Бактерии-деструкторы дибутилфталата, выделенные из ризосферы мятлика лугового (Poa pratensis L.)
")
Автор: Корсакова Е.С., Пьянкова А.А., Плотникова Е.Г.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 4, 2023 года.
Бесплатный доступ
Изучена способность трех бактериальных штаммов класса Actinomycetes, выделенных из ризосферы растений мятлика лугового (Poa pratensis L.), произрастающих на территории промышленных разработок Верхнекамского месторождения солей (Пермский край), к росту на дибутилфталате (ДБФ) в качестве единственного источника углерода и энергии. На основе анализа гена 16S рРНК показано, что штамм Rh7bel проявлял сходство на уровне 100% с Rhodococcus wratislaviensis NBRC 100605T, а штаммы NKDBFbel и NKDBFgelt филогенетически близки двум типовым штаммам видов Pseudarthrobacter oxydans и Pseudarthrobacter polychromogenes (сходство 99.83%). Штаммы-деструкторы ДБФ характеризуются эффективным ростом на ключевом метаболите разложения ДБФ - ортофталевой кислоте, и утилизацией этого метаболита. Штамм Rhodococcus sp. Rh7bel демонстрировал наиболее высокие показатели утилизации ДБФ: максимальную удельную скорость потребления субстрата 0.018±0.002 ч-1, утилизацию субстрата 70.7% за 72 ч. (начальная концентрация ДБФ 0.2 г/л). Таким образом, ризосферные штаммы-деструкторы ДБФ Rhodococcus sp. Rh7bel, Pseudarthrobacter sp. NKDBFbel и NKDBFgelt являются перспективными для дальнейшего изучения и разработки технологии фиторемедиации почв, загрязненных фталатами.
Мятлик луговой, ризосфера, дибутилфталат, деструкция, rhodococcus, pseudoarthrobacter
Короткий адрес: https://sciup.org/147242768
IDR: 147242768 | УДК: 579.26: | DOI: 10.17072/1994-9952-2023-4-349-355
Dibutyl phthalate degrading bacteria isolated from the rhizosphere of bluegrass (Poa pratensis L.)
The ability of three bacterial strains of the class Actinomycetes, isolated from the rhizosphere of bluegrass (Poa pratensis L.) plants growing in the industrial development area of the Verkhnekamsk salt deposit (Perm krai), to grow on dibutyl phthalate (DBP) as the only source of carbon and energy was studied. Based on 16S rRNA gene analysis, it was shown that the strain Rh7bel showed similarity at 100% level with Rhodococcus wratislaviensis NBRC 100605T, and strains NKDBFbel and NKDBFgelt are phylogenetically close to two type strains of the species Pseudarthrobacter oxydans and Pseudarthrobacter polychromogenes (99.83% similarity). DBP-degrading strains are characterized by efficient growth on the key metabolite of DBP degradation, orthophthalic acid, and utilization of this metabolite. The strain Rhodococcus sp. Rh7bel demonstrated the highest rates of DBP utilization: the maximum specific substrate consumption rate was 0.018±0.002 h-1, substrate utilization was 70.7% in 72 hours (initial DBP concentration 0.2 g/L). Thus, rhizosphere DBP-degrading strains Rhodococcus sp. Rh7bel, Pseudarthrobacter spp. NKDBFbel and NKDBFgelt are promising for further study and development of technology for phytoremediation of soils contaminated with phthalates.
Текст научной статьи Бактерии-деструкторы дибутилфталата, выделенные из ризосферы мятлика лугового (Poa pratensis L.)
В настоящее время вследствие интенсивного производства и применения синтетических полимеров наблюдается широкое распространение эфиров фталевой кислоты (фталатов) в объектах окружающей среды. Наиболее широко используется в различных областях промышленности один из эфиров фталевой кислоты – дибутилфталат (ДБФ). Он обладает высокой растворяющей способностью и низкой вязкостью, что обусловливает его применение для изменения свойств полимеров с целью повышения их эластичности, морозостойкости. ДБФ применяется в резинотехническом производстве, при изготовлении искусственных пленок, кабельных пластикатов, эфира целлюлозы, линолеумов, лаков и искусственных смол. Фталаты и их метаболиты признаны потенциально опасными для человека и животных, т.к. обладают гепатотоксичными, тератогенными и канцерогенными свойствами [Liang et. al., 2008].
