Архитектоника растительных пологов цистозировых фитоценозов, Черное море

Автор: Празукин Александр Васильевич, Фирсов Юрий Константинович
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 1, 2016 года.
Бесплатный доступ
Рассмотрена архитектоника растительного полога верхнего яруса цистозировых ассоциаций. Для него характерен одномодальный тип вертикального распределения биомассы с максимумом в его нижней половине, где сосредоточено от 75 до 85% всей сухой массы растений. Распределение биомассы слоевищ разных возрастных групп цистозир по профилю полога описывается серией одновершинных кривых. Виды эпифитных синузий располагаются преимущественно в верхней половине полога.
Фитоценоз, фитобентос, цистозира, эпифиты, биомасса, многоклеточные водоросли, архитектоника растительного полога
Короткий адрес: https://sciup.org/146116621
IDR: 146116621 | УДК: 574.4+574.58
The architectonics of cystoseira phytocenoses canopies in the Black Sea
The article reviews the architectonics of the vegetative canopy as represented in the upper synfolium of cystoseira associations. This canopy is distinguished by a unimodal type of the biomass vertical distribution where 75%-85% of the total dry mass is concentrated at a maximum in its lower parts. The manner in which the cystoseira thallome biomass of various age groups is distributed in the canopy from top to bottom is described by a series of unimodal curves. The epiphyte synusia species are predominantly found in the upper parts of the canopy.
Текст научной статьи Архитектоника растительных пологов цистозировых фитоценозов, Черное море
Введение. Основная роль в создании первичной продукции в прибрежных экосистемах Черного моря принадлежит макрофитобентосу. В настоящее время в литературе рассматривался несколько направлений в его изучении. Большое место в исследованиях принадлежит установлению и уточнению флористического состава фитоценозов и изучению биологии отдельных видов с учетом их значимости в экосистемах и в деятельности человека (Мильчакова, Рябогина, 2002; Костенко и др., 2005; Евстигнеева, 2006). В этих же работах разносторонне обсуждается проблема видового разнообразия и роль заповедных объектов в сохранении редких и исчезающих видов. Ряд публикаций специально посвящено оценке природных запасов тех или иных групп видов водорослей, а также оценке антропогенного воздействия на прибрежные фитоценозы и отдельные группы водорослей (Сабурин, 2004; Костенко и др., 2005; Миронова и др., 2012). Затрагиваются проблемы рационального использования, охраны и воспроизводства растительных ресурсов прибрежных экосистем. Вопросы вертикального и горизонтального распределения водорослей в прибрежных экосистемах и их продуктивности были и остаются актуальными (Калугина-Гутник, 1975; Сабурин, 2004; Симакова, 2011). Имеется подробная характеристика ярусной структуры ассоциаций многоклеточных водорослей черноморских фитоценозов (Калугина-Гутник, 1975).
Несмотря на высокую степень изученности макрофитобентоса Черного моря, вопросы, связанные с архитектурой (вертикальной структурой) растительного покрова как целостной системы, изучены слабо. Необходимость в этих знаниях продиктована двумя важными и взаимосвязанными проблемами: проблемой сохранения естественного состояния прибрежных фитоцензов и проблемой целевого использования цистозир в качестве потенциального сырья в получении фармакологически ценных препаратов. В современной литературе проблема архитектоники растительных пологов обсуждается преимущественно по отношению к наземным растениям и в связи с вопросами в получении высоких и гарантированных урожаев сельскохозяйственных культур (Росс, Росс, 1969; Росс, 1975; Тооминг, 1984; Шульгин, 2013), и, в меньшей степени, по отношению к водным макрофитам. В последнем случае спектр интересов постоянно расширяется и охватывает разносторонние вопросы, связанные с изучением естественных (Завалко, Ковальчук, 1994; Cenzato, Ganf, 2001; Празукин, Фирсов, 2013; Празукин, 2015а, б) и искусственно создаваемых (Празукин, Хайлов, 1998) фитосистем.
В настоящем исследовании мы задались целью провести параметрическое описание архитектоники (вертикальной структуры) растительного полога верхнего яруса цистозировой ассоциации черноморского фитоценоза в мелководных участках прибрежья.
Методика. В местах проведения исследований (рис. 1), на мелководных участках прибрежья района Севастопольской бухты (глубина, Н = 55 - 60 см) с активной гидродинамикой, формируются многоярусные, сомкнутые фитоценозы с хорошо развитыми растительными покровами. На станциях отбора проб 1, 5, 7 и 8 (рис. 1)
Крымский
\ п-ов
Чёрное море /7
г. Севастополь м. Херсонес
-
4) г. Севастополь
Рис. 1 . Расположение станций отбора проб (1 - 8) в прибрежной акватории г. Севастополя. На станциях 1, 5, 7 и 8 растительный полог формируется вертикальными «побегами» С. crinita и её эпифитами, а на станциях 2, 3, 4 и 6 - вертикальными «побегами» двух видов цистозир: (С. crinita, С. barbata^ и их эпифитами.
