Археозоологические контексты к результатам изотопного анализа костей домашних копытных из древнерусских городов

Автор: Антипина Е. Е., Яворская Л. В.
Журнал: Краткие сообщения Института археологии @ksia-iaran
Рубрика: Естественнонаучные методы в археологических исследованиях
Статья в выпуске: 266, 2022 года.
Бесплатный доступ
В статье обсуждаются специфические аспекты средневековой скотоводческой отрасли на территории Восточной Европы, связанные с использованием природных и культивируемых кормовых ресурсов. Представлена новая информация по изотопному составу (13С и 15N) коллагена костей домашних копытных из четырех древнерусских городов, расположенных в разных регионах центральной части лесной зоны. Вариации изотопных данных по домашним копытным рассмотрены на фоне морфологических особенностей животных и археологического контекста памятников. Одновременный анализ этих сведений, вероятно, впервые индивидуализирует изотопную информацию по домашним копытным восточноевропейского средневековья. Дешифровка значений 515N обозначила не только состав диеты и особенности содержания животных, но и некоторые характеристики хозяйственных систем. Показатели 513С по каждому виду в сопряжении с характеристиками природной растительности городской округи позволили предполагать поставку в древнерусские города крупного и мелкого рогатого скота из дальних областей.
Палеоэкономика, средневековые домашние копытные, стабильные изотопы 13с и 15n, палеодиета, древнерусские города
Короткий адрес: https://sciup.org/143179076
IDR: 143179076 | DOI: 10.25681/IARAS.0130-2620.266.265-279
Archaeozoological contexts of the isotopic analysis of domestic ungulate bones from medieval Russia cities
The paper discusses specific aspects of medieval herding sector in Eastern Europe related to the use of natural and cultivated fodder resources. It provides new information on the isotopic composition (13C and 15N) of the collagen from the bones of domestic ungulates from four Medieval Russia cities located in different regions of the central forest belt. Variations in isotopic data on domestic ungulates were examined based on morphological features of animals and archaeological contexts of the sites. The analyses of these data in parallel, maybe, for the first time, individualize isotopic information on domestic ungulates from the East European medieval period. The interpretation of the 515N values helped establish not only dietary habits and specific traits of animal keeping but also identify specific aspects of economic systems. The values of 513С for each species as well as characteristics of natural vegetation in the urban suburb suggest that cattle, goats/sheep were supplied to Medieval Russia cities from far away regions.
Текст научной статьи Археозоологические контексты к результатам изотопного анализа костей домашних копытных из древнерусских городов
Среди главных направлений современной археологии актуальным остается изучение экономического устройства древних обществ, в частности продовольственного сектора, разнообразия задействованных в нем природных ресурсов и способов их освоения. Такие исследования базируются в значительной мере на результатах определения кухонных остатков костей животных. Особая роль
1 Исследование выполнено при финансовой поддержке РФФИ, проект № 20-0900310 «Археозоологические материалы как источник для реконструкции палеоэконо-мических процессов в лесной и степной зонах Восточной Европы в XII–XVI вв.».
отводится информации о размерах домашних и диких копытных, по которой общепринятыми методиками рассчитываются объемы послеубойной или прижизненной продукции. Эти данные позволяют анализировать стратегии жизнеобеспечения в конкретных природных условиях.
Для восточноевропейского средневековья наиболее полное морфологическое описание домашних копытных было сделано еще в середине прошлого века по материалам из древнерусских городов XII–XVI вв. ( Цалкин , 1956). Современные археозоологические обследования сельских и городских поселений Древней Руси подтверждают превалирование в хозяйствах крайне мелких коров и овец, лошадей и свиней ( Антипина, Маслов , 1998; Антипина, Лебедева , 2012; Антипина, Яворская , 2014; Зиновьев , 2015; Антипина , 2019).
Главными природно-хозяйственными факторами, влияющими на размеры животных, являются состав, полноценность и достаточность кормовой базы, основой которой выступают два разных компонента, напрямую связанные со структурой сельскохозяйственного сектора. Это природные растительные корма (луговые и болотные травы, листья и ветки деревьев и кустарников) и культивированные (отходы от производства, переработки и потребления зерновых, технических и огородных культур). Однако морфологические параметры животного, которые обычно регистрируют по остаткам его костей (рост в холке и тип конституции), оказываются недостаточными для разработки подобных вопросов.