Наличие значительного количества фталатов (в том числе ДБФ) выявлено в районах интенсивной работы предприятий горнодобывающей промышленности. Так, данные соединения обнаружены в глинисто-солевых шламах, избыточных рассолах и отходах калийного производства вследствие использования в технологическом цикле обогащения калийных руд реагентов (оксиэтилированных жирных кислот, нефтепродуктов, диоксановых спиртов и др.), продуктами трансформации которых являются фталаты [Бачурин, Одинцова, 2006].
Известно, что период полураспада ДБФ в окружающей среде составляет от нескольких месяцев до 20 лет, и основную роль в процессе разложения этого соединения выполняют бактерии-деструкторы [Liang et. al., 2008]. Среди штаммов, осуществляющих аэробную деструкцию ДБФ, обнаружены бактерии различных филумов, наиболее часто встречаются представители родов Arthrobacter , Bacillus , Burkholderia , Gordonia , Pseudomonas , Rhodococcus [Liang et al., 2008; Jin et al., 2010; Stanislauskiene et al., 2011; Ka-naujiya, Sivashanmugam, Pakshirajan, 2022]. Метаболические пути разложения эфиров фталевой кислоты аэробными бактериями сочетают два процесса – первичную деградацию диэфиров фталата до моноэфиров и последующую их деструкцию до орто -фталевой кислоты (ФК), являющейся центральным метаболитом деструкции большинства фталевых эфиров. Дальнейшее разложение ФК осуществляется через образование протокатеховой кислоты (в качестве ключевого метаболита) до основных продуктов жизнедеятельности микробной клетки [Eaton, 2001; Liang et al., 2008; Vamsee-Krishna, Phale, 2008; Kasai et al,, 2019].
Фиторемедиация становится все более популярной и востребованной технологией восстановления загрязненных почв, поскольку является высокоэффективным, надежным, малозатратным и экологически чистым методом. При этом для эффективной очистки почв предполагается использование устойчивых растительно-микробных ассоциаций, содержащих в ризосфере бактерии, способные осуществлять деструкцию органических поллютантов, присутствующих в почве [Wenzel, 2009; Randika et al., 2022]. В ряде исследований показано положительное влияние высадки растений на очистку почвы от фталатов [Li et al., 2014; Liao, Nishikawa, Shih, 2019; Wu et al., 2019]. Однако исследования микроорганизмов ризосферы растений, выращенных на загрязненной фталатами почве, на сегодняшний день крайне немногочисленны [Zhang et al., 2015; Wu et al., 2019].
Цель работы – характеристика бактерий-деструкторов дибутилфталата, выделенных из ризосферы растений мятлика лугового, произрастающих в районе промышленных разработок Верхнекамского месторождения солей (Пермский край).
Материалы и методы
Объекты исследования . Из рабочей коллекции микроорганизмов Лаборатории микробиологии техногенных экосистем Института экологии и генетики микроорганизмов УрО РАН для исследования были отобраны 76 штаммов, выделенных из ризосферы растений, произрастающих на территории Верхнекамского промышленного региона (Пермский край).
Также в работе использованы два штамма NKDBFbel и NKDBFgelt, полученные из накопительной культуры при инкубировании на ДБФ в качестве субстрата. Штаммы были выделены из образца ризосферы мятлика лугового ( Poa pratensis L.), отобранного вблизи солеотвала (г. Соликамск, Пермский край, Россия).
Среды и условия культивирования . Для культивирования бактерий использовали минеральную среду Раймонда (МСР) следующего состава (г/л): NH 4 NO 3 – 2.0, MgSO 4 х 7H 2 O – 0.2, K 2 HPO 4 – 2.0, Na 2 HPO 4 - 3.0, CaCl 2 x 6H 2 O - 0.01, Na 2 CO3 - 0.1, дополненную 1%-ным раствором MnSO 4 x 2H 2 O -2 мл/л и 1%-ным раствором FeSO 4 x 7H 2 O - 1 мл/л среды [Raymond, 1961]. В качестве субстратов использовали ФК и ДБФ в концентрации 1.0 г/л. Для приготовления богатой среды Раймонда (БСР) в МСР добавляли 5 г/л триптона («VWR Life Science Amresco», США) и 2.5 г/л дрожжевого экстракта («Bi-ospringer», Франция) в качестве ростовых субстратов. Для приготовления плотных сред вносили агар («Helicon», Россия) до конечной концентрации 15 г/л.
