тем оценка разнообразия представляет для лесных древесных растений значительные трудности. На это неоднократно обращалось внимание в печати. К настоящему времени предлагаются три подхода для оценки генетического разнообразия на индивидуальном и популяционном уровнях:
-
1. Применение биохимических методов для оценки аллельной вариации [13, 14].
-
2. Использование количественного анализа изменчивости мерных признаков [1, 5, 15, и др.].
-
3. Установление эффективного размера популяции, основанного на родстве генов, индивидуумов и групп [16, 17].
Есть иные направления и разные вариации указанных выше подходов [18, 19, 9, 20, 21, 22 и др.].
Принимая во внимание трудоемкость работы, необходимость получения репрезентативных и достоверных данных, а также обеспечения повторяемости результатов, очень трудно однозначно ответить на вопрос, как же оценивать разнообразие лесных древесных пород, особенно внутривидовое, на уровне популяций. Вероятно, на современном этапе целесообразно было бы использовать фенетические подходы, разработанные отечественными исследователями для изучения популяций живых организмов [1, 23, 24].
Изучение варьирования фенотипических признаков в сочетании с их математико-статистической оценкой для изучения разнообразия популяций, в том числе и для лесных древесных пород, предлагалось также рядом отечественных и зарубежных исследователей [7, 8, 9, 25, 26, 27, 28, 29, 30, 31 и др.].
Учитывая накопленный опыт, все изучаемые признаки лесных древесных пород целесообразно было бы разделять на следующие группы:
-
1. Морфологические , включая признаки, оцениваемые при лесоустроительных работах:
-
• общая высота, диаметр на высоте 1,3 м, высота до первого сучка, протяженность бес-сучковой части ствола, бонитет, объем среднего дерева, полнота, запас, товарность и др.;
-
• в эту же группу относятся ширина, плотность и архитектоника крон; строение, окраска и толщина коры; угол прикрепления сучьев
-
2. Фенологические : распускание почек, облиствение, окончание роста побегов, осенняя окраска листьев, листопад, продолжительность периода роста, вегетации, цветение (пыление), созревание плодов и семян, раскрытие (рассыпание) шишек, опадение семян и др.
-
3. Специфически биологические : плотность древесины, таннидность, смолистость, содержание салицина и др.
-
4. Экологические: светолюбие и теневыносливость, влаголюбие и засухоустойчивость, теплолюбие и зимостойкость, ветро- и газо-устойчивость, отношение к биотическим факторам среды и др.
и их толщина; очищаемость ствола от сучьев; параметры и цвет листьев; окраска цветков и стробилов; характеристика плодов и шишек (форма, размер, масса, окраска, форма апофизов, диссиметрия и др.); характеристика семян (размер, масса, цвет, всхожесть и др.).
Для обработки указанных и других интересующих исследователя и производство признаков с целью изучения их варьирования, оценки внутри- и межпопуляционного внутривидового разнообразия могут быть использованы некоторые методы математического статистического анализа. Например, индекс Шеннона – Н , показатель внутрипопуляционного разнообразия – µ и доля редких морф – h µ Л. А. Животовского, обобщенное расстояние Махаланобиса – D2 , индекс Симпсона – D , мера разнообразия генов – GD Дага Линдгрена и др. (табл. 1).
Очевидно, что подобные подходы, разработанные в популяционной генетике, таксономии и биогеоценологии следовало бы попытаться использовать и при исследовании биоразнообразия лесных древесных пород. Но, естественно, первоначально они должны быть апробированы на применимость их в этом направлении.
В настоящей работе представлены результаты исследований по использованию первых из указанных в табл. 1 методов (К. Шеннона и Л. А. Животовского) для оценки внутривидового разнообразия сосны обыкновенной.
ХАРАКТЕРИСТИКА ОБЪЕКТОВ И ОБЩИХ ТЕОРЕТИЧЕСКИХ ПОДХОДОВ
Оценка внутрипопуляционного разнообразия проводилась в естественных лесных древостоях Pinus sylvestris L. в зоне средней тайги Карелии. Фактические данные собраны в сосняках черничниковых Pinetum myrtillosum Петрозаводского лесхоза. Для примера взяты две пробные площади (№ 1 – в б. Заозерском лесничестве и № 2 – в б. Шуйском лесничестве) размером по 0,25 га.