верхний, господствующий ярус зарослей определяется многолетними слоевищами Cystoseira crinita Вогу, а на станциях 2, 3, 4 и 6 - слоевищами двух видов цистозир (С. crinita, С. barbata (Good, et Wood.) Ag). На цистозирах эпифитируют одноклеточные и многоклеточные водоросли (табл. 1). По классификации донной растительности Черного моря (Калугина-Гутник, 1975) рассматриваемое нами фитосообщество относится к цистозировой ассоциации (Асе. Cystoseira crinita + С. barbata - Cladostephus verticillatus - Corallina mediterranean В наших исследованиях ограничимся только рассмотрением вертикальной структуры растительного полога (РП, в английской транскрипции «vegetative canopy») верхнего яруса цистозировой ассоциации.
Пробы макроводорослей отбирали в июле 2013 г. по методике Калугиной (1969) учетной рамкой 50 х 50 см из серединной части растительного покрова (рис. 2а) на относительно горизонтальных участках дна сложенного твердыми грунтами. Прямые источники загрязнения на рассматриваемых отрезках побережья отсутствовали, однако в летнее время значительно увеличивается рекреационная нагрузка в акваториях станций 2 - б, 8 (Ковардаков и др., 2009) и, в меньшей мере, этот фактор оказывает влияние в районах мысов Херсонес (станция 1) и Толстый (станция 7). Станция 6 располагалась непосредственно в межбунном пространстве городского пляжного комплекса «Парк Победы».
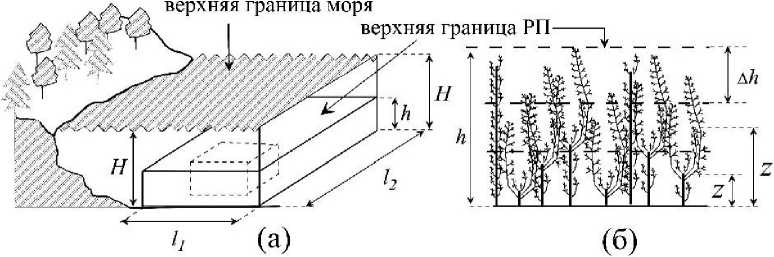
Рис. 2. Участок растительного полога (РП) многоклеточных водорослей Qi,b, 1^ в нижней литоральной зоне (глубина, Н= 55 -60 см) открытого участка побережья (а). РП - разделен на горизонтальные слои с шагом Ah = 5 см. h, /1, /2 - линейные характеристики РП, Z - расстояние от нижней границы РП до середины любого его горизонтального слоя (б).
Для изучения вертикальной структуры растительного полога использовали метод послойного среза (Полевая геоботаника, 1964). Каждое слоевище (вертикальный «побег») цистозир (С. barbata, С. crinita) с их эпифитами рассекали на горизонтальные слои с шагом Ah = 5 см (рис.1 б). Полученные таким образом высечки талломов всех растений одной возрастной группы цистозир (Т, год) послойно объединяли вместе, высушивали и определяли сухую массу (Wcvx(z)). Для каждого слоя полога по каждой группе объектов (стволы, ветви первого порядка слоевищ цистозир Т-го возраста, талломы эпифитов)
рассчитывали сухую массу на единицу поверхности дна (W/So)z = Wcyx(z) /So), где: So - площадь поверхности дна; Z - расстояние от нижней границы РП до середины любого его горизонтального слоя.
Архитектура РП описывалась следующими функциями от Z:
-
1) сухая масса всех водорослей и отдельно каждого вида на единицу горизонтальной поверхности РП на высоте Z - W/So(Z);
-
2) сухая масса структурных элементов слоевищ разного возраста (Т) цистозир на единицу горизонтальной поверхности РП на высоте Z - Wt/S0(Z). '
Результаты. Черноморские цистозиры (С. crinita, С. barbata') - крупные многоклеточные водоросли, виды-эдификаторы, являются основой донных фитоценозов прибрежных экосистем и видами, во многом определяющими архитектуру растительного покрова. Ассоциации многоклеточных водорослей с участием цистозир на твердых грунтах формируют обширные растительные покровы вдоль береговой линии Крымского полуострова и Кавказского побережья до глубин 15 - 20 м (Калугина-Гутник, 1975). Фитоценозы в границах растительного покрова имеют многоярусную структуру. В каждом случае ярус - это совокупность нескольких видов по горизонтальной составляющей с характерными для него биотопическими свойствами, а все виды растений отдельно взятого участка растительного покрова в границах их общего обитаемого пространства, включая в эти границы и минеральную составляющую окружающей их среды, представляют собой иерархически организованную биокосную систему (Празукин, 2015а). ” - - -
Таблица!
