Ассоциации нарушений ядерной ДНК с индивидуальными вариантами генов системы репарации ДНК APEX1, ERCC2, ERCC5, PARP1 и HOGGL у работников угольных теплоэлектростанций

Автор: Марущак А.В., Торгунакова А.В., Титов Р.А., Соболева О.А., Минина В.И.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Генетика
Статья в выпуске: 4, 2024 года.
Бесплатный доступ
Комбинации генетических вариантов и факторов окружающей среды ответственны за развитие многих заболеваний человека. Информация об индивидуальной способности накапливать повреждения ДНК необходима для развития разработки персонализированной профилактики заболеваний у населения, подвергшегося воздействию факторов окружающей среды. Угольные тепловые электростанции сжигают огромное количество угля и выделяют соответствующее количество токсичных газов, золы, летучих органических веществ. Элементы, входящие в состав угля, являются большой группой разнообразных загрязнителей, связанных с угрозами для здоровья человека и окружающей среды. Настоящая работа основана на определении рисков, которые связаны со здоровьем работников угольных теплоэлектростанций. Проведена оценка степени повреждений ДНК в связи с индивидуальной чувствительностью у 455 работников угольных теплоэлектростанций г. Кемерово и 533 жителей этого же региона, не связанных с производством. Для регистрации нарушений ДНК применялось микроядерное тестирование с цитокинетическим блоком на лимфоцитах периферической крови. Определение влияния генетических факторов на степень повреждения ДНК проводилось методом аллель-специфичной полимеразной цепной реакции полиморфных вариантов генов систем репарации ДНК APEX1 rs1130409, ERCC2 rs13181, PARP1 rs1136410. hOGG1 rs1052133 и ERCC5 rs17655. Выявлены повышенные частоты нарушений ДНК у носителей гетерозиготного генотипа гена APEX1 и минорных вариантов APEX1, ERCC2, hOGG1 и ERCC5.
Работники, угольные теплоэлектростанции, микроядерный тест, полиморфизм, гены репарации днк, apex1, ercc2, parp1, hogg1, ercc5
Короткий адрес: https://sciup.org/147247247
IDR: 147247247 | УДК: 575.22 | DOI: 10.17072/1994-9952-2024-4-421-432
Associations of nuclear DNA disorders with individual variants of the DNA repair system genes APEX1, ERCC2, ERCC5, PARP1 and HOGG1 in workers of coal thermal power plants
Combinations of genetic variants and environmental factors are responsible for the development of many human diseases. Information on individual capacity to accumulate DNA damages is essential to advance the development of personalized disease prevention in environmentally exposed populations. Coal-fired thermal power plants burn huge amounts of coal per year and emit corresponding amounts of toxic gases, ash, and volatile organic compounds. The elements that make up coal are a large group of diverse pollutants associated with threats to human health and the environment. The current work is based on identifying the risks associated with the health of workers at coal-fired thermal power plants. The study is devoted to the assessement of the DNA damage degree in connection with individual sensitivity in 455 workers of coal-fired thermal power plants in the city of Kemerovo and 533 residents of the same region not associated with production. The research materials are represented by 988 samples of peripheral venous blood from residents of the Kemerovo region. Control donors were selected according to the gender, age, and smoking status of coal-fired power plant workers. Micronucleus testing with a cytokinetic block on peripheral blood lymphocytes was used to register DNA disorders. Determination of the influence of genetic factors on the degree of DNA damage was carried out using allele-specific polymerase chain reaction of polymorphic variants of the DNA repair system genes APEX1 rs1130409, ERCC2 rs13181, PARP1 rs1136410, hOGGl rs1052133 and ERCC5 rs17655. Increased frequencies of DNA disorders were detected in carriers of the heterozygous genotype of the APEX1 gene and minor variants of APEX1, ERCC2, hOGG1 and ERCC5.
Текст научной статьи Ассоциации нарушений ядерной ДНК с индивидуальными вариантами генов системы репарации ДНК APEX1, ERCC2, ERCC5, PARP1 и HOGGL у работников угольных теплоэлектростанций
Оценка всех видов веществ с точки зрения безопасности для здоровья человека проводится при помощи тестов на изучение генотоксичности. Генотоксические факторы не только могут убивать клетки, но и повреждать их генетический материал [Sommer, Buraczewska, Kruszewski, 2020]. Некоторые повреждения могут сохраняться в клетке при условии ее выживаемости и наследоваться при делении [Sommer, Buraczewska, Kruszewski, 2020]. Методы определения генотоксичности включают в себя изучение обратимых и необратимых повреждений структуры ДНК, а также нарушение механизмов сохранения геномной стабильности [Corvi, Madia, 2017]. Особые виды хромосомных повреждений удается зафиксировать при помощи микроядерного теста. Анализ микроядер может способствовать понимаю механизмов взаимодействия генетических факторов и эффектов окружающей среды, поскольку он считается маркером воздействия мутагенов внешней среды и генетической нестабильности. Данный метод является стандартизированным, позволяет проводить анализ нескольких типов ядерных повреждений [Vral, Fenech, Thierens, 2011].
Совокупность генетических вариантов и факторов окружающей среды является предиктором развития целого ряда заболеваний человека. Репарационные механизмы ДНК определяют результаты после выявления нарушений ДНК и необходимы для поддержания стабильности генома [Wang et al., 2022]. Под геномной нестабильностью подразумевают склонность генома к формированию мутаций [López-Gi, Pascual-Ahuir, Proft, 2023]. Такие процессы происходят при дисфункции механизмов репарации и репликации или увеличении воздействия канцерогенных факторов на организм [Langie et al., 2015].
Целостность генома измеряется увеличенной встречаемостью мутаций в геноме, например, модификациями в нуклеотидных последовательностях [Ferragut Cardoso et al., 2021]. К потере стабильности генома может привести экзогенно-индуцированное повреждение ДНК, вызываемое различными химическими факторами окружающей среды, радиационными агентами, наночастицами [Langie et al., 2015].