Общее количество деревьев сосны на пробной площади № 1 – 174, а на пробной площади № 2 – 157 экземпляров. Их средний возраст – 85 и 81 год соответственно. Для предварительных оценок рандомизированным способом отобрано
Таблица 1
Методы изучения внутривидового и внутрипопуляционного разнообразия лесных древесных пород
|
Наименование метода |
Формулы и пояснения |
Источник |
|
Индекс Шеннона – Н |
H =— T p *ln p , i = 1 где s – число видов, pi – доля от общего образца i-го вида |
32, 33, 5, 7 |
|
Показатель внутрипопуляци-онного разнообразия - ц |
m Ц = ( S V p ")2, i = 1 где p i– частота морф, m – количество обнаруженных морф |
6 |
|
Доля редких морф - h ц |
h ц= 1 - ( ц / m ) |
6 |
|
Обобщенное расстояние Махаланобиса – D2 |
p D 2 = S Q, ( Z k..- z» )( Z k.,- Z i,. ), i , j = 1 где Qi j – соответствующий элемент матрицы, обратной к корреляционной матрице; Zk,i; Zl,i– средние нормированные значения i-го признака, соответственно для k -й и l -й совокупностей; Zk, j ; Zl, j – то же для j-го признака; p – число учтенных признаков |
34 |
|
Индекс Симпсона – D |
^L » ( n i — 1) D = 1 - ^=1---------, N ( N - 1) где s – число видов, ni – число индивидуумов в i-м виде, N – общее число индивидуумов в сообществе |
5,7 |
|
Мера разнообразия генов – GD |
GD = 1 - 0 , где Θ – групповое коанцестри (мера родства генов) |
16,17 |
36 и 37 модельных деревьев разной величины. На пробных площадях были проведены измерения и наблюдения по комплексу фенотипических признаков (высота деревьев, диаметр на высоте 1,3 м, высота прикрепления первых живого и мертвого сучьев, толщина и цвет коры, высота подъема грубой корки); определен возраст каждого дерева с помощью бурава, класс роста по Крафту, товарность, диаметр, форма и густота кроны, прирост в высоту, толщина ветвей и степень зарастания сучьев, прямизна ствола и его полнодревесность, тип сексуализации, санитарное состояние и др.
Для оценки внутрипопуляционного и межпопуляционного разнообразия использованы следующие качественные и количественные признаки (фены):
-
• толщина ветвей (толстые, средние, тонкие);
-
• сексуализация (преимущественно мужские стробилы, преимущественно женские, примерно равное соотношение мужских и женских стробилов на дереве, деревья без стробил с невыраженной сексуализацией);
-
• относительная протяженность кроны (% от высоты ствола);
-
• относительная высота (высота/диаметр в одинаковых единицах) по С. Я. Медведеву [35].
Показатель К. Шеннона, или информационная мера разнообразия анализируемого признака в популяции ( H ), для биологических объектов [5] определяется по формуле (табл. 1):
n
H = — ^ P i ln P i , (1)
i = 1
где p i – относительные частоты этого признака; ln – натуральный логарифм.
Для определения достоверности найденной информационной меры Н рассчитывается ее ошибка s H по следующей формуле:
sH
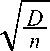
n где D = S Pi (ln Pi)2 — H ." (2а)
i = 1
Различия между двумя популяциями по показателю К. Шеннона определяются по критерию Стьюдента ( t ):
t =
H 1 — H 2
V s H 1 + s H 2
Показатель Шеннона изменяется обратно пропорционально показателю доминирования, то есть дает представление, насколько та или иная совокупность разнообразна. Чем выше показатель Н , тем выше соответственно и разнообразие.
Показатель внутрипопуляционного разнообразия , или среднее число фенотипов ( µ ) , по Л. А. Животовскому определялось по формуле (табл. 1):
m
µ = ( ∑ p i ) , (4)
i = 1
где p i – частота морф, m – количество обнаруженных морф.