Количественные соотношения макрофитов в верхнем ярусе цистозировой ассоциации на разных участках побережья
|
№ станции |
Виды-эдификаторы в верхнем ярусе цистозировой ассоциации |
Сухая масса водорослей, мг см2 |
Доля эпифитов в общей массе водорослей, % |
|||
|
Цистозиры и эпифиты |
С. crinita |
С. barbata |
Эпифиты |
|||
|
8 |
Cystoseira crinita Bory |
135,50 |
128,92 |
- |
6,58 |
4,85 |
|
1 |
100,01 |
98,57 |
- |
1,44 |
1,44 |
|
|
5 |
93,50 |
80,34 |
- |
13,16 |
14,07 |
|
|
7 |
74,44 |
74,08 |
- |
0,36 |
0,48 |
|
|
3 |
Cystoseira crinita Bory, C. barbata (Good, et Wood.) Ag |
160,12 |
45,31 |
114,50 |
0,31 |
0,19 |
|
4 |
98,75 |
85,07 |
6,47 |
7,21 |
7,30 |
|
|
2 |
90,60 |
47,08 |
42,14 |
1,38 |
1,52 |
|
|
6 |
50,55 |
37,19 |
12,06 |
1,31 |
2,59 |
|
В табл. 1 приведены количественные соотношения макрофитов в верхнем ярусе цистозировой ассоциации на разных участках побережья (станции отбора проб). Представленные данные разделены на две группы в соответствии с присутствием в сообществе
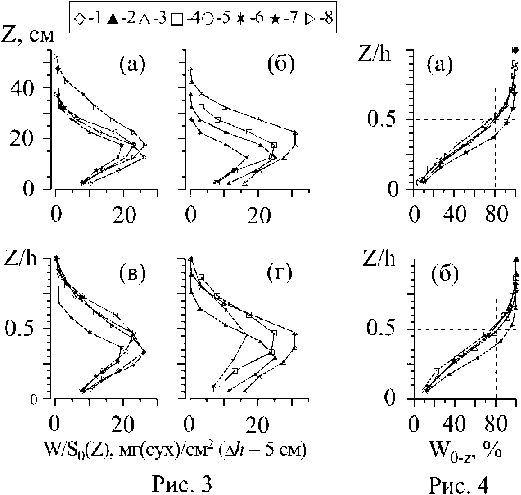
Рис. 3 . Вертикальное распределение сухой массы W/So (Z) в растительном пологе (РП), формируемом вертикальными «побегами» С. crinita и её эпифитами (а, в) и вертикальными побегами двух видов цистозир (С. crinita, С. barbata) и их эпифитами (б, г) на станциях (1 - 8) в прибрежной акватории г. Севастополя (рис. 1). На рис. в, г Z нормировано по отношению к высоте (h) ' РП. '
Рис. 4 . Изменение кумулятивной массы (W0.z, %) по относительной высоте (Z/h) в растительном пологе (РП), формируемом вертикальными «побегами» С. crinita и её эпифитами (а) и вертикальными «побегами» двух видов цистозир (С. crinita, С. barbata) и их эпифитами (б) на станциях (1-8) в прибрежной акватории г. Севастополя (рис. 1).Обозначения те же, что и на рис. 3
одного или двух видов цистозир. В обоих случаях, общая биомасса водорослей на единице поверхности дна (W/So) варьируется примерно в одном диапазоне значений (51 - 160 мг см"2) и средние значения W/So по каждой группе объектов имеют одинаковую величину - 100 мг см"2. Наблюдается невысокий уровень эпифитирования цистозир многоклеточными водорослями. Сухая биомасса эпифитных синузий на разных участках прибрежья составляет от 0,2 до 14 % общей биомассы макрофитов РП. Весовые соотношения между двумя видами цистозир при совместном их произрастании складываются по-разному. Равные соотношения наблюдаются на станции 2, заметное превышение (2,5 раза) в пользу С. barbata - на станции 3, и значительное, в 3 и 13 раз в пользу С. crinita, соответственно, на станциях б и 4.