Информация об индивидуальной чувствительности к повреждениям ДНК необходима для развития разработки персонализированной профилактики заболеваний у населения, подвергшегося воздействию окружающей среды. Оценка индивидуальной изменчивости имеет решающее значение в выявлении биомаркеров не только для первичной профилактики заболеваний, в том числе рака, и ранней диагностики, но и для их лечения благодаря индивидуально скорректированной терапии [Nagel, Chaim, Samson, 2014].
Кузбасс расположен в юго-западной части Сибири и является одним из крупнейших угольных бассейнов мира [Labzovskii, Belikov, Damiani, 2022]. В столице Кузбасса, г. Кемерово, расположены три теплоэлектростанции (ТЭС), использующие в качестве топлива каменный уголь, – Кемеровская Государственная районная электрическая станция (ГРЭС), Кемеровская теплоэлектростанция и НовоКемеровская теплоэлектростанция (ТЭЦ).
На угольных теплоэлектростанциях сжигается огромное количество угля в год и выделяется не менее огромный объем углекислого газа, оксидов азота и серы, летучих органических веществ, золы [Munawer,
-
2018] . В состав угля входят элементы, которые определяют большую группу опасных для здоровья человека веществ [Hossain, Paul, Hasan, 2015]. В список таких элементов входят тяжелые металлы, которые представляют серьезную угрозу для здоровья человека из-за их широкого распространения и долгого хранения, которое приводит к накоплению в биологических системах [Balali-Mood et al., 2021]. Помимо этого, установлено, что воздействие высокого уровня шума на работников повышено на угольных теплоэлектростанциях по сравнению с электростанциями на природном газе или гидроэлектростанциями [Sarker et al., 2010a, 2010b].
В связи с этим целью данной работы является анализ частот цитогенетических повреждений в связи с унаследованными вариантами генов ферментов репарации ДНК: APEX1 rs1130409 , ERCC2 rs13181 , PARP1 rs1136410. hOGG1 rs1052133 и ERCC5 rs17655 .
Материалы и методы исследования
Исследование проводилось в соответствии с рекомендациями Хельсинской декларации Всемирной ассоциации «Этические принципы проведения научных медицинских исследований с участием человека» с поправками 2000 г. и «Прaвилaми клинической прaктики в Российской Федерaции» (утвержденными Прикaзом Минздрaвa РФ от 19.06.2003 г. № 2660). Дизайн исследования (кандидатное исследование по принципу случай-контроль) был одобрен локальным этическим комитетом Федерального исследовательского центра угля и углехимии СО РАН (г. Кемерово). Участие людей в исследовании базировалось на принципах добровольности и информированности о целях, методах и результатах работы.
Материалы исследования представлены 988 образцами периферической венозной крови жителей Кемеровской области. Было обследовано 455 работников угольных теплоэлектростанций г. Кемерово и 533 условно-здоровых жителей этого же региона, не связанных с промышленностью и составивших группу контроля (табл. 1). Контрольные доноры подбирались соответственно полу, возрасту и статусу курения работников угольных теплоэлектростанций.
Таблица 1
Характеристика исследуемых групп [Characteristics of the studied groups]
|
Параметр |
Работники |
Группа сравнения |
|
|
Пол |
женщины |
101 |
168 |
|
мужчины |
354 |
365 |
|
|
Средний возраст, μ ± SEM |
51,9 ± 0.5 |
49,4 ± 4 |
|
|
Статус курения |
курящие |
169 |
170 |
|
некурящие |
286 |
363 |
|
|
Наличие хронических заболеваний |
обладают |
79 |
- |
|
не обладают |
161 |
- |
|
|
Всего |
455 |
533 |
|
Примечание. μ – среднее значение; SEM – стандартная ошибка среднего.
Для определения степени повреждения ядерной ДНК применялся микроядерный тест с блокировкой цитокинеза на лимфоцитах периферической крови с рекомендациями M. Fenech [2007] и модификациями Ф. И. Ингель [2006]. В культуральные флаконы вносили 3 мл среды RPMI 1640. 0.8 мл эмбриональной телячьей сыворотки, 0.25 мл венозной крови, 0.05 мл фитогемагглютинина и оставляли в инкубаторе при 37°С на 44 ч. По истечении 44 ч. в культуры вводили ингибитор цитокинеза цитохалазин В с конечной концентрацией 6 мкг/мл и продолжали инкубацию еще на 28 ч. Полученные клеточные культуры переносили в фальконы объемом 15 мл и центрифугировали 10 мин. при 1 000 rpm. Избавлялись от супернатанта, разбивали осадок и добавляли 6 мл охлажденного гипотонического раствора (0.125 М калий хлористый) для лизиса эритроцитов, перемешивали и добавляли 1 мл ледяного фиксатора Карнуа (3 объема метанола на 1 объем ледяной уксусной кислоты), перемешивали и центрифугировали 10 мин. при 1 000 rpm. Удаляли супернатант, разбивали осадок и вносили 9 мл ледяного метанол-уксусного фиксатора, перемешивали и выдерживали экспозицию при 4°С в течении 20 мин., центрифугировали 10 мин. при 1 000 rpm. Избавлялись от супернатанта, разбивали осадок и вносили 9 мл ледяного метанол-уксусного фиксатора, перемешивали и выдерживали экспозицию при 4°С на протяжении 50 мин. Центрифугировали 10 мин., удаляли супернатант и переносили полученный осадок во флаконы объемом 1.5 мл. Полученные суспензии переносили на предварительно обезжиренные и замороженные предметные стекла, выдерживали ночь и окрашивали 3% красителем Гимза. Полученные препараты просматривали на микроскопах Axio Scope.A1 (Zeiss, Германия) и Eclipse E100 (Nikon, Япония). Регистрировались 500 клеток любой ядерности, на стадии митоза и апоптоза, нарушения (микроядра, мосты, протрузии), после учитывали только двухъядерные лимфоциты и их нарушения, набирая количество данных клеток до 1 000 rpm. Критерии отбора двуядерных лимфоцитов, включаемых в анализ, и критерии для регистрации цитогене- тических нарушений соответствовали общепринятым рекомендациям1. Расчет индекса репликации проводили по формуле, предоставленной там же:
ИР
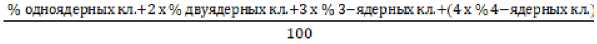
Выделение ДНК проводилось фенол-хлороформным методом. В анализ включены кодирующие ферменты репарации ДНК гены, для которых ранее установлены связи с повреждениями ДНК [Tavares et al., 2020; Liu et al., 2021; Xin et al., 2021] (изучены базы SNPinfo Web Server и RegulomeDB Version 1.1 ). Характеристика (по данным базы National Center for Biotechnology Information ) отобранных SNP генов представлена в табл. 2.