При этом под термином «морфа» Л. А. Жи-вотовским [6] понимается «фенотип, генотип, аллель, градация количественного признака (выделено мной. – А. Ц .) и пр.». При равных частотах всех фенотипов µ = m ; при неравномерном распределении частот фенотипов µ < m ; при мономорфизме µ = 1.
Стандартная ошибка µ :
s µ = µ ( m - µ )/ N . (5)
Одновременно с показателем µ для популяций Л. А. Животовский предлагает еще один показатель, названный им «доля редких морф» – h µ :
h µ = 1 - ( µ / m ) . (6)
При этом он отмечает, что если распределение частот морф равномерное, то h µ = 0. При неравномерном распределении частот всегда h µ > 0.
Показатели µ и h µ как бы дополняют друг друга при характеристике популяций. Показатель h µ , по мнению его автора, дает новую по сравнению с µ информацию о характере разнообразия фенотипического состава популяции. В то время как µ оценивает степень разнообразия, показатель h µ дает определенную характеристику структуры этого разнообразия в смысле соотношения между частотами наиболее редких и наиболее частых в этой выборке фенотипов.
Стандартная ошибка доли редких морф:
s h = s µ / m , (7)
или s h = V h µ (1 - h µ )/ N . (8)
Л. А. Животовский [6] отмечает еще одну особенность применения показателей µ и h µ . Они, по его оценке, очень чувствительны к наличию в выборках единичных особей с редким признаком. Их оценки в этом случае становятся стабильными лишь при очень больших объемах выборки. В связи с этим он рекомендует при наличии редких фенотипов объединять их в одну группу, чтобы каждая морфа была представлена как минимум у 3–5 особей.
Оценка значимости различий между популяциями по среднему числу морф и доле редких морф может производиться на основе критерия Стьюдента по формуле (3), в которой вместо значений H 1 и H 2 подставляются соответственно значения µ 1 и µ 2 и h µ 1 и h µ 2 .
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Результаты расчетов показателей внутрипо-пуляционного разнообразия К. Шеннона ( H ) и Л. Животовского ( µ и h ) для разных признаков приведены в таблицах 2–3.
Таблица 2
Внутрипопуляционное разнообразие в сосняках-черничниках Петрозаводского лесхоза (по К. Шеннону)
|
Признаки |
Б.Заозерское лесничество |
Б. Шуйское лесничество |
Коэффициент достоверности различий |
|
H ± S H |
H ± S H |
t |
|
|
Высота |
1,88 ± 0,30 |
1,82 ± 0,01 |
0,20 |
|
Диаметр |
1,80 ± 0,09 |
1,94 ± 0,08 |
1,17 |
|
Относительная |
1,51 ± 0,11 |
1,37 ± 0,10 |
0,93 |
|
высота (H/D) |
|||
|
Толщина ветвей |
0,70 ± 0,13 |
0,55 ± 0,08 |
0,98 |
|
Тип сексуализации |
1,27 ± 0,08 |
0,99 ± 0,14 |
1,75 |
Таблица 3
Внутрипопуляционное разнообразие и доля редких форм в сосняках-черничниках Петрозаводского лесхоза (по Л. А. Животовскому)
|
Признаки |
Б.Заозерское лесничество |
Б. Шуйское лесничество |
Коэффициент достоверности различий |
|
Показатели внутрипопуляционного разнообразия |
|||
|
µ ± S µ |
µ ± S µ |
t |
|
|
Высота |
7,17 ± 0,41 |
6,52 ± 0,29 |
0,46 |
|
Диаметр |
6,51 ± 0,29 |
7,41 ± 0,34 |
1,98 |
|
Относительная |
5,11 ± 0,36 |
4,39 ± 0,27 |
0,35 |
|
высота (H/D) Толщина ветвей |
2,40 ± 0,20 |
1,85 ± 0,09 |
2,56 |
|
Тип сексуализации |
3,76 ± 0,16 |
3,24 ± 0,26 |
1,73 |
|
Доля редких форм |
h ± Sh |
h ± Sh |
t |
|
Высота |
0,10 ± 0,05 |
0,070 ± 0,04 |
0,05 |
|
Диаметр |
0,07 ± 0,042 |
0,074 ± 0,043 |
0,06 |
|
Относительная |
0,15 ± 0,06 |
0,12 ± 0,08 |
0,71 |
|
высота (H/D) Толщина ветвей |
0,20 ± 0,07 |
0,08 ± 0,04 |
1,45 |
|
Тип сексуализации |
0,06 ± 0,04 |
0,19 ± 0,07 |
1,73 |
При оценке показателей все данные группировались по определенным классам. Так, для высоты все данные наблюдений разбивались по 2-метровым классам. Диаметры группировались в 4-сантиметровые ступени толщины. Относительные высоты (H/D, по С. Я. Медведеву) разбивались на классы величиной в 20 единиц. Признак толщины ветвей, как уже было отмечено выше, разбивался на три класса (толстые, средние, тонкие), а тип сексуализации – на 4 (1 – преимущественно мужские стробилы; 2 – преимущественно женские стробилы; 3 – смешанный; 4 – стробилы отсутствуют).