Независимым источником информации о кормовом рационе домашних копытных выступает изотопный состав их костей. Многочисленные эксперименты показали, что накопление стабильных изотопов углерода 13С и азота 15N в организме животных связано со спецификой их диеты (наиболее полный список таких публикаций и их анализ см.: Bocherens , 2003; Bocherens, Drucker , 2003). Вместе с тем содержание этих изотопов в почве и растениях определяется разнообразными факторами ( Тиунов , 2007); неоднозначными оказываются и физиологические пути их накопления в тканях копытных ( Drucker et al. , 2008). Несмотря на отмеченные сложности, исследователями все же выявлены устойчивые закономерности обогащения изотопного состава костей млекопитающих в современных наземных экосистемах, что позволяет ориентироваться в результатах изотопного анализа археологических образцов.
В лаборатории естественнонаучных методов ИА РАН создается информационная база по домашним и диким животным разных исторических эпох. Она включает разнообразные сведения, в том числе и морфологические описания конкретных особей, а также их изотопные сигналы. Наибольшие объемы таких данных накоплены по средневековым городам, что позволило нам сопоставить параметры средневековых домашних копытных с изотопным составом их костей в конкретном археозоологическом контексте памятника. Введение в научный оборот полученных результатов этой работы является непосредственной целью данной статьи.
Специфика материала, изотопные данные и методические приемы их расшифровки
Основным материалом для исследования были фрагменты трубчатых костей крупного рогатого скота (КРС), лошадей, овец и свиней из четырех древнерусских городов: Пскова, Ярославля, Переяславля-Рязанского (XIII–XIV вв.) и Ростова (конец XI – перв. пол. XII в.). По этим образцам определены относительный возраст и размеры животных. Среди КРС наиболее многочисленны коровы, по лошадям имеются лишь единичные определения половой принадлежности; а для овец и свиней пол не установлен. Изотопный анализ проведен по методике, общепринятой в международных исследованиях. Ее описание приводится в большинстве отечественных публикаций этого направления, в том числе и с нашим участием ( Яворская и др. , 2015). Изотопные данные по домашним животным из Ярославля и Ростова уже опубликованы (Там же), а по копытным из Пскова и Переяславля-Рязанского представлены впервые2. В качестве сравнительных реперов привлечены результаты изотопного анализа образцов из золотоордынского слоя города Бóлгара (табл. 1).
В совокупности обсуждается 150 образцов костей средневековых домашних копытных с индивидуальными «морфологическими паспортами» и изотопными координатами (сигналы, подписи, метки), при расшифровке которых необходимо учитывать ряд общих сведений. Во-первых, это установленные по историческим и археозоологическим источникам различия в облике разводимых на территории Восточной Европы животных. Древнерусские коровы по высоте в холке были в среднем на 20 см ниже степного скота, хорошо известного по золотоордынским памятникам (рис. 1); по индивидуальным отклонениям разница в росте у них достигала даже 30 см ( Цалкин , 1956; 1967; Петренко , 1984). Наряду с этим размеры лошадей, овец и свиней из разных памятников перекрываются. Во-вторых, хронологические рамки культурных напластований (XIII–XIV вв.), из которых происходят образцы, заставляют обратить внимание на экономические реалии Восточной Европы того времени: перемещения скота между разными социумами в ходе как торговых, так и военных операций. Кроме того, кости домашних копытных в средневековых городах могли происходить и от животных, доставляемых на мясо из сельской округи разной дальности, и от особей, годами содержавшихся непосредственно в городе в качестве молочных и транспортных.
Таблица 1. Изотопные данные δ13С и δ15N в промилле по образцам костей копытных животных из восточноевропейских средневековых городов
|
Псков лес δ 13 C δ 15 N |
Ростов лес δ 13 C δ 15 N |
Ярославль лес δ 13 C δ 15 N |
Переяславль лес δ 13 C δ 15 N |