Таблица 2
Характеристика выбранных для исследования полиморфных вариантов
[Characteristics of polymorphic variants selected for study]
|
Ген |
SNP |
Локализация (GRCh38) |
Замена нуклеотидов |
|
APEX1 |
rs1130409 |
Chr 14:20456995 |
T> G |
|
ERCC2 |
rs13181 |
Chr 19:45351661 |
T> G |
|
PARP1 |
rs1136410 |
Chr 1:226367601 |
T> C |
|
hOGG1 |
rs1052133 |
Chr 3:9757089 |
C> G |
|
ERCC5 |
rs17655 |
Chr 13:102875652 |
G> C |
Примечание. SNP – однонуклеотидный полиморфизм.
Определение полиморфных вариантов генов APEX1 rs1130409, ERCC2 rs13181, PARP1 rs1136410. hOGG1 rs1052133 и ERCC5 rs17655 проводили с использованием наборов реактивов НПФ «Литех» (г. Москва) с последующим разделением продуктов амплификации методом горизонтального электрофореза в 3% агарозном геле. Визуализацию результатов проводили универсальной системой гель-документации Gel Doc (BioRad, США).
Статистическая обработка полученных данных проводилась с использованием программного обеспечения StatSoft Statistica 10.0. Соответствие выборки нормальному распределению проверялось критерием Колмогорова-Смирнова. Исходя из вида распределения, применялись методы непараметрической статистики, в частности U-критерий Манна-Уитни, с целью определения различий между изучаемыми выборками. Критический уровень значимости принимался равным 0.05. Оценка частоты редкого аллеля, соответствие распределения частот равновесию Харди–Вайнберга (χ2), статистическую значимость различий между группами по частотам аллелей и генотипов для теста χ2 на гомогенность выборок выполняли при помощи онлайн-ресурса:
Результаты и обсуждение
Для определения влияния угольной производственной среды на формирование цитогенетических нарушений проведено сравнение показателей микроядерного теста работников угольных теплоэлектростанций с группой неработающих жителей той же области (табл. 3).
Выявлены повышенные частоты ( p < 0.000001) лимфоцитов с микроядрами, мостами и протрузиями, а также частота клеток на стадии апоптоза у работников угольных теплоэлектростанций по сравнению с контрольными донорами. Однако наблюдается повышенная встречаемость лимфоцитов на этапе клеточного деления ( p < 0.000001) у жителей Кемеровской обл.
Микроядро представляет собой небольшую хроматин-содержащую структуру, наблюдаемую в клеточной цитоплазме [Fenech, 2007]. Микроядра могут образовываться в результате естественных процессов, таких как метаболизм или старение, или как результат взаимодействия с разнообразными факторами окружающей среды и многими заболеваниями [Sommer, Buraczewska, Kruszewski, 2020].
Формирование цитогенетических нарушений обусловлено несколькими механизмами: 1) отставание хромосомы во время анафазы клеточного деления [Liu, Pellman, 2020]; 2) разрушение хромосом (хромот-рипсис) в результате ошибок на стадии митоза и неспособности разрушить оболочку микроядра, содержащего хромосомные фрагменты; 3) локальные нарушения репликации ДНК, приводящие к перестройкам [Hatch, 2018].
Установлено влияние пола ( p < 0.02) и возраста ( p < 0.01) работника угольных теплоэлектростанций на накопление цитогенетических нарушений. Статус курения и хронические заболевания не влияют на формирование цитогенетических нарушений у работников угольных теплоэлектростанций ( p > 0.05). Выявлено влияние стажа работы на угольном производстве ( p < 0.0005) и профессиональной специализации ( p < 0.009) на образование геномных нарушений и пролиферативные показатели.
-
1 ГОСТ 32635-2014. Методы испытания по воздействию химической продукции на организм человека. Микро-ядерный тест на клетках млекопитающих in vitro. М.: Стандартинформ, 2019. 14 с.