Из данных таблиц 2–3 видно, что показатели для насаждений сосны, произрастающих в разных пунктах одного и того же типа леса, несмотря на некоторое варьирование, оказались довольно близкими.
Сравнение показателей одних и тех же признаков в разных популяциях по критерию Стьюдента ( t ) для показателя К. Шеннона во всех случаях, а для показателей Л. А. Животовского (внутрипопуляционного разнообразия и доли редких форм) в большинстве случаев достоверных различий не выявило.
Изучение внутривидового разнообразия является одним из важных направлений изучения биоразнообразия живой природы и одним из самых сложных. Возможно, поэтому во многих работах последнего времени, посвященных изучению, мониторингу и сохранению биоразнообразия, эти вопросы обходятся молчанием или затрагиваются в незначительной степени [36, 37, 38 и др.].
Представленные в настоящем исследовании результаты показывают, что, несмотря на различие признаков и варьирование их абсолютных значений, по используемым критериям получены близкие результаты для различных насаждений близких типов леса.
Это позволяет надеяться на применимость этих критериев для оценки внутривидового разнообразия лесных древесных пород по разным качественным и количественным признакам. Данные результаты целесообразно было бы проверить на более обширном материале в разных условиях произрастания .
Кроме того, при апробации этих методов в различных вариантах оценок было установлено, что на абсолютную величину как критерия К. Шеннона, так и критериев Л. А. Животовско-го влияет степень дробности градации признака при разделении его значений на классы.
Так, для признака относительной протяженности кроны сосны обыкновенной на пробной площади при градации в 10 % (10–19, 20–29, ..., 50-59) критерий К. Шеннона Н =1,28 ± 0,11. При градации этого же признака по 5-процентным классам на той же пробной площади критерий К. Шеннона оказался равным 1,90 ± 0,09. То есть показатель Н увеличился в 1,5 раза. Такого же порядка увеличение получено и при других вариантах сравнения (табл. 4).
Таблица 4
Изменение величины критерия Шеннона ( Н ) в зависимости от дробности града-ци и количественных признаков (сосна обыкновенная)
|
№ |
Размеры классов (числитель) и их число (знаменатель) при степени градации признака |
Н ± sН при градации признака |
Величина превышения при более дробной градации признака (5/4) |
||
|
менее дробной |
более дробной |
менее дробной |
более дробной |
||
|
1 |
2 |
3 |
4 |
5 |
6 |
Относительная, протяженность кроны, %
1 2
10/5 5/8 1,28 ± 0,11 1,90 ± 0,09
1,48
1,51
10/5 5/8 1,11 ± 0,09 1,68 ± 0,13
Относительная высота (H/D) по Я. С. Медведеву
1 2
20,0/6 10,0/10 1,51 ± 0,11 2,06 ± 0,10
20,0/5 10,0/9 1,37 ± 0,10 2,01 ± 0,09
1,36
1,47
Во всех случаях при переходе к более дробным градациям величина критерия К. Шеннона увеличивалась пропорционально увеличению числа классов. Отсюда следует вывод, что для непрерывно варьирующих количественных признаков его применение не всегда корректно и должно обусловливаться рядом ограничений. В частности, указанный критерий возможно использовать для сравнительных целей по определенным признакам и с обязательной одинаковой градацией признака. То же относится и к балльным шкалам (с числом градаций 3, 4, 5 и больше баллов) количественных и качественных признаков, с учетом того, что при таких наблюдениях и классификациях имеет место определенный субъективизм наблюдателя. В приведенных примерах это можно отнести к фену «толщина ветвей».