Болгар лесостепь δ 13 C δ 15 N |
|
|
КРС min max |
5 -22,6 4,4 -22,0 5,7 |
4 -22,3 5,0 -20,1 5,7 |
12 -22,5 6,6 -19,6 9,7 |
8 -22,0 3,7 -19,5 9,4 |
16 -22,0 5,0 -18,0 8,2 |
|
Δ |
0,6 1,3 |
2,2 2,6 |
2,9 3,1 |
2,5 5,7 |
4,0 3,2 |
|
среднее |
-22,4 5,2 |
-21,6 6,4 |
-21,3 8,0 |
-21,1 7,3 |
-20,1 6,8 |
|
Лошадь min max |
1 -22,0 6,0 |
3 -22,0 4,0 -21,7 6,2 |
16 -23,9 5,6 -22,2 7,1 |
2 -22,0 6,0 -21,6 6,8 |
19 -24,4 4,0 -20,6 10,2 |
|
Δ |
0,3 2,2 |
1,7 1,5 |
0,4 0,8 |
3,8 6,2 |
|
|
среднее |
-21,8 4,8 |
-23,1 6,2 |
-21,8 6,4 |
-21,6 6,4 |
|
|
МРС min max |
2 -22,0 4,4 -21,7 6,0 |
3 -21,7 7,6 -21,3 8,4 |
4 -21,4 6,0 -19,8 8,2 |
19 -19,9 5,5 -18,5 9,4 |
|
|
Δ |
0,3 1,1 |
0,4 0,8 |
1,6 2,2 |
1,4 3,9 |
|
|
среднее |
-21,9 5,6 |
-21,5 8,1 |
-20,6 7,3 |
-19,2 7,9 |
|
|
Свинья min max |
3 -21,7 8,5 -21,3 9,7 |
3 -22,2 7,3 -21,2 8,8 |
11 -23,0 6,6 -20,3 12,4 |
10 -22,7 7,2 -19,1 10,4 |
|
|
Δ |
0,4 1,2 |
1,0 1,5 |
2,7 5,8 |
3,6 3,2 |
|
|
среднее |
-21,5 9,2 |
-21,7 7,8 |
-21,9 8,3 |
-21,4 8,4 |
|
|
Лось min max |
2 -23,1 2,5 -22,8 3,8 |
2 -23,4 5,5 |
7 -22,7 4,0 -21,1 7,3 |
||
|
Δ |
0,3 1,3 |
1,6 3,3 |
|||
|
среднее |
-22,9 3,2 |
-21,9 6,1 |
|||
|
Образцы всего |
13 |
12 |
40 |
14 |
71 |

Рис. 1. Пястные кости коров XIV в. из древнерусского Переяславля-Рязанского ( А ) и золотоордынского Бóлгара ( Б )
Методической основой для интерпретации изотопных данных служат широко используемые в международных исследованиях закономерности накопления стабильных изотопов углерода и азота в коллагене костей диких и домашних копытных ( Bocherens , 2003; Bocherens, Drucker , 2003). По этим представлениям, аккумуляция тяжелых изотопов этих двух элементов в тканях животных происходит независимо, но на каждом трофическом уровне фиксируется их обогащение. Так, содержание углерода 13С в организме копытного увеличивается на 2–5 ‰, а азота 15N на 3–4 ‰. Иными словами, в зависимости от видовых особенностей животного именно на такие величины в сторону обогащения должны отличаться изотопные сигналы копытных от изотопных меток поедаемого ими корма.
Растительный фон умеренного климатического пояса Земли составляют так называемые С-3 растения (деревья, кустарники и травы с фотосинтезом через цикл Кальвина). Для них установлены границы по накоплению углерода 13С от -35 до -22 ‰ и азота 15N от -4 до +13 ‰ (Bocherens, 2003). Наиболее низкое содержание тяжелых изотопов у С-3 растений наблюдается для северной флоры, особенно в условиях заболоченности, а самые высокие его оценки типичны для травянистых растений аридных областей, при повышенной солености почв, а также в высокогорьях (Bocherens, 2003; Drucker et al., 2008). Наиболее ярко по указанным факторам различается растительность лесных и степных природных зон. Соответственно, в смешанных и хвойных лесах углеродная изотопная подпись местных диких копытных, как правило, находится в рамках от -30 до -22 ‰, а в степях она превышает -20 ‰ (Bocherens, 2003). Несоответствие углеродных меток домашних копытных изотопным диапазонам диких копытных в конкретном регионе расположения поселения позволяет обсуждать на «пришлый» статус домашних особей (из областей с иными кормовыми ресурсами). Показатели азота (15N) в костях современных диких копытных зависят от конкретного состава корма (травы, листья, ветки, кора в свежем и сухом виде и т. п.). Одним из существенных факторов, изменяющим содержание азота в почвах и растениях, считается антропогенное освоение земельных ресурсов, в том числе культивация сельскохозяйственных угодий. Обнаружено, что зерновые культуры на унавоживаемых землях имеют более высокие сигналы по азоту в сравнении с природными лесолуговыми травами (Bogaard et al., 2007). Этот факт указывает на связь максимальных значений 15N у домашних копытных с наличием в их кормах отходов зерновых. Хотя наравне с этим для копытных остается обоснованной корреляция повышенных оценок этого элемента с потреблением ими сухих и неполноценных, как культивируемых, так и природных, кормов (Choi et al., 2003; Fraser et al., 2011).