Таблица 3
[Parameters of the micronucleus test in the studied groups]
|
Показатель |
Рабочие теплоэлектростанций (n = 455) |
Группа сравнения (n = 533) |
||||||
|
Медиана |
Мода |
Нижний квартиль |
Верхний квартиль |
Медиана |
Мода |
Нижний квартиль |
Верхний квартиль |
|
Частота двухъядерных клеток с цитогенетическими нарушениями (%)
|
Микроядра |
3.3* |
3.6 |
1.9 |
4.5 |
0.6 |
0.8 |
0.4 |
0.9 |
|
Мосты |
1.5* |
0.5 |
0.8 |
2.3 |
0.3 |
0 |
0 |
0.5 |
|
Протрузии |
2* |
0 |
1.2 |
3 |
0.3 |
0 |
0 |
0.5 |
Пролиферативные показатели (%)
|
Ядерность клеток |
1 |
31.2 |
Мн. |
24.6 |
37.8 |
32.1 |
Мн. |
25.2 |
38.4 |
|
2 |
45.8 |
48.2 |
40.6 |
50.8 |
44.0 |
43.6 |
39.6 |
50.2 |
|
|
3 |
7 |
8.4 |
4.2 |
9.2 |
6.2 |
4.4 |
4.4 |
8.8 |
|
|
4 |
15.2 |
11.2 |
10.8 |
18.6 |
14.5 |
17.4 |
10.4 |
17.2 |
|
|
Митоз |
0.6 |
0.4 |
0.4 |
1 |
2.4* |
2.2 |
1.3 |
3.0 |
|
|
Апоптоз |
1.2* |
0.2 |
0.4 |
1.8 |
0.4 |
0.2 |
0.2 |
1.0 |
|
|
Индекс репликации |
1.9 |
Мн. |
1.7 |
2.1 |
2.0 |
1.9 |
1.8 |
2.1 |
|
Примечание. * – отличается от группы сравнения по критерию Манна-Уитни: * р < 0.000001.
Параметры микроядерного теста в изученных группах
Частоты встречаемости цитогенетических нарушений в клетках крови в связи с различными полиморфными вариантами генов APEX1 rs1130409 , ERCC2 rs13181 , PARP1 rs1136410. hOGG1 rs1052133 и ERCC5 rs17655 работников угольных теплоэлектростанций представлены в табл. 4.
Таблица 4
Цитогенетические показатели микроядерного теста у рабочих теплоэлектростанций с различными вариантами изученных генов
[Cytogenetic parameters of the micronucleus test in thermal power plant workers with different variants of the studied genes]
|
Цитогенетические показатели |
Медиана |
Квартили (25-75%) |
Медиана |
Квартили (25-75%) |
Медиана |
Квартили (25-75%) |
APEX1 rs1130409
|
SNP |
TT (n = 138) |
TG (n = 234) |
GG (n = 83) |
|||
|
Микроядра |
3,3 |
1,9-4,5 |
3,6*1 |
2-4,9 |
3 |
1,8-3,9 |
|
Мосты |
1,5 |
0.8-2,3 |
1,5 |
0.8-2,4 |
1,4 |
0.7-2,2 |
|
Протрузии |
2,1 |
1,1-2,9 |
1,9 |
1,1-3,2 |
2,1*2 |
1,5-2,9 |
ERCC2 rs13181
|
SNP |
TT (n = 191) |
TG (n = 217) |
GG (n = 47) |
|||
|
Микроядра |
3,3 |
1,9-4,5 |
3,3 |
2-4,6 |
3 |
1,3-4,2 |
|
Мосты |
1,4 |
0.9-2,4 |
1,4 |
0.9-2,3 |
1.7*3 |
0.8-2,4 |
|
Протрузии |
2 |
1,2-2,2 |
2,1 |
1,2-2,3 |
2,3*3 |
1,2-3,2 |
PARP1 rs1136410
|
SNP |
CC (n = |
245) |
CG (n = |
180) |
GG (n |
= 30) |
|
Микроядра |
3,5 |
1,6-4,6 |
2,9 |
1,7-4,3 |
3,7*4 |
2,2-4,8 |
|
Мосты |
1,5 |
0.9-2,5 |
1,4 |
0.8-2,2 |
1,4 |
0.6-2,4 |
|
Протрузии |
2 |
1,1-3,1 |
2 |
1,2-3 |
2,2 |
1,3-2,9 |
ERCC5 rs17655
|
SNP |
GG (n = 279) |
GC (n = 149) |
CC (n = 27) |
|||
|
Микроядра |
3,3 |
2-4,5 |
3,3 |
1,7-4,6 |
3,6*5 |
1,9-4,4 |
|
Мосты |
1,4 |
0.8-2,4 |
1,5 |
0.8-2,3 |
1,7*5 |
0.7-2,8 |
|
Протрузии |
1,8 |
1,2-2,8 |
2 |
1,1-3,2 |
1,9 |
1,5-3 |
Примечание. *1 TC от TT и GG p = 0.04; *2 GG от TT и TG p = 0.02; *3 GG от TT и TG p < 0.01; *4 GG от CC и GC p = 0.004; *5 CC от GG и GC p < 0.002.
Установлены повышенные частоты клеток с микроядрами у носителей генотипа TG гена APEX1 ( p = 0.04). Также носители полиморфного варианта GG того же гена обладают повышенной частотой лимфоцитов с протрузиями ( p = 0.02). У обладателей минорного генотипа гена ERCC2 наблюдаются повышенные частоты встречаемости клеток крови с мостами и протрузиями ( p = 0.01). Носители минорного генотипа GG гена hOGG1 обладают повышенной частотой встречаемости клеток с микроядрами ( p < 0.01). У работников с минорным генотипом CC гена ERCC5 установлены повышенные показатели клеток с микроядрами и мостами (p < 0.002).
Не выявлено статистически значимых отличий между генотипами гена PARP1 rs1136410 ( p > 0.05).
Частоты встречаемости цитогенетических нарушений в клетках крови в связи с различными полиморфными вариантами генов APEX1 rs1130409 , ERCC2 rs13181 , PARP1 rs1136410. hOGG1 rs1052133, ERCC5 rs17655 контрольных доноров представлены в табл. 5.