Морфологические характеристики бактерий определяли при выращивании на агаризованной БСР. Культивирование осуществляли при температуре 28°С. Описание морфологии колоний проводили на седьмой день культивирования. Для микроскопирования клеток использовали 48-часовые культуры бактерий [Методы ..., 1983].
Секвенирование и анализ генов 16S рРНК. Определение нуклеотидных последовательностей проводили с применением набора реактивов «GenSeq-100» («Синтол», Россия) на автоматическом секвенаторе Нанофор 05 («Синтол», Россия) согласно рекомендациям производителя. Анализ полученных последовательностей осуществляли с использованием программ Sequence Scanner v. 2.0, MEGA v. 11.0 . Поиск гомологов генов 16S рРНК осуществляли по международным базам данных GenBank (http:/ и EzBioCloud .
Рост бактерий на ФК и ДБФ оценивали при культивировании в жидкой МСР. Субстрат вносили до конечной концентрации 1.0 г/л. Инокулятом служила культура, выращенная на агаризованной БСР при 28°С. Биомассу бактерий ресуспендировали в МСР, полученную суспензию бактериальных клеток (ОП 600 =1.0) добавляли в 100 мл среды в количестве 1% об./об. Культивирование штаммов бактерий осуществляли в колбах объемом 250 мл (объем среды – 100 мл) на термостатируемом шейкере Environmental Shaker Incubator ES-20/60 («BioSan», Латвия) при температуре 28°С и скорости вращения 140 об/мин. Оптическую плотность (ОП) культуральной жидкости определяли на спектрофотометре UV-Visible BioSpec-mini («Shimadzu», Япония) при длине волны 600 нм в кювете с длиной оптического пути 1 см. Расчет удельной скорости роста ( μ , ч-1) проводили по стандартной формуле
μ = (lnB2 – lnB1) / (t2 – t1), где В1 и В2 – оптические плотности культуры в моменты времени t1 и t2 соответственно [Нетрусов, 2005].
Оценку разложения ДБФ бактериями осуществляли методом газожидкостной хроматографии-масс-спектрометрии (ГЖХ-МС). Штаммы выращивали в среде МСР, содержащей дибутилфталат (1.0 г/л), клетки собирали центрифугированием (9 000 об/мин, 5 мин.) и отмывали при помощи МСР. Отмытые дважды клетки (ОП 600 =2.0) инокулировали в 1 мл МСР с ДБФ (0.2 г/л) и инкубировали при 28°С в течение трех суток при аэрации на роторном шейкере (150 об/мин). Экстракцию ДБФ осуществляли равным объемом гексана в течение 120 мин. на шейкере при 100 об/мин. Остаточную воду удаляли из образцов путем введения безводного сульфата натрия. Анализ проводили на газовом хроматографе-масс-спектрометре «Agilent» 7890B модель G3440B («Agilent», США), с кварцевой колонкой RESTEK RTx-5MS («Restek», США). Анализ хроматограмм проводили с использованием программы MassHunter Qualitative Analysis 10.0 («Agilent», США.). Идентификацию ДБФ проводили путем сравнения времени выхода пиков со временем выхода пиков контрольного соединения, а также по хромато-масс спектрам.
Расчет удельной скорости утилизации ( μ , сут-1) проводили по стандартной формуле
μ = (lnС1 – lnC2) / (t2 – t1), где С1, С2 – концентрация субстрата в начальный и конечный моменты времени t1 и t2 соответственно [Нетрусов, 2005].
Статистическая обработка результатов . Все эксперименты были выполнены в трехкратной повторности. Полученные данные обрабатывали с использованием программы Microsoft Excel 2007.