Согласно этим закономерностям, для визуализации полученных нами результатов на диаграммах с изотопными данными обозначены эмпирические рамки, разграничивающие домашних копытных с разными диетами. Так, шкала по углероду (δ13С) разделена на две части зеленой вертикальной линией (рис. 2), справа от нее располагаются изотопные сигналы животных с рационом на основе лесной/лесостепной растительности, а слева – с рационом на степных травянистых ресурсах. По шкале азота (δ15N) отмечены границы сигналов копытных первого трофического уровня от 3,0 до 8,0 ‰ (рис. 2: горизонтальные красные линии ), к которому теоретически должны быть отнесены три из обсуждаемых видов: КРС, МРС и лошадь. Для свиньи, как всеядного консумента второго трофического уровня, подразумеваются более высокие показатели. В качестве маркера диеты диких копытных лесной зоны нами принят изотопный состав костей лосей из древнерусского Пскова и Ярославля (табл. 1). Для подтверждения корректности обозначенных выше эмпирических рамок расшифровки изотопных данных привлекаются изотопные координаты домашнего скота из золотоордынского Бóлгара (XIII–XIV вв.). Разводимые на территории лесостепных ландшафтов его округи домашние копытные характеризуются в археозоологи-ческом плане как антиподы мелкорослым древнерусским «лесным» домашним животным ( Петренко , 1984).
Обсуждение
Результаты изотопного анализа представлены в таблице и диаграмме в виде средних арифметических по δ13С и δ15N, максимальных и минимальных их значений, а также индивидуальных изотопных координат (табл. 1; рис. 2). Средние значения по углеродным подписям (δ13С) представителей всех четырех видов (КРС, лошадь, МРС и свинья) из обсуждаемых древнерусских городов оказались
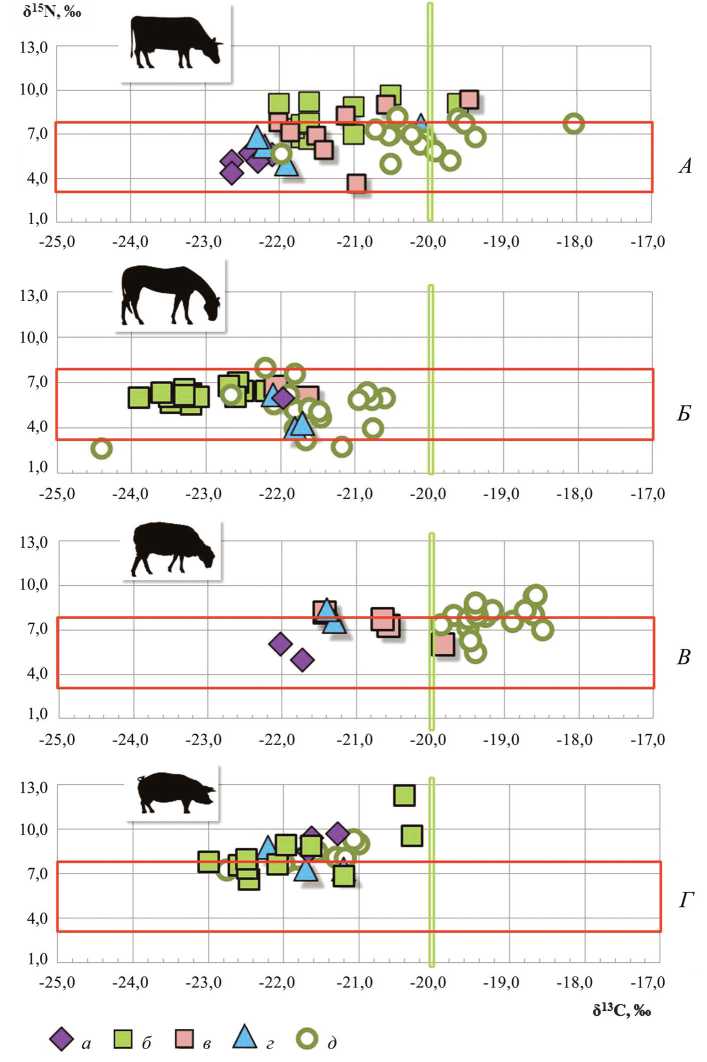
Рис. 2. Индивидуальные изотопные сигналы по углероду и азоту δ13С (‰) и δ15N (‰) домашних копытных из восточноевропейских средневековых городов
А – крупного рогатого скота; Б – лошадей; В – овец; Г – свиней
Условные обозначения: а – Псков; б – Ярославль; в – Переяславль; г – Ростов; д – Бóлгар близки друг к другу с разницей всего в 1,8 ‰ (табл. 1). Эти цифры подтверждают очевидную ситуацию: кормовой рацион животных был основан на С-3 растениях, типичных для флористического покрова центральных областей Восточной Европы. Средние значения по азоту (δ15N) также подтвердили лишь единую трофическую позицию домашних копытных как травоядных животных. Однако индивидуальные изотопные метки конкретных особей очерчивают более сложную и информативную картину: диапазоны максимальных отклонений δ13С (от -23,9 до -19,5 ‰) и δ15N (от 4,8 до 9,2 ‰) обнаруживают значительные различия в составе диеты обсуждаемых видов.