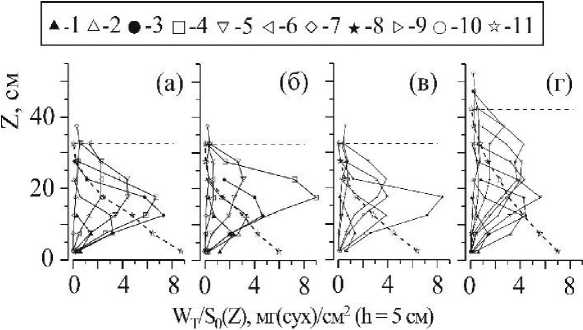
Рис. 5 . Вертикальное распределение суммарной сухой массы WT/SO (Z) ветвей каждой возрастной группы вертикальных «побегов» С. crinita (1 - 10) и стволов растений всех возрастных групп вместе (И) в РП на станциях: 1 (а), 5 (б), 7 (в), 8 (г). Возрастные группы вертикальных «побегов»: 1 - 0,13 < 1,9;
2 - 1,9 < 3,7; 3 - 3,7 < 5,4; 4 -5,4 < 7,2; 5 -7,2 < 8,9; 6 -8,9 < 10,7; 7 -10,7 < 12,5;
8 -12,5 < 17,2; 9 -17,2 < 16; 10 -16 < 17,7 лет
Обратимся к группе данных (рис. За, в), характеризующих архитектонику растительного полога складывающегося вертикальными «побегами» С. crinita и её эпифитами преимущественно открытого прибрежья (рис. 1, станции; 1, 5, 7, 8). В пологе названных видов (рис. За, в) наблюдается неравномерность в распределении W/So(z), причем максимум биомассы во всех рассматриваемых ситуациях располагается в нижней половине полога и в этой части полога сосредоточено около 80% всей фитомассы, включая талломы цистозиры и её эпифиты (рис. 4а). Величины значений максимумов W/So(z) варьируются в относительно узком диапазоне (19,4 - 26,4 мг(сух) см"2, рис. 3 а, в).
Одномодальный тип распределения W/So(z) (рис. За, в) обусловлен характером расположения по высоте РП верхушечных и адвентивных ветвей слоевищ С. crinita разного возраста (Wt/So(z), рис. 5). Распределение сухой массы ветвей со слоевищ соответствующего возраста описываются чаще одновершинными кривыми, и они могут быть как симметричными, так и ассиметричными относительно своего максимума. С увеличением возраста слоевищ максимум биомассы ветвей, как правило, смещается в сторону верхних границ полога. Но уже в возрасте старше 5 лет, а чаще всего, старше 9 лет наблюдается относительно равномерное распределение Wt/Sq(z) и связано это с тем, что в структуре слоевищ в этом возрасте преобладают ветви адвентивного комплекса (Празукин, 2008).
В отличие от ветвей суммарная биомасса стволов вертикальных «побегов» всех возрастных групп закономерно увеличивается с глубиной полога и в нижних его горизонтах стволы вносят существенный вклад (74 - 85%) в формировании общей биомассы.
Wp/S0(Z), W/S0(Z), мг(сух)/см2 (A/z = 5 см)
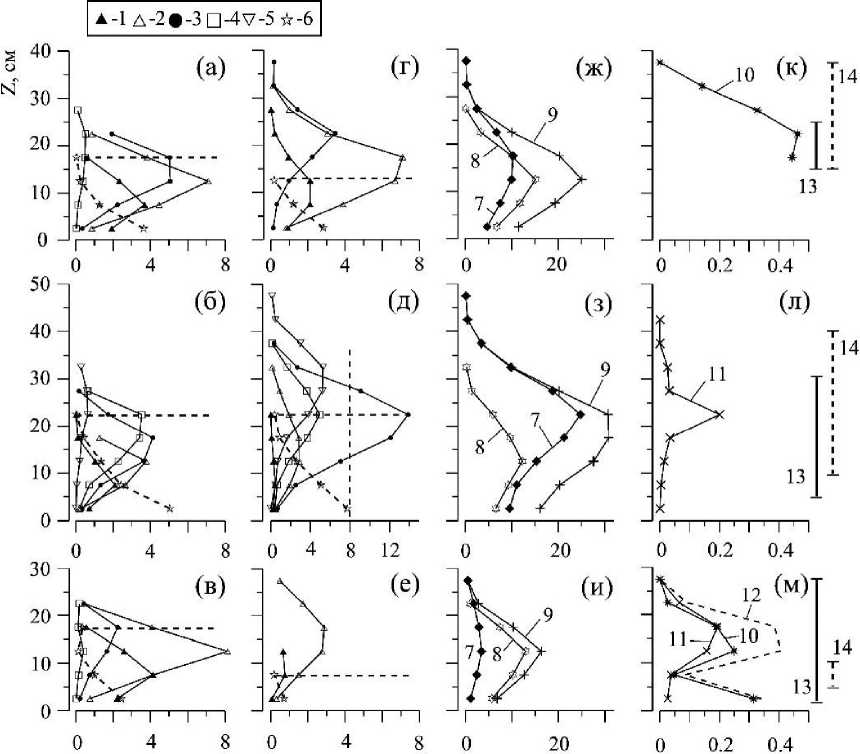
Рис. 6. Вертикальное распределение суммарной сухой массы WT/S0 (Z) ветвей каждой возрастной группы вертикальных «побегов» С. crinita (а - в) и С. barnata (г - е) (1 - 5), стволов каждого вида растений всех возрастных групп вместе (6), вертикальных побегов С. barbata (7) и С. crinita (8) в отдельности и вместе (9) и их эпифитов (к - м) в РП на станциях: 2 (а, г, ж, к), 3 (б, д, з, л), 6 (в, е, и , м). Возрастные группы вертикальных «побегов»: 1 - 0,13 < 1,9;
2 - 1,9 < 3,7; 3 - 3,7 < 5,4; 4 -5,4 < 7,2; 5 -7,2 < 8,9 лет. Эпифиты с обоих видов цистозир: 10 - Р. subulifera:, 11 - С. sericea; 12 - объединенная масса Р. subulifera и С. sericea. Вертикальными линиями показаны пределы вертикального распределения К. virgatula на слоевищах С. crinita (13) и С. barbata (14)