Таблица 5
Цитогенетические показатели микроядерного теста у индивидов контрольной группы с различными вариантами изученных генов
[Cytogenetic parameters of the micronucleus test in individuals of the control group with different variants of the studied genes]
|
Цитогенетические показатели |
Медиана |
Квартили (25-75 %) |
Медиана |
Квартили (25-75 %) |
Медиана |
Квартили (25-75 %) |
|
APEX1 rs1130409 |
||||||
|
SNP |
TT (n = 280) |
TG (n = 221) |
GG (n = 32) |
|||
|
Микроядра |
0.76 |
0.5-1.63 |
0.7 |
0.58-1.01 |
0.6 |
0.6-0.8 |
|
Мосты |
0.49 |
0.2-1.13 |
0.4 |
0.2-0.8 |
0.2 |
0.2-0.4 |
|
Протрузии |
0.51 |
0.3-1.4 |
0.6 |
0.3-1.7 |
0.7 |
0.3-1.5 |
|
ERCC2 rs13181 |
||||||
|
SNP |
TT (n = 272) |
TG (n = 225) |
GG (n = 36) |
|||
|
Микроядра |
0.7 |
0.6-1.4 |
0.8 |
0.6-1.6 |
0.5 |
0.3-0.9 |
|
Мосты |
0.46 |
0.2-1.14 |
0.4 |
0.2-0.9 |
0.25 |
0-0.4 |
|
Протрузии |
0.5 |
0.3-1.43 |
0.5 |
0.3-1.5 |
0.35 |
0-1.2 |
|
PARP1 rs1136410 |
||||||
|
SNP |
TT (n = 311) |
TC (n = 192) |
CC (n = 30) |
|||
|
Микроядра |
0.8 |
0.6-1.4 |
0.7 |
0.6-0.9 |
0.5 |
0.5-0.5 |
|
Мосты |
0.42 |
0.2-1.03 |
0.4 |
0.2-0.93 |
0.4 |
0.1-0.4 |
|
Протрузии |
0.5 |
0.3-1.4 |
0.7 |
0.2-1.5 |
1.7* |
1.4-2.2 |
|
hOGG1 rs1052133 |
||||||
|
SNP |
CC (n = 325) |
CG (n = 185) |
GG (n = 23) |
|||
|
Микроядра |
0.6 |
0.4-0.8 |
0.6 |
0.4-0.9 |
0.5 |
0.3-0.9 |
|
Мосты |
0.3 |
0-0.5 |
0.4 |
0-0.5 |
0.4 |
0.1-0.5 |
|
Протрузии |
0.3 |
0-0.5 |
0.3 |
0-0.5 |
0.5 |
0.1-0.5 |
|
ERCC5 rs17655 |
||||||
|
SNP |
GG (n = 290) |
GC (n = 210) |
CC (n = 33) |
|||
|
Микроядра |
0.5 |
0.3-0.8 |
0.6 |
0.3-0.8 |
0.4 |
0.2-0.8 |
|
Мосты |
0.3 |
0-0.5 |
0.3 |
0-0.5 |
0.4 |
0-0.5 |
|
Протрузии |
0.3 |
0-0.5 |
0.3 |
0-0.5 |
0.4 |
0-0.6 |
|
Примечание. * CC от TT и TC p = 0.04. |
||||||
В группе контроля наблюдается повышенная частота встречаемости клеток с протрузиями у обладателей генотипа CC гена PARP1 ( p = 0.04).
В остальных исследуемых случаях в контрольной группе статистически значимых отличий выявлено не было ( p > 0.05).
Апуриновая/апиримидиновая эндонуклеаза 1/окислительно-восстановительный эффекторный фактор 1 (APEX1/Ref-1) является многофункциональным ферментом, который необходим для поддержания клеточного гомеостаза. APEX1 является основной апуриновой/апиримидиновой эндонуклеазой в пути эксцизионной репарации оснований и действует как окислительно-восстановительный регулятор нескольких транскрипционных факторов, включая NF-κB, AP-1, HIF-1α и STAT3. Эти функции делают APEX1 жизненно важным для регуляции клеточного цикла, старения и воспалительных путей. В дополнение к регуляции экспрессии цитокинов и хемокинов путем активации окислительно-восстановительных факторов транскрипции, APEX1 участвует в других процессах иммунного ответа, включая производство активных форм кислорода. Кроме того, участвуя в активном деметилировании хроматина, функция APEX1 заключается в регуляции транскрипции некоторых генов, включая цитокины, такие как TNFα [Oliveira et al., 2022].
Ген АДФ-рибозилтрансферазы ( PARP1 ) кодирует хроматин-ассоциированный фермент, поли(АДФ-рибозу) полимеразу-1 (PARP-1), и синтезирует после повреждения ДНК и участвует в целостности генома, регулируя клеточные реакции на повреждение ДНК и апоптоз [Gadad et al., 2021]. Участвует в регуляции различных важных клеточных процессов, таких как дифференцировка, пролиферация и трансформация опухоли, а также в регуляции молекулярных событий, участвующих в восстановлении клетки после повреждения ДНК [Zakharenko et al., 2023].
Пигментная ксеродерма группы D ( ERCC2 ) является основным белком, участвующим в NER. Являясь важным геном репарации ДНК, он играет важную роль в белково-опосредованном комплексном NER транскрипционного фактора IIH (TFIIH) и его процессе транскрипции [Wang et al., 2020].
Ген hOGG1 расположен на хромосоме 3p26.2 и кодирует гликозилазу, катализирующую эксцизию 8-оксогуаниновых аддуктов из поврежденной ДНК. Во многих исследованиях подтверждена связь между полиморфизмом hOGG1 rs1052133 и риском развития гинекологического рака в европейской популяции [Shi, Xu, Zhang, 2020]. Полиморфизм hOGG1 rs1052133 , наиболее изученный полиморфизм hOGG1, влияет на активность белка и, таким образом, способствует канцерогенезу [Zhang et al., 2017].