Индивидуальные данные домашних копытных из Пскова и Ростова соответствуют теоретическим представлениям о кормовом рационе травоядных лесной зоны. Сигналы углерода у КРС, лошади, МРС и свиньи (δ13С от -2,6 до -21,2 ‰) маркирует этих животных как местных особей, содержавшихся в ближайшей городской округе (рис. 2: А–Г ). Изотопные метки азота коров, лошадей и овец локализуются в границах первого трофического уровня и фиксируют состав их диеты на основе природных кормов. На таком теоретически ясном фоне исключением стал образец коровы из Ростова с его существенным изотопным обогащением: δ13С = -20,1‰, а δ15N = 7,6 ‰ (рис. 2: А ), что свидетельствует о принадлежности животного к другой, неместной, популяции из более южных, вероятно, лесостепных регионов. Однако эта особь, как и все остальные псковские и ростовские коровы, была типичной представительницей мелкорослого древнерусского скота лесной зоны.
Коровы и лошади из ближайшего к Ростову древнерусского Ярославля имеют совсем иные изотопные подписи, хотя оба памятника расположены в одном природно-ландшафтном регионе. Сигналы углерода и азота у ярославских коров явно выше показателей ростовского скота. Фиксируется также широкая вариабельность индивидуальных значений по обоим элементам: разброс достигает почти 3‰, что указывает на трофическую позицию уже всеядных животных, к которым коровы не могут быть отнесены. Рамки трофического уровня всеядных закономерно занимают изотопные метки свиней, подчеркивая тем самым необычность диеты ярославских коров (рис. 2: А, Г ). При этом изотопные координаты одной из коров (δ13С = -19,6 ‰; δ15N = 9,1 ‰) вообще уходят в диапазон степных копытных, что обозначает ее «пришлый» статус. Но опять же, размеры этой взрослой особи крайне небольшие. В сравнении с ростовскими особями у ярославских лошадей сигналы по углероду, напротив, понижены – даже средняя оценка δ13С опускается у них до -23,1 ‰ (табл. 1), тогда как отметки по шкале азота оказываются существенно выше, хотя и остаются в пределах первого трофического уровня (рис. 2: Б ).
Столь явные несоответствия изотопных данных по домашним копытным из Ростова и Ярославля на фоне значительного сходства природных ландшафтов окрестностей памятников заставляют обратиться к хозяйственным реалиям этих городов. Ростовские (так же, как и псковские) образцы – это остатки костей из обыденного мясного рациона горожан, обеспечение которого происходило из ближайшей сельской округи. Основная часть ярославских образцов – это фрагменты костей животных, содержавшихся непосредственно в городе: их скелеты обнаружены в санитарных захоронениях, связанных с разгромом Ярославля при нашествии Бату-Хана в 1238 г. (Энговатова и др., 2009). Среди них были молочные коровы мелких размеров, небольшие по размерам свиньи, а также малорослые и среднерослые лошади, как верховые, так и гужевые (Антипина, Лебедева, 2012).
Ранее по первым результатам изотопного анализа образцов из Ростова и Ярославля мы уже предполагали преобладание в кормах ростовского КРС природных компонентов, а в кормах ярославских коров – отходов от зерновых культур, обогащенных стабильными изотопами азота ( Яворская и др. , 2015). Эта гипотеза достаточно очевидна, потому что свежие травы природных лугов с относительно низким содержанием азота составляют обычный летний корм сельского скота, но малодоступны городским коровам. И все же для ярославских сельских коров был проведен изотопный анализ дополнительной серии образцов их костей из кухонных остатков от обыденного мясного стола горожан. По этим образцам ожидались изотопные показатели, близкие меткам ростовского скота. Но полученные оценки оказались почти идентичными высоким изотопным сигналам у городских особей; появилась также и указанная выше «степная» корова. Эти результаты заставляют признать, что и в городе, и в сельской округе Ярославля кормовой рацион КРС в значительной мере базировался на отходах зерновых и технических культур.