Наблюдаемые отличия в архитектонике растительного полога (рис. 5) на разных участках прибрежья главным образом объясняются различием в возрастной структуре ценопопуляций С. crinita. В каждом отдельном случае (рис. 5 а-г) между возрастными группами слоевищ на соответствующих горизонтах полога складываются характерные соотношения по биомассе. На станциях 1, 5, 7 ценопопуляции С. crinita представлены семью возрастными группами, и во всех этих случаях полог имеет одинаковую высоту, 37 см, а на станции 8 высота полога достигает 52 см при участии растений десяти возрастных групп.
Другая группа данных характеризует архитектонику растительного полога, складывающегося из вертикальных «побегов» двух видов цистозир (С. barbata, С. crinita) и их эпифитов в преимущественно закрытых акваториях прибрежья (рис. 36, г; 46; 6; станции: 2, 3, 4, 6). Как и в случае с пологом, формируемым вертикальными «побегами» С. crinita и её эпифитами, для полога из слоевищ двух видов цистозир и их эпифитов характерен одномодальный вид распределения W/So(z) (рис. Зг). Максимум биомассы располагается в нижней части полога, ближе к его серединной части; в нижней части полога сосредоточено около 80% фитомассы (рис. 4 б).
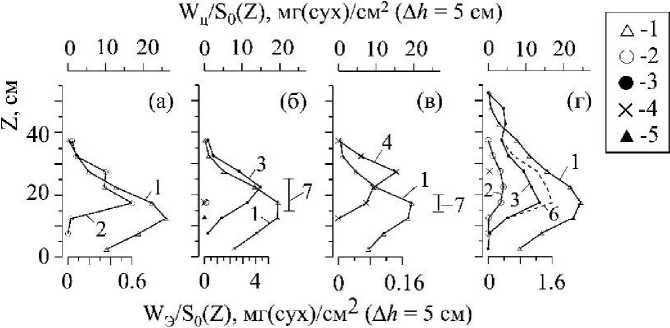
Рис. 7 . Вертикальное распределение сухой массы вертикальных побегов С. crinita (1, W4/So) и их эпифитов (2 - 6, W3/So) в РП на станциях: 1 (а), 5 (б), 7 (в), 8 (г). Эпифиты: L. coronopus (2), Р. subulifera (3), С. sericea (4), Grateloupia dichotoma J.Ag. (5), 6 - объединенная масса L. coronopus и P. subulifera. Вертикальной линией показаны пределы вертикального распределения К. virgatula на слоевищах С. crinita (7)
Упомянутые выше виды цистозир в соответствии с особенностями их морфологической структуры по-разному откликаются на условия подвижности воды (Ковардаков и др., 1985). Это может существенным образом отражаться в участии того или иного вида в создании полога, его вертикальной структуры. На станции 6 основная роль в формировании растительного полога принадлежит С. crinita (рис. 6 и), на станции 3 - С. barbata (рис. 6 з), а на станции 2 вклад обоих видов в общую биомассу примерно одинаков. У этих же видов наблюдается и несовпадение положений максимумов W/So(z) (рис. 6 ж, з). С. crinita не всегда охватывает всю толщину полога. Основная часть массы её ветвей располагается в пределах высоты её стволов (на рис. 5, 6 верхняя граница расположения стволов показана горизонтальной пунктирной линией). В то же время слоевища С. barbata размещаются по всей высоте полога и, тем самым, определяют его верхнюю границу, причём ветви этого вида располагаются как в пределах, так и далеко за пределами высоты стволов вертикальных побегов (рис. 6 г-е). На станции 6, в условиях сильной рекреационной нагрузки прибрежья, наблюдается существенное сокращение возрастного ряда слоевищ С. barbata, соответственно, здесь и наименьшая высота полога в рассматриваемом ряду станций.