Таким образом, исчисленные величины критерия К. Шеннона на основании количественных признаков не являются всеобъемлющими и универсальными. Однако они могут давать относительное представление о внутрипопуляционном разнообразии сравниваемых популяций.
Другими словами, этот критерий может быть использован при четко проявляющихся фенотипических различиях (пирамидальность или рас-кидистость кроны, раннее и позднее распускание листьев, цвет генеративных органов и т. п.), что подтверждается вышеприведенными данными. Если же с его помощью сравниваются количественные признаки, то необходимо соблюдение идентичности подходов к оценке сравниваемых показателей.
Критерии Л. А. Животовского, как и критерий К. Шеннона, также могут использоваться для сравнительных целей в случае одинаковой градации признаков. Для установления этого факта, как и при оценке критерия К. Шеннона, нами были проранжированы значения количественных признаков протяженности кроны и относительной высоты Я. С. Медведева (H/D) при разных степенях их градации. Результаты приведены в таблице 5.
Таблица 5
Изменение показателей внутрипопуля-ционного разнообразия (µ) и доли ред ких форм (h) Л . А . Животовского в зависимости от дробности градации количественных признаков (сосна обыкновенная)
|
№ |
Размеры классов (числитель) и их число (знаменатель) при степени градации признака |
р± S p и h± S h при градации признака |
Величина превышения при более дробной градации признака (5/4) |
||
|
менее дробной |
более дробной |
менее дробной |
более дробной |
||
|
1 |
2 |
3 |
4 |
5 |
6 |
Показатель внутрипопуляционного разнообразия ( р )
|
Отно 1 2 Отно 1 2 Показ |
сительная 10/5 10/5 сительная 20,0/6 20,0/5 атель дол |
протяже 5/8 5/8 высота р 10,0/10 10,0/9 и редких |
нность кр 4,12 ± 0,32 3,75 ± 0,36 H/D) по Я. 5,11 ± 0,36 4,39 ± 0,27 морф (h) |
оны, % 7,26 ± 0,39 6,40 ± 0,53 С. Медведе 8,76 ± 0,55 8,16 ± 0,43 |
1,76 1,71 ву 1,71 1,86 |
|
Отно 1 2 Отно 1 2 |
сительная 10/5 10/5 сительная 20,0/6 20,0/5 |
протяже 5/8 5/8 высота р 10,0/10 10,0/9 |
нность кр 0,18 ± 0,06 0,25 ± 0,07 H/D) по Я. 0,15 ± 0,06 0,12 ± 0,05 |
оны, % 0,09 ± 0,05 0,20 ± 0,07 С. Медведе 0,12 ± 0,05 0,10 ± 0,05 |
0,50 0,80 ву 0,80 0,83 |
Как видно из данных таблицы 5, и при использовании критерия µ Л. А. Животовского наблюдается его увеличение прямо пропорционально степени дробления градаций признака (в 1,7– 1,8 раза). В то же время величина доли редких генотипов ( h) на увеличение степени дробления признака реагирует значительно меньше. Это понятно, так как бывшие редкими генотипы при более дробной градации становятся одними из членов растянувшегося вариационного ряда.
Тем не менее все это говорит о том, что при использовании критериев Л. А. Животовского для сравнительных исследований количественных признаков необходимо также соблюдение равных подходов в отношении их градации, как и в случае применения критерия К. Шеннона.
Получаемые оценки разнообразия имеют определенное экологическое значение. Однако они недостаточно информативны для оценки хозяйственной значимости имеющегося разнообразия. С этой целью необходима модификация данных оценок.