Такая реконструкция состава корма ярославских коров косвенно подтверждает значительную степень сельскохозяйственной освоенности окрестностей Ярославля в XIII в. и существование системы унавоживания пахотных угодий. Изотопные данные по ростовскому КРС фиксируют иную хозяйственную ситуацию. По-видимому, в конце XI – перв. пол. XII в. природные кормовые ресурсы округи Ростова позволяли регулярное сезонное содержание стада на суходольных и пойменных лугах. Однако оценка экстерьера ярославских и ростовских коров показывает, что базовые различия в составе кормов не отразились на размерах животных: обе популяции относятся к мелкорослому древнерусскому скоту. Небольшие размеры восточноевропейского скота в лесной зоне Восточной Европы обычно связывают с обедненным составом и малой питательностью лесных кормовых ресурсов – трав лесных лугов и болот, а также лиственного фуража ( Цалкин , 1956). Но, как выясняется, культивируемые корма в виде отходов от зерновых культур (мякина и солома) также не отличались полноценностью и удобоваримостью и не способствовали увеличению размеров животных. На низкое качество такого фуража указывают нередко обнаруживаемые на челюстях ярославских городских и сельских коров последствия оссифицирующего периостита и травм, обусловленные потреблением сухих жестких кормов.
Узкие рамки изотопных подписей группы ярославских лошадей из санитарных захоронений – δ13С от -23,9 до -23,1 ‰; δ15N от 6,1 до 6,8 ‰ (рис. 2: Б) – выглядят типичными для диких копытных лесной зоны (данные по лосю) (табл. 1). Вероятнее всего, их кормовой рацион базировался на местных природных ресурсах (лугопастбищная растительность и сено). Одновременно с этим изотопные координаты другой группы ярославских лошадей, останки которых найдены на территории богатой усадьбы – «княжого двора» (Энговатова и др., 2009), показали совсем иной рацион. Диета этих шести лошадей характеризуется изотопным обогащением по обоим элементам: δ13С от -22,7 до -22,1 ‰; δ15N от 6,2 до 7,1 ‰ (рис. 2: Б). Эти факты можно связать с использованием в кормах наряду с сеном добавок полноценного зерна; любопытно, что самый высокий сигнал по азоту (δ15N = 7,1 ‰) оказался у наиболее крупного жеребца ростом 155 см в холке. Учитывая археологический контекст обнаружения останков этих коней, сходство их изотопных меток можно рассматривать как маркер принадлежности всех шести особей к одной конюшне. На фоне таких заключений зафиксированные у ростовских лошадей самые низкие значения по азоту находят объяснение исключительно в круглогодичном их содержании на вольном выпасе. Именно подножный зимний корм (замерзшие сухие травы с сезонно низким азотом), вероятнее всего, был главной причиной столь существенного понижения сигнала этого элемента.
Как уже отмечено выше, изотопные метки свиней из Ярославля достаточно точно соответствуют всеядным копытным второго трофического уровня. Однако у двух особей значения по обоим элементам оказались необычно высокими – δ13С = -20,3 и -20,4 ‰; δ15N = 9,6 и 12,3 ‰ (рис. 2: Г ) – и сходными с показателями диеты собак ( Яворская и др. , 2015). Две гипотезы в равной степени могут объяснить эти отклонения. Первая связана с включением в обычный корм свиней растительных добавок, обогащенных изотопами углерода и особенно азота: испорченного зерна, жмыха технических культур, а также корнеплодов. Вторая гипотеза подчеркивает животный компонент добавок: отходы от разделки туш животных и рыбы. Отмеченные у этих свиней зубные патологии (пародонтоз и оссифицирующий периостит, а также неравномерность и избыточность стирания зубов) вместе с небольшими размерами особей уточняют специфику добавок грубого и неполноценного корма и указывают на преимущественно животные и рыбные отбросы (связки, кости, чешуя).
Результаты изотопного анализа костей коров и лошадей из Переяславля-Ря-занского, несмотря на более южное его расположение по отношению к Ярославлю, обнаруживают значительное сходство с изотопными координатами ярославских животных (табл. 1; рис. 2). Полной аналогией выглядит кормовой рацион малорослых переяславльских коров с повышенным содержанием азота, фиксирующим использование отходов зерновых (мякины и соломы). Однако даже в нем есть два исключения. Первое – это диета, несомненно, местной коровы с поразительно низким сигналом по азоту (δ15N = 3,7 ‰) (рис. 2: А ), что может отражать лишь ситуацию летнего лесного выпаса животного и зимнего его содержания на веточном корме. Второе исключение относится к выявленной, как и в Ярославле, «степной» особи: δ13С = -19,4 ‰ (рис. 2: А ). Но в Переяславле «степная» корова, в отличие от ярославской, характеризуется крупными размерами с высотой в холке около 120 см. Такая конституция вызывает явные параллели с высокорослым степным, в частности – золотоордынским, скотом. Появления степного скота в Переяславле-Рязанском кажется вполне вероятным, ведь земли Рязанского княжества напрямую граничили с Диким полем, а после монгольского нашествия и с территорией Золотой Орды.