Третья группа данных (рис. 6 к-л, 7, 8) позволяет рассмотреть вопрос о том, как в растительном пологе складываются отношения между двумя экологическими группами водорослей: макроэпифитами и их хозяевами. По условиям отбора проб в количественный учет эпифитов включались только виды макрофитов, играющих основную роль по массе в формировании полога. Кюлинию (Kylinia virgatula (Harv.) Papenf.), встречающуюся в небольших количествах по массе на стволах и на верхушках ветвей цистозир практически на всех станциях наблюдения, за исключением станций 1 и 8, количественно не оценивали.
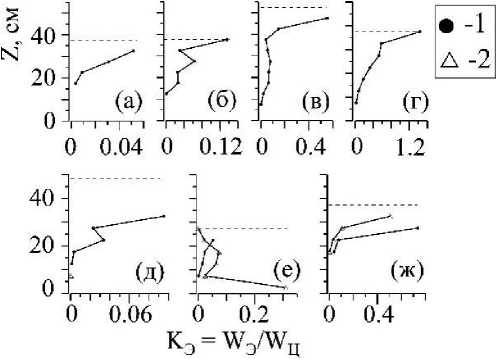
Рис. 8 . Весовая доля макроэпифитов по отношению к массе слоевищ С. crinita (1) и С. barbata (2) (W3/W4) на отдельно взятых горизонтальных участках полога на станциях: 7 (а), 1 (б), 8 (в), 5 (г), 3 (д), 6 (е), 2 (ж).
Для фитоценозов каждого отдельного участка прибрежья характерны определенные виды макроэпифитов, доминирующих по массе в эпифитных синузиях. На станции 1 таким видом являлся Laurencia coronopus J. Ag., на станциях 2 и 5 - Polysiphonia subulifera (С. Ag.) Harv., а на станциях 3 и 7 - Cladophora sericea (Huds.) Kutz. Ha двух станциях наблюдения заметный вклад в биомассу синузий вносят два вида водорослей: Р. subulifera и L. coronopus на станции 8, и Р. subulifera и С. sericea на станции 6.
Как и в случае с распределением W/So(z) цистозир, распределение биомассы макроэпифитов по вертикальному профилю полога описывается одновершинной зависимостью, при этом, в большинстве случаев, максимум биомассы эпифитов располагается выше максимума W/So(z) цистозир (рис. 6 к-л, 7). Весовая доля макроэпифитов по отношению к массе цистозир (Wg/Wp) на отдельно взятых горизонтальных участках полога хотя и увеличивается с приближением к верхней границе РП, но все же остаётся очень малой величиной (рис. 8).
На рис. 6 к-м, 7 вертикальными линиями показаны пределы вертикального распределения кюлинии в РП. В одних случаях это относительно узкий диапазон распределения и преимущественно в серединной части полога (рис. 7 б, в), в других - кюлиния охватывает значительную часть полога и, в силу особенностей каждого вида цистозир, по-разному заселяет их (рис. 6 к-м).
Обсуждение. Мощным экологическим фактором в прибрежной мелководной части моря, оказывающим влияние на возрастную структуру популяций цистозир и видовой состав фитосообщества, а с ней и на пространственную организацию полога, является подвижность воды. Другими, не менее важными факторами, определяющими архитектонику полога, являются солнечная радиация и уровень антропогенного воздействия. В нашем случае выделяются две группы участков прибрежья, визуально отличающиеся по характеру подвижности воды и по степени антропогенного воздействия. При всей очевидности различий в архитектонике пологов фитоценозов на этих участках прибрежья существуют общие закономерности в их формировании. На этом мы акцентируем внимание. Оптимальная структура полога понимается нами как структура при соответствующих возможностях выполнять фитоценозом максимальную фотосинтетическую работу. Исходя из этого, архитектоника любого естественно созданного растительного полога оптимально организована по отношению к соответствующим условиям окружающей среды и, в частности, к соответствующему уровню поступающей энергии.