Дальнейшие исследования должны идти в направлении разработки комплексных методов учета биоразнообразия. Эти методы должны будут оценивать как собственно биоразнообразие, так и содержать указания, за счет варьирования каких компонентов исследуемой совокупности оно достигается.
ВЫВОДЫ
-
1. К настоящему времени почти повсеместно осознана необходимость сохранения биоразнообразия живой природы. Но прежде чем принимать какие-либо решения, необходимо оценить состояние биоразнообразия. Для его изучения нужно разработать методы, особенно в части исследования внутривидового разнообразия таких важных для формирования биоты видов, какими являются лесные древесные виды, имеющие огромные ареалы и изменчивость.
-
2. Обычные описательные методы не всегда позволяют оценить внутренние процессы, происходящие в природе, и не всегда могут позволить производить сравнения. Требуется разработка методов, которые были бы относительно доступными, недорогими, объективными и позволяли бы оценивать как отдельные показатели популяции, так и их совокупность.
-
3. Исследователями в разных направлениях науки разработаны некоторые подходы в этом плане, однако они недостаточно широко внедряются в практику лесного хозяйства. Данная работа представляет одну из попыток проанализировать возможности использования количественных методов с целью практического изучения внутривидового разнообразия основных лесообразующих древесных пород.
Апробированные в настоящей работе методы не являются исчерпывающими. Требуются комплексные исследования для изучения использования и других подходов применительно к лесным древесным породам. Но для этого необходимы время, средства и соответствующие специалисты в области изучения, мониторинга, сохранения и восстановления (в случае его нарушения) внутривидового разнообразия лесных древесных пород.
Список литературы Апробация методов оценки популяционного разнообразия в естественных насаждениях сосны обыкновенной
- Яблоков А. В. Популяционная биология. М.: Высшая школа, 1987. 303 с.
- Волков А. Д., Громцев А. Н. Проблема исследования и регулирования биоразнообразия в лесах таежной зоны России. Препринт доклада. Петрозаводск: Ин-т леса КарНЦ РАН, 1997. 24 с.
- Царев А. П. Методы количественной оценки внутривидового разнообразия лесных древесных пород. Петрозаводск: Изд-во Петрозаводского университета, 1998. 52 с.
- Шмальгаузен И. И. Факторы эволюции. 2-е изд. М.: Наука, 1968. 451 с.
- Одум Ю. Основы экологии: пер. с английского. М.: Мир, 1975. 742 с., ил.
- Животовский Л. А. Показатель внутрипопуляционного разнообразия//Журнал общей биологии. 1980. Т. XLI. № 6. С. 828-836.
- Standovar T. Aspects of Diversity in Forest Vegetation//EFI Proceedings. 1996. No 6. P. 17-28.
- Tomppo E. Biodiversity Monitoring in Finnish Forest Inventories//EFI Proceedings. 1996. No 6. P. 87-94.
- Егоров М. Н. Введение в фенетику древесных растений. Воронеж: Изд-во Воронеж. гос. ун-та, 2004. 120 с.
- Convention on Biological Diversity, 1992-last updated 8 September, 1997 [Electronic resource] -Mode access:http://www.biodiv.org/convtext/cbd0000.htm
- Zobel B., Talbert J. Applied Forest Tree Improvement. Printed in the United States of America. New York: John Wiley & Sons, 1984. 505 p.
- Mullin T. J., Bertrand S. Forest management impacts on genetics of forest tree species. Report under Order Number 122994 Ontario Ministry of Natural Resources. Genesis Forest Science Canada Inc. C. P. 64. Quebec. Succursale Haute-Ville. Quebec G 1R4M8. 1998. 36 p.
- Goncharenko G. G., Silin A. E., Padutov V. E. Allozime variation in natural populations of Eurasian pines. III. Population structure, diversity, differentiation and gene flow in central and isolated populations of Pinus sylvestris in Eastern Europe and Siberia//Sylvae Genetica. 1994. V. 43. P. 119-132.
- Падутов В. Е., Баранов О. Ю., Воропаев Е. В. Методы молекулярно-генетического анализа. Минск: Юнипол, 2007. 176 с.