XPG , также называемая ERCC5 , является основной эндонуклеазой DDR, дефицит которой приводит к серьезным дефектам развития, прогерии и раку. Он в основном известен своей ролью в вырезании повреждений ДНК в эксцизионной репарации нуклеотидов (NER), но в последние годы было обнаружено, что он функционирует и в других механизмах поддержания генома. ERCC5 входит в семейство структурно-специфических нуклеаз XPG/RAD2, которое у млекопитающих также включает FEN1, GEN1 и EXO1 и выполняет важные функции поддержания генома. Наследственные мутации гена ERCC5 связаны с несколькими заболеваниями человека, характеризующимися широким спектром симптомов, включая предрасположенность к раку, прогрессирующую нейродегенерацию и нарушение развития [Muniesa-Vargas et al., 2022]. Некоторые мутации ERCC5 вызывают пигментную ксеродерму (XP), которая характеризуется фоточувствительностью, аномальной пигментацией кожи, повышенным риском развития рака и иногда неврологическими заболеваниями [Rizza et al., 2021].
Важность репарации ДНК для поддержания целостности генома и предотвращения развития опухолей подчеркивается тем фактом, что дефекты в репарации ДНК связаны с повышенной восприимчивостью клеток к токсическим, мутагенным и канцерогенным эффектам воздействия окружающей среды [Huang, Zhou, 2021].
Важность удаления продуктов окисления ДНК отражается в избыточности ферментов репарации ДНК для этих поражений. Из-за этой избыточности определение роли АФК в развитии рака и ферментов репарации в его предотвращении является сложной задачей. Однако вмешательство окружающей среды в восстановление таких повреждений может способствовать развитию рака [Langie et al., 2015].
Одним из примеров связи между индукцией АФК и развитием рака является повышенная заболеваемость раком легких, кожи, мочевого пузыря и печени у людей, которые подвергались воздействию агента, индуцирующего АФК, мышьяка [Wang et al., 2012]. Кроме того, было показано, что оксид азота ингибирует ферменты репарации ДНК [Khan et al., 2020]. Также сообщалось, что репарация нуклеотидов может быть ингибирована окислительным стрессом [Van Houten, Santa-Gonzalez, Camargo, 2018].
Генетическая изменчивость в генах репарации ДНК может модулировать восприимчивость к повреждению ДНК [Angelini et al., 2012]. В связи с этим в работе проведена оценка влияния полиморфизмов генов репарации ДНК APEX1 , ERCC2, PARP1. hOGG1 , ERCC5 на формирование микроядер, мостов и протрузий, выявляемых при помощи микроядерного теста.
В работе [Angelini et al., 2005] изучена частота микроядер в группе, подвергшейся воздействию низких уровней ионизирующего излучения, и в соответствующей контрольной группе. Как в исследуемой выборке, так и в контрольной группе также оценивалось возможное влияние однонуклеотидных полиморфизмов генов XRCC1, XRCC3 и ERCC2 на частоту микроядер. Также было рассмотрено влияние статуса курения, возраста и пола. Авторами установлены повышенные частоты подвергшихся воздействию работников, обладающими аллелем дикого типа гена ERCC2 , чем группа сравнения. Статус курения статистически значимо не влиял на частоту встречаемости микроядер в обеих группах. Однако увеличение возраста связано с повышением частоты клеток с микроядрами у работников. Также при объединении обеих групп исследования у женщин наблюдались повышенные частоты лимфоцитов с микроядрами.
У рабочих-носителей генотипа CG гена hOGG1 выявлена повышенная чувствительность к хромосомным повреждениям, выраженная в увеличенном формировании микроядер.
В исследовании [Qiu et al., 2011] рассматривалась взаимосвязь между генетическим полиморфизмом семи генов системы репарации ДНК (XPA, ERCC2, XPC, ERCC5, XPF, ERCC1 и APEX1) и восприимчивостью к хромосомному повреждению после воздействия винилхлорида. Было исследовано сто восемьдесят рабочих, подвергшихся профессиональному воздействию, и 43 контрольных донора, не подвергших- ся воздействию. У работников с вариантом минорного генотипа ERCC2 частота микроядер была незначительно выше (p = 0.07).
Sinitsky M.Y. с соавт. [2015] изучал влияние полиморфных вариантов генов системы репарации на частоту микроядер у детей, проживающих в школе-интернате г. Таштагол (Кемеровская обл., Россия). Выявлено достоверное увеличение частоты двуядерных лимфоцитов с микроядрами у носителей полиморфизма генотипа CC гена ERCC5 по сравнению с гетерозиготными и гомозиготными носителями. Кроме того, генотип CC гена ADPRT ассоциировался с повышенной частотой двуядерных лимфоцитов с нук-леоплазматическими мостами.
Этим же коллективом [Sinitsky et al., 2017] установлена повышенная частота двуядерных лимфоцитов с микроядрами у шахтеров с генотипом GG гена hOGG1 rs1052133 по сравнению с носителями генотипа CC.
В исследовании [Angelini et al., 2012] оценено влияние полиморфизмов генов репарации ДНК ( APEX1, hOGG1, NBS1, ERCC2, XRCC1 и XRCC3 ) на формирование микроядер как биомаркера ранних биологических эффектов при контакте с бензолом. Значительно более высокая частота микроядер была зарегистрирована у регулировщиков дорожного движения, чем в контрольной группе.
Cheng J с соавт. [2007] изучали ассоциации между несколькими генетическими полиморфизмами генов эксцизионной репарации нуклеотидов ( ERCC1, ERCC2, ERCC4, ERCC5 и ERCC6 ) и уровнем повреждения хромосом у 140 коксохимических рабочих, подвергшихся воздействию высокого уровня полиа-роматических углеводородов (ПАУ), и у 66 доноров, не подвергавшихся воздействию. Анализ показал значительную связь между полиморфизмом ERCC2 G23591A и частотами параметров микроядерного теста среди пожилых рабочих коксовых печей. Результаты показывают, что полиморфизмы ERCC1 C19007T, ERCC6 A3368G и ERCC2 G23591A связаны с частотами показателей микроядерного теста среди коксовых рабочих.