Привлекаемый для сравнения изотопный состав образцов домашних копытных из золотоордынского Бóлгара отличается от изотопного статуса древнерусского скота, что соответствует теоретическим представлениям. Это подтверждает корректность выбранных нами методических подходов к расшифровке обсуждаемых данных. Для животных из Бóлгара изотопные координаты маркируют диету на основе природных растительных кормов лесостепного региона с максимальным разнообразием ландшафтов. Особенно ярко это прослеживается по сигналам КРС, большая часть которых образуют компактную «лесостепную» группу. Одновременно по углеродной шкале фиксируется присутствие единичных «лесных» (δ13С = -22,0 ‰) и однозначно «степных» особей (δ13С= -18,0 ‰), хотя для них значения по шкале азота оказываются ниже типичных показателей для ярославских городских коров с диетой на основе отходов зерновых культур (рис. 2: А). Не менее отчетливо выглядит расхождение «степных» золотоордынских овец – по всем образцам δ13С больше -20,0 ‰ (рис. 2: В) – с древнерусскими «лесными» представителями этого вида. Только в Переяславле-Рязанском обнаружена овца, углеродная метка которой (δ13С = -19,8 ‰) близка к сигналам бóлгарских особей, что позволяет предполагать ее степное происхождение. Обнаружение таких степных овец и коров в Бóлгаре подтверждает археозоологиче-скую реконструкцию функционирования городской торгово-производственной отрасли на основе регулярных поставок из степных кочевий многочисленных стад рогатого скота на убой для получения шкур (Яворская, 2021).
Крайне любопытны изотопные координаты лошадей из Бóлгара: большая часть оценок по углеродной шкале происходит от животных, в диете которых доминировали природные корма лесостепной зоны. Вместе с тем по ряду сигналов азота можно предполагать присутствие в Бóлгаре, как и в Ростове, лошадей с круглогодичным вольным выпасом (δ15N = 2,8, 3,2, 4,0 ‰), а также исключительно городских верховых коней с обеспечением их качественным зерновым кормом: δ15N = 7,6 и 8,0 ‰ (рис. 2: Б ). На этом фоне выделяется явно неместная лошадь с особыми, крайне низкими, изотопными метками (δ13С = -24,4; δ15N = 2,7 ‰), которые указывают на северо-западные территории ее круглогодичного содержания на природных подножных кормах.
Заключение
Проведенные изотопные исследования костных остатков позволили нам сделать ряд важных обобщений.
Прежде всего становится очевидным, что наиболее информативной оказалась дешифровка изотопных сигналов углерода и азота (δ13С и δ15N) не по средним их значениям для вида, а по изотопным подписям конкретных особей. Сопоставление результатов изотопного анализа с морфологическим описанием животного дает возможность, вероятно, впервые индивидуализировать изотопные данные по восточноевропейским средневековым домашним копытным. Такой подход раскрывает не только состав диеты и особенности содержания животного, но и указывает на специфические аспекты хозяйственной системы, связанные с использованием природных и/или культивируемых кормовых ресурсов. Так, на основе сигналов азота (δ15N) по образцам из Ярославля выявлено использование отходов от переработки зерновых культур в качестве основного фуража для городских и сельских коров, что отражает господство в округе города земледельческих пахотных угодий и существование системы их унавоживания уже в XIII в.
К сожалению, по изотопным данным оказалось невозможным оценить сбалансированность, удобоваримость и достаточность кормов. Решающая роль в прояснении этого вопроса принадлежит морфологическим характеристикам изучаемых животных. Именно обнаруженные патологии и малорослость древнерусского скота в разных регионах лесной зоны, изотопные сигналы которого зафиксировали разный кормовой рацион, свидетельствуют о неполноценности и низкой удобоваримости фуража и на основе природной растительности лесной зоны, и на отходах от зерновых культур.
Вместе с тем индивидуальные сигналы по углероду могут транслироваться непосредственно на статус нахождения животных на поселении: «местные» или «пришлые». В нашем исследовании удалось среди основного местного стада обнаружить «пришлых» коров в Ярославле, а в Переяславле-Рязанском – коров и овец из степных областей. В Бóлгаре оказались коровы и лошади из северо-западных регионов, а овцы – из дальних степных кочевий. Эта информация подтверждает известные по историческим источникам масштабные разнонаправленные перемещения скота по территории Восточной Европы между древнерусскими княжествами, золотоордынскими улусами, северо-западными и южными государствами в ходе как торговых, так и военных операций.
И наконец, стоит пожелать, чтобы сопряженность рассмотрения изотопного статуса и морфологических характеристик домашних животных, раскрывающая новые аспекты в изучении экономического устройства древних обществ, стала традиционной частью междисциплинарных исследований.