Формирование вертикальной структуры (архитектоники) растительного полога, по мнению агрофизиологов (Росс, Росс, 1969; Росс, 1975; Тооминг, 1984), исследователей леса (Уткин, 1975) и водных фитосообществ (Завалко, Ковальчук, 1994), направлено на стабилизацию продукции в градиентах внешних переменных. Один из эффективных способов стабилизации - ярусная структура РП (Тооминг, 1984). Как было показано выше, верхний ярус цистозировой ассоциации имеет собственную многослойную, ярусную структуру из слоевищ разного возраста (рис. 4) и эпифитных синузий (рис. 3 б, в).
Как следует из представленных данных (рис. 5, 6), каждая возрастная группа слоевищ цистозир занимает по высоте полога характерный для них горизонтальный слой, где ее масса достигает максимума. И каждый такой горизонт, как правило, перекрывается слоевищами других возрастных групп. Уровень доминирования по массе той или иной возрастной группы в том или ином горизонтальном слое может варьироваться в широком диапазоне значений (35 - 100%) (Празукин, 2015а). При совместном произрастании двух видов цистозир, положение их максимумов биомассы (W/So(z) в пологе, как правило, разобщено (рис. бж, з). Согласно наших данных и данных других авторов (Завалко, Ковальчук, 1994), эпифитные синузии располагаются преимущественно в верхней части полога. В работе упомянутых авторов показано, что местоположение эпифитов по высоте полога во многом зависит от уровня солнечной радиации над ним. На глубине 0.5 м, в условиях достаточного количества света, расхождение максимумов биомассы слоевищ цистозиры и её эпифитов может и не наблюдаться. Тогда как заметное расхождение начинается с глубины 1.5 -2м (Завалко, Ковальчук, 1994) и в этих условиях достигается более равномерное распределение W/So(z) по профилю полога.
К выше сказанному добавим, что в наземных фитоценозах вертикальное распределение концентрации массы и поверхности разных видов в пологах также описывается серией одновершинных кривых, причем в большинстве случаев наблюдается расхождение максимумов (Карманова и др., 1987). Также известно (Тооминг, 1984), что в наземном многовидовом растительном пологе площадь ассимилирующей поверхности каждого вида наиболее велика в том слое РП, уровень освещенности которого соответствует максимальному значению коэффициента полезного действия газообмена данного вида, в нашем случае, у ветвей разных возрастных групп слоевищ цистозир и у растений эпифитных синузий. Благодаря этому, наземный фитоценоз во всем занимаемом им объеме усваивает свет с максимальным КПД (Тооминг, 1984), это может быть справедливо и по отношению к водным фитоценозам.
В заключение отметим, что адаптивные возможности полога цистозирового фитоценоза определяются многовариантностью сложения его структуры. И отражается это во взаимном расположении в пространстве морфологических элементов слоевищ цистозир разных возрастных групп, и во взаимном расположении отдельных видов эпифитных синузий как относительно друг друга, так и по отношению к их хозяину.
Выводы: 1) Для архитектоники растительного полога верхнего яруса цистозировых ассоциаций характерен одномодальный тип вертикального распределения биомассы с максимумом в его нижней половине, где сосредоточено от 75 до 85% всей сухой массы растений. 2) Распределение биомассы слоевищ разных возрастных групп цистозир по профилю полога описывается серией одновершинных кривых и, в большинстве случаев, наблюдается расхождение их максимумов и, возможно, это позволяет цистозирам с участием других групп водорослей стабилизировать продукционные характеристики на большей части вертикального профиля. 3) Виды эпифитных синузий располагаются преимущественно в верхней половине полога, где они в определенные сезоны года могут занимать доминирующее положение по массе и по площади поверхности и формировать собственный ярус. За счет этой группы растений достигается адаптивная лабильность растительного полога к топическим и сезонным условиям среды.
Список литературы Архитектоника растительных пологов цистозировых фитоценозов, Черное море
- Евстигнеева И.К. 2006. Макрофитобентос прибрежного эколона бухты Балаклавской (Черное море, Украина)//Альгология. Т. 16. № 2. С. 167-180.
- Завалко С.Е., Ковальчук Н.А. 1994. Ярусность как средство стабилизации и оптимизации вертикальной структуры сообщества черноморских макрофитов//Ботанический журнал. Т. 79. № 3. С. 30-39.
- Калугина А.А. 1969. Исследование донной растительности Черного моря с при-менением легководолазной техники//Морские подводные исследования. М.: Наука. С. 105-113.