- Драгавцев В. А. Методы анализа внутривидовой изменчивости в лесных популяциях и методы прогноза эффективности аналитической лесной селекции. М.: Центральный НИИ лесной генетики и селекции, 1973. 16 с.
- Lindgren D. Status number for measuring genetic diversity//Forest Genetics, 1997. 4 (2). P. 69-76.
- Lindgren D. Doctorand course OH A2; Kurs 99 A. doc/Dag Lindgren, gen/fys SLU; file:KURSA. DO6. 1999. 54 p. [Electronic resource] -Mode access:http://linne.genfys.slu.se/bred/bred.htm
- Алтухов Ю. П. Генетические процессы в популяциях. М.: ИКЦ «Академкнига», 2003. 429 с., ил.
- Белоконь Ю. С., Белоконь М. М., Голиков А. М., Политов Д. В. Сравнительный анализ генетической структуры сосняков Псковской области//Генетика в XXI веке: современное состояние и перспективы развития. Т. II. III съезд ВОГиС. М. 6-12 июня 2004. С. 192.
- Роне В. М. Генетический анализ лесных популяций. М.: Наука, 1980. 160 с.
- Романовский М. Г. Групповая изменчивость древесных растений по количественным признакам//Фенетика популяций: Сб. науч. тр. М., 1985. С. 88.
- Яхнева Н.В., Ларионова А. Я. Генетическая изменчивость лиственницы Гмелина в природных и индуцированных популяциях//Генетика в XXI веке: современное состояние и перспективы развития. Т. II. III съезд ВОГиС. М., 6-12 июня 2004. С. 190.
- Тимофеев-Ресовский Н. В., Яблоков А. В., Глотов Н. В. Очерки учения о популяции. М.: Наука, 1973. 278 с., ил.
- Яблоков А. В. Фенетика (эволюция, популяция, признак). М.: Наука, 1980. 136 с., ил.
- Сукачев В. Н., Зонн С. В. Методические указания к изучению типов леса. М.: Изд-во АН СССР, 1961. 144 с.
- Правдин Ф. Л. Сосна обыкновенная. Изменчивость, внутривидовая систематика и селекция. М.: Наука, 1964. 190 с.
- Правдин Ф. Л. Ель европейская и ель сибирская в СССР. М.: Наука, 1975. 190 с.
- Ирошников А. И., Мамаев С. А., Правдин Ф. Л., Щербакова М. А. Методика изучения внутривидовой изменчивости древесных пород. М.: ЦНИИЛГиС, 1973. 32 с.
- Животовский Л. А. Популяционная биометрия. М.: Наука, 1991. 271 с.
- Путенихин В. П. Лиственница Сукачева на Южном Урале (изменчивость, популяционная структура и сохранение генофонда). Уфа: Уфимский научный центр РАН, 1993. 195 с.
- Gustafsson L. Biodiversity Assessments in Swedish Forestry Based on Data on Red-Listed Species//EFI Proceedings. 1996. No 6. P. 143-148.
- Шеннон К. Математическая теория связи//Работы по теории информации и кибернетике. М.: Издательство иностранной литературы, 1963. С. 243-322.
- Шмальгаузен И. И. Количество фенотипической информации о строении популяции и скорость естественного отбора//Применение математических методов в биологии. Л.: Изд-во Ленинградского университета, 1960. С. 95-109.
- Перфильев В. Е. Методические рекомендации по применению статистических методов в генетике и селекции плодовых культур. Мичуринск: Центральная генетическая лаборатория, 1980. 34 с.
- Мелехов И. С. Лесоведение. М.: Лесная промышленность, 1980. 406 с., ил.
- Biodiversity and Boreal Forest Ecosystems, Silva Fennica -Special Issue on Climate Change. 1996. Vol. 30(2-3). 383 p.
- Assessment of Biodiversity for Improved Forest Management/EFI proceeding. № 6. 1996. 192 p.
- Assessment of Biodiversity for Improvement Forest Planning/EFI proceeding. № 18. 1998. 420 p.