Заключение
В работе подтверждено влияние производственной среды угольных теплоэлектростанций на формирование повреждений ДНК в виде повышенных частот микроядер, мостов и протрузий в клетках крови работников.
Установлены ассоциации минорного варианта ERCC2 rs13181 с повышенной частотой нуклеоплаз-менных мостов и протрузий, минорного варианта ERCC5 rs17655 с увеличением формирования микроядер и нуклеоплазменных мостов, гетерозиготного варианта гена APEX1 rs1130409 с повышенной частотой микроядер, минорного генотипа этого же гена с частотой клеток с протрузиями, минорного генотипа hOGG1 rs1052133 c микроядрами при воздействии на организм работников промышленной среды угольных теплоэлектростанций. Обладатели данных генотипических вариантов должны стать приоритетной группой при проведении профилактических мероприятий. Полученные результаты свидетельствуют о значимом вкладе генетических и средовых факторов в формирование генотоксических эффектов у работников угольных теплоэлектростанций.
Список литературы Ассоциации нарушений ядерной ДНК с индивидуальными вариантами генов системы репарации ДНК APEX1, ERCC2, ERCC5, PARP1 и HOGGL у работников угольных теплоэлектростанций
- Ингель Ф.И. Перспективы использования микроядерного теста на лимфоцитах крови человека, культивируемых в условиях цитокинетического блока. Ч. 2. Факторы среды и индивидуальные особенности в системе нестабильности генома человека. Дополнительные возможности теста. Методика проведения экспериментов и цитогенетического анализа // Экологическая генетика. 2006. Т. 4, № 4. С. 38-54.
- Angelini S. et al. Micronuclei in humans induced by exposure to low level of ionizing radiation: influence of polymorphisms in DNA repair genes // Mutat. Res. 2005. Vol. 570, № 1. P. 105-117. DOI: 10.1016/j.mrfmmm.2004.10.007.
- Angelini S. et al. Environmental exposure to benzene, micronucleus formation and polymorphisms in DNA-repair genes: a pilot study // Mutat. Res. 2012. Vol. 743, № 1-2. P. 99-104. DOI: 10.1016/j.mrgentox.2011.10.018.
- Balali-Mood M. et al. Toxic Mechanisms of Five Heavy Metals: Mercury, Lead, Chromium, Cadmium, and Arsenic // Front Pharmacol. 2021. Vol. 12. Art. 643972. DOI: 10.3389/fphar.2021.643972.
- Cheng J. et al. Association between nucleotide excision repair gene polymorphisms and chromosomal damage in coke-oven workers // Biomarkers. 2007. Vol. 12, № 1. P 76-86. DOI: 10.1080/13547500600950168.
- Corvi R., Madia F. In vitro genotoxicity testing-Can the performance be enhanced? // Food Chem. Toxicol. 2017. Vol. 106(Pt B). P. 600-608. DOI: 10.1016/j.fct.2016.08.024.
- Fenech M. Cytokinesis-block micronucleus cytome assay // Nat. Protoc. 2007. Vol. 2, № 5. P. 1084-1104. DOI: 10.1038/nprot.2007.77.
- Fenech M. Cytokinesis-Block Micronucleus Cytome Assay Evolution into a More Comprehensive Method to Measure Chromosomal Instability // Genes. 2020. Vol. 11, № 10. Art. 1203. DOI: 10.3390/genes11101203.
- Ferragut Cardoso A.P. et al. miRNA dysregulation is an emerging modulator of genomic instability // Semin. Cancer Biol. 2021. Vol. 76. P. 120-131. DOI: 10.1016/j.semcancer.2021.05.004.
- Gadad S.S. et al. PARP-1 Regulates Estrogen-Dependent Gene Expression in Estrogen Receptor aPositive Breast Cancer Cells // Mol. Cancer Res. 2021. Vol. 19, № 10. P. 1688-1698. DOI: 10.1158/1541-7786.MCR-21-0103.
- Hatch E.M. Nuclear envelope rupture: little holes, big openings // Curr. Opin. Cell Biol. 2018. Vol. 52. P. 66-72. DOI: 10.1016/j.ceb.2018.02.001.
- Hossain M.N., Paul S.K., Hasan M.M. Environmental impacts of coal mine and thermal power plant to the surroundings of Barapukuria, Dinajpur, Bangladesh // Environ. Monit. Assess. 2015. Vol. 187, № 4. Art. 202. DOI: 10.1007/s10661-015-4435-4.
- Huang R., Zhou P.K. DNA damage repair: historical perspectives, mechanistic pathways and clinical translation for targeted cancer therapy // Signal Transduct. Target Ther. 2021. Vol. 6, № 1. Art. 254. DOI: 10.103 8/s41392-021-00648-7.
- Khan F.H. et al. The Role of Nitric Oxide in Cancer: Master Regulator or NOt? // Int. J. Mol. Sci. 2020. Vol. 21, № 24. Art. 9393. DOI: 10.3390/ijms21249393.
- Labzovskii L.D., Belikov D.A., Damiani A. Spaceborne NO2 observations are sensitive to coal mining and processing in the largest coal basin of Russia // Scto Rep. 2022. Vol. 12, № 1. Art. 12597. DOI: 10.1038/s41598-022-16850-8.
- Langie S.A. et al. Causes of genome instability: the effect of low dose chemical exposures in modern society // Carcinogenesis. 2015. Vol. 36, № 1. P. 61-88. DOI: 10.1093/carcin/bgv031.
- Liu J. et al. Association between APEX1 rs1760944 and rs1130409 polymorphism with prostate cancer risk: A systematic review and meta-analysis // Medicine (Baltimore). 2021. Vol. 100, № 46. Art. e27630. DOI: 10.1097/MD.0000000000027630.