Список литературы Археозоологические контексты к результатам изотопного анализа костей домашних копытных из древнерусских городов
- Антипина Е. Е., 2019. Глава 8. Остеологическая коллекция из Житного раскопа: некоторые аспекты системы жизнеобеспечения Переяславля-Рязанского // Материалы по археологии Переяславля-Рязанского. Вып. 3. Комплексные археологические исследования Переяславля-Рязанского. М.: Таус. С. 240–270.
- Антипина Е. Е., Лебедева Е. Ю., 2012. Глава 6. Растения и животные // Археология древнего Ярославля: загадки и открытия. 2-е изд., доп. и перераб. М.: ИА РАН. С. 144–229.
- Антипина Е. Е., Маслов С. П., 1998. Системы земледелия и размер домашних животных Древней Руси // Труды VI Международного Конгресса славянской археологии. Т. 4. Общество, экономика, культура и искусство славян. / Ред. В. В. Седов. М.: Эдиториал УРСС. С. 52–60.
- Антипина Е. Е., Яворская Л. В., 2014. Реконструкция размеров крупного рогатого скота по остеологическим материалам из средневековых городов и селищ на территории европейской части России // КСИА. Вып. 236. С. 331–338.
- Зиновьев А. В., 2015. Обзор конституционных особенностей крупного рогатого скота и лошадей из средневековых слоев Твери // ТАС. Т. 10. Тверь: Тверской гос. объед. музей. С. 326–330.
- Петренко А. Г., 1984. Древнее и средневековое животноводство Среднего Поволжья и Предуралья. М.: Наука. 174 с.
- Тиунов А. В., 2007. Стабильные изотопы углерода и азота в почвенно-экологических исследованиях // Известия РАН. Серия биологическая. № 4. С. 475–489.
- Цалкин В. И., 1956. Материалы к истории скотоводства и охоты в Древней Руси. М.: АН СССР. 183 с. (МИА; № 51.)
- Цалкин В. И., 1967. Домашние животные Золотой Орды // Бюллетень Московского общества испытателей природы. Отделение биологии. Т. LXXII. № 1. С. 114–130.
- Энговатова А. В., Осипов Д. О., Фараджева Н. Н., Бужилова А. П., Гончарова Н. Н., 2009. Массовые средневековые захоронения в Ярославле: анализ археологических и антропологических материалов // РА. № 2. С. 68–78.
- Яворская Л. В., 2021. Археозоологические исследования городов Золотой Орды: современные интерпретации // Аналитические исследования лаборатории естественнонаучных методов. Вып. 5 / Отв. ред., сост.: Е. Н. Черных, В. И. Завьялов. М.: ИА РАН. С. 216–226.
- Яворская Л. В., Антипина Е. Е., Энговатова А. В., Зайцева Г. И., 2015. Стабильные изотопы углерода и азота в костях домашних животных из трех городов европейской части России: первые результаты и интерпретации // Вестник Волгоградского государственного ун-та. Серия 4: История. Регионоведение. Международные отношени. № 1. С. 54–64.
- Bocherens H., 2003. Isotopic biogeochemistry and the palaeoecology of the mammoth steppe fauna // Advances in Mammoth research / Eds.: W. F. Reumer, J. De Vos, D. Mol. Rotterdam. P. 57–76. (Deinsea; vol. 9.)
- Bocherens H., Drucker D., 2003. Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems // International Journal of Osteoarchaeology. Vol. 13. Iss. 1–2. P. 46–53.
- Bogaard A., Heaton T. H. E., Poulton P., Merbach I., 2007. The impact of manuring on nitrogen isotope ratios in cereals: Archaeological implications for reconstruction of diet and crop management practices // JAS. Vol. 34. Iss. 3. P. 335–343.
- Choi W. J., Ro H. M., Hobbie E. A., 2003. Patterns of natural 15N in soils and plants from chemically and organically fertilized uplands // Soil Biology and Biochemistry. Vol. 35. Iss. 11. P. 1493–1500.
- Drucker D. G., Bridault A., Hobson K. A., Szuma E., Bocherens H., 2008. Can carbon-13 in large herbivores reflect the canopy effect in temperate and boreal ecosystems? Evidence from modern and ancient ungulates // Palaeogeography, Palaeoclimatology, Palaeoecology. Vol. 266. Iss.1–2. P. 69–82.
- Fraser R. A., Bogaard A., Heaton T., Charles M., Jones G., Christensen B. T. et al., 2011. Manuring and stable nitrogen isotope ratios in cereals and pulses: towards a new archaeobotanical approach to the inference of land use and dietary practices // JAS. Vol. 38. Iss. 10. P. 2790–2804.