- Калугина-Гутник А.А. 1975. Фитобентос Черного моря. Киев: Наук. Думка. 248 с.
- Карманова И. В. Судницына Т. Н., Ильина Н. А. 1987. Пространственная структура сложных сосняков. М.: Наука. 199. 200 с.
- Ковардаков С.А. Празукин А.В., Фирсов Ю.К. 2009. Сравнение многолетних изменений структурных и функциональных состояний прибрежных черноморских фитоценозов с различной антропогенной нагрузкой//Системы контроля окружающей среды: Средства, информационные технологии и мониторинг: сборник научных трудов. Севастополь: Морской Гидрофизический институт НАН Украины. С. 377-384.
- Ковардаков С.А., Празукин А.В., Фирсов Ю.К., Попов А.Е. 1985. Комплексная адаптация цистозиры к градиентным условиям. Киев: Наук. Думка. 216 с.
- Костенко Н.С., Дикий Е.А., Заклецкий А.А., Марченко В.С. 2005. Многолетние изучения в сообществах макрофитобентоса района Карадага (Крым, Черное море)//Морской Экологический журнал. Отд. Вып. 1. С. 48-60.
- Мильчакова Н.А., Рябогина В.Г. 2002 Флористическая характеристика морских акваторий объектов природно-заповедного фонда региона Севастополя (Черное море)//Экология моря. Вып. 60. С. 5-11.
- Миронова Н.В., Мильчакова Н.А., Александров В.В. 2012. Тенденции долговременного изменения запасов макрофитов в бухте Казачья (Севастополь, Крым, Черное море)//Морской экологический журнал. Т. 11. № 3. С. 68-78.
- Полевая геоботаника. 1964/ред. Корчагина А.А., Лавренко Е.М., Понятовский В.М. М.-Л.: Изд-во АН СССР. Т. 3. 530 с.
- Празукин А.В. 2008. Морфофункциональные образы растения (на примере многоклеточных водорослей)//Вестн. ТвГУ. Сер. Биология и экология. Вып. 9. С. 206-218.
- Празукин А.В. 2015а. Экологическая фитосистемология. М.: Перо. 375 с.
- Празукин А.В. 2015б. К вопросу об иерархии обитаемых пространств многоклеточных водорослей/ред. А.Г. Лапиров, Д.А. Филиппов, Э.Г. Гарин//Материалы VIII Всесоюзной конференции с международным участием по водным макрофитам, п. Борок, 16-20 октября 2015 Ярославль: Филигрань. С. 33-36.
- Празукин А.В., Фирсов Ю.К. 2013. Вертикальное распределение растительной массы в обитаемом пространстве ценоза многоклеточных водорослей (Чёрное море)//Системы контроля окружающей среды: сб. науч. тр./НАН Украины, Морской гидрофизический ин-т. Севастополь. Вып. 19. С. 253-257.
- Празукин А.В., Хайлов К.М. 1998. Пространственная организация полога диатомового обрастания на экспериментальных конструкциях в сравнении с другими фитосистемами//Гидробиол. журн. Т 34. № 5. С. 38-48.
- Росс Ю., Росс В. 1969. Пространственная ориентация листьев в посевах//Фотосинтетическая продуктивность растительного покрова. Тарту: ИФА АН ЭстССР. С. 60-82.
- Росс Ю.К. 1975. Радиационный режим и архитектоника растительного покрова. Л.: Гидрометеоиздат. 342 с.
- Сабурин М.Ю. 2004. Фитоценозы черноморской цистозиры: структура, восстановление и перспективы использования: автореф. дис. … канд. биол. наук. М.: МГУ. 24 с.
- Симакова У.В. 2011. Структура и распределение сообществ макрофитобентоса в зависимости от рельефа дна: автореф. дис. … канд. биол. наук. М.: МГУ. 22 с.
- Тооминг Х.Г. 1984. Экологические принципы максимальной продуктивности посевов. Л.: Гидрометеоиздат. 264 с.
- Уткин А.И. 1975. Биологическая продуктивность лесов//Итоги науки и техники. М.: ВИНИТИ. Т 1. С. 9-189.
- Шульгин И.А. 2013. Энергетический баланс и физиологические критерии продуктивности посевов сельскохозяйственных культур в условиях изменения климата//Труды Института глобального климата и экологии Росгидромета и РАН. Т. 25. С. 224-250.
- Cenzato D., Ganf G. 2001 A comparison of growth responses between two species of Potamogeton with contrasting canopy architecture//Aquatic Botany. V. 70. P. 53-66.