- Liu S., Pellman D. The coordination of nuclear envelope assembly and chromosome segregation in meta-zoans // Nucleus. 2020. Vol. 11. P. 35-52. DOI: 10.1080/19491034.2020.1742064.
- Lopez-Gil L., Pascual-Ahuir A., Proft M. Genomic Instability and Epigenetic Changes during Aging // Int. J. Mol. Sci. 2023. Vol. 24, № 18. Art. 14279. DOI: 10.3390/ijms241814279.
- Munawer M.E. Human health and environmental impacts of coal combustion and post-combustion wastes // Journal of Sustainable Mining. 2018. Vol. 17, № 2. P. 87-96. DOI: 10.1016/j.jsm.2017.12.007.
- Muniesa-Vargas A. et al. XPG: a multitasking genome caretaker // Cell Mol. Life Sci. 2022. Vol. 79, № 3. Art. 166. DOI: 10.1007/s00018-022-04194-5.
- Nagel Z.D., Chaim I.A., Samson L.D. Inter-individual variation in DNA repair capacity: a need for multi-pathway functional assays to promote translational DNA repair research // DNA Repair (Amst). 2014. Vol. 19. Art. 199-213. DOI: 10.1016/j.dnarep.2014.03.009.
- Oliveira T.T. et al. APEX1/Ref-1 Role in Inflammation and Immune Response // Front Immunol. 2022. Vol. 13. Art. 793096. DOI: 10.3389/fimmu.2022.793096.
- Qiu Y.L. et al. DNA repair gene polymorphisms and micronucleus frequencies in Chinese workers exposed to vinyl chloride monomer // Int. J. Hyg. Environ. Health. 2011. Vol. 214, № 3. P. 225-230. DOI: 10.1016/j.ijheh.2010.12.001.
- Rizza E.R.H. et al. Xeroderma Pigmentosum: A Model for Human Premature Aging // J. Invest. Dermatol. 2021. Vol. 141, № 4S. P. 976-984. DOI: 10.1016/j.jid.2020.11.012.
- Sarker S.K. et al. Effect of Barapukuria coal mine on soil environment // Journal of Agro for Environment. 2010a. Vol. 4, № 2. P. 181-183.
- Sarker S.K. et al. Effect of Barapukuria power plant on environment // Journal of Agro for Environment. 2010b. Vol. 4, № 2. P. 125-127.
- Shi Y., Xu W., Zhang X. Association of the hOGG1 Ser326Cys polymorphism with gynecologic cancer susceptibility: a meta-analysis // Biosci. Rep. 2020. Vol. 40, № 12. Art: BSR20203245. DOI: 10.1042/BSR20203245.
- Sinitsky M.Y. et al. Association of DNA repair gene polymorphisms with genotoxic stress in underground coal miners // Mutagenesis. 2017. Vol. 32, № 5. P. 501-509. DOI: 10.1093/mutage/gex018.
- Sinitsky M.Y. et al. Associations of DNA-repair gene polymorphisms with a genetic susceptibility to ionizing radiation in residents of areas with high radon (222Rn) concentration // Int. J. Radiat. Biol. 2015. Vol. 9, № 6. P. 486-494. DOI: 10.3109/09553002.2015.1012306.
- Sommer S., Buraczewska I., Kruszewski M. Micronucleus Assay: The State of Art, and Future Directions // Int. J. Mol. Sci. 2020. Vol. 21, № 4. Art. 1534. DOI: 10.3390/ijms21041534.
- Tavares C.B. et al. Association between Single Nucleotide Polymorphisms and Glioma Risk: A Systematic Literature Review // Cancer Invest. 2020. Vol. 38, № 3. P. 169-183. DOI: 10.1080/07357907.2020.1719502.
- Van Houten B., Santa-Gonzalez G.A., Camargo M. DNA repair after oxidative stress: current challenges // Curr. Opin. Toxicol. 2018. Vol. 7. P. 9-16. DOI: 10.1016/j.cotox.2017.10.009.
- Vral A., Fenech M., Thierens H. The micronucleus assay as a biological dosimeter of in vivo ionising radiation exposure // Mutagenesis. 2011. Vol. 26, № 1. P. 11-17. DOI: 10.1093/mutage/geq078.
- Wang T.S. et al. Lymphocyte-based challenge DNA-repair assays for personalized health risk assessment // Mutat. Res. Rev. 2022. Vol. 790. Art. 108427. DOI: 10.1016/j.mrrev.2022.108427.
- Wang X. et al. Arsenic and chromium in drinking water promote tumorigenesis in a mouse colitis-associated colorectal cancer model and the potential mechanism is ROS-mediated Wnt/ß-catenin signaling pathway // Toxicol. Appl. Pharmacol. 2012. Vol. 262, № 1. P. 11-21. DOI: 10.1016/j.taap.2012.04.014.
- Wang Y. et al. Cloning of the ERCC2 gene and its function in malignant melanoma cells // Oncol. Lett. 2020. Vol. 20, № 2. P. 1803-1809. DOI: 10.3892/ol.2020.11708.
- Xin Y. et al. PARP1 rs1136410 Val762Ala contributes to an increased risk of overall cancer in the East Asian population: a meta-analysis // J. Int. Med. Res. 2021. Vol. 49, № 3. Art. 300060521992956. DOI: 10.1177/0300060521992956.
- Zakharenko A.L. et al. PARP1 Gene Knockout Suppresses Expression of DNA Base Excision Repair Genes // Dokl. Biochem. Biophys. 2023. Vol. 508, № 1. P. 6-11. DOI: 10.1134/S1607672922700028.
- Zhang D. et al. Association between hOGG1 polymorphism rs1052133 and gastric cancer // Oncotarget. 2017. Vol. 8, № 21. P. 34321-34329. DOI: 10.18632/oncotarget.16124.