Результаты и их обсуждение
В ходе проведенной работы 76 штаммов, выделенные из образцов ризосферы растений с территории солеразработок (г. Березники, г. Соликамск, Пермский край), были исследованы на способность к росту на ДБФ и ФК – ключевом метаболите бактериального разложения ДБФ. В результате из 14 активных деструкторов ФК только один штамм (Rh7bel), изолированный из ризосферы мятлика лугового (Poa pratensis L.), был способен к росту на дибутилфталате как единственном источнике углерода и энергии. При культивировании на агаризованной БСР штамм Rh7bel формировал округлые непрозрачные колонии диаметром 1.5–2.5 мм с ровным краем, гладкой матовой поверхностью, выпуклым профилем, мелкозернистой структурой и мягкой консистенцией. Клетки штамма грамположительные, неподвижные, в стационарной фазе роста представлены длинными палочками. На основе анализа фрагмента гена 16S рРНК (808 п.н.) было установлено, что штамм Rh7bel проявлял сходство на уровне 100% с Rhodococcus wratislaviensis NBRC 100605T.
Штаммы-деструкторы дибутилфталата NKDBFbel и NKDBFgelt имели округлые полупрозрачные колонии диаметром 2–4 мм с ровным краем, гладкой блестящей поверхностью, выпуклым профилем, однородной структурой и мягкой консистенцией. Штамм NKDBFbel на агаризованной БСР формировал колонии белого цвета, а штамм NKDBFgelt – желтого цвета. Клетки обоих штаммов грамположительные, неподвижные, в стационарной фазе представлены утолщенными короткими палочками. На основе анализа фрагмента гена 16S рРНК штаммы NKDBFbel и NKDBFgelt по гену 16S рРНК (594 и 602 п.н., соответственно) имели сходство на уровне 99.83% с двумя типовыми штаммами – Pseudarthrobacter oxydans DSM 20119T и Pseudarthrobacter polychromogenes DSM 20136T.
Исследованы ростовые характеристики штаммов Rh7bel, NKDBFbel и NKDBFgelt при росте на ДБФ и ФК в качестве субстратов (рис. 1, 2; таблица). Штаммы имели высокие показатели максимальной ОП при росте на ФК (>1.0), но только у штамма NKDBFgelt были зарегистрированы сравнимые значения ОП при росте на ДБФ (таблица). Максимальные значения удельной скорости роста культуры выявлены у штамма NKDBFgelt при росте на обоих субстратах. Штамм Rhodococcus sp. Rh7bel демонстрировал высокие показатели при росте на ФК, однако в процессе инкубирования на ДБФ клетки штамма формировали конгломераты, что могло повлиять на регистрацию низких значений параметров роста культуры (рис. 1, 2; таблица).
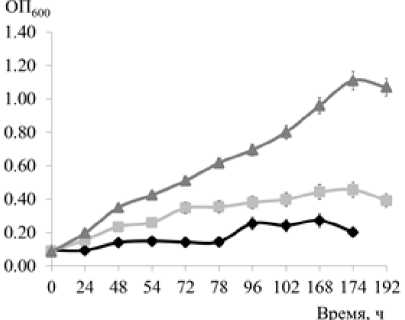

О 24 48 54 72 78 96 102 168 174
Параметры роста штаммов Rh7bel, NKDBFbel, NKDBFgelt на ДБФ и утилизации ДБФ [Growth parameters of strains Rh7bel, NKDBFbel, NKDBFgelt and DBP utilization]
|
Параметры роста |
Rhodococcus sp. Rh7bel |
Pseudarthrobacter sp. NKDBFbel |
Pseudarthrobacter sp. NKDBFgelt |
|||
|
Ростовой субстрат, 1.0 г/л |
ФК |
ДБФ |
ФК |
ДБФ |
ФК |
ДБФ |
|
Удельная скорость роста, ч-1 |
0.041± 0.003 |
0.002± 0.001 |
0.008± 0.001 |
0.003± 0.001 |
0.054± 0.003 |
0.016± 0.002 |
|
Максимальное значение ОП 600 |
1.10 |
0.27 |
1.04 |
0.46 |
1.25 |
1.11 |
|
Лаг-фаза роста, ч |
24 |
78 |
78 |
24 |
54 |
24 |
|
Удельная скорость утилизации, ч-1 |
н.о. |
0.018± 0.002 |
н.о. |
0.008± 0.002 |
н.о. |
0.009± 0.003 |
|
Утилизация, % |
н.о. |
70.7±0.4 |
н.о. |
43.3±0.3 |
н.о. |
45.1±0.3 |
Примечание: н.о. – не определяли.
Время, ч
-♦■Rh7bcl 1 NKDBFbel *NKDBFgelt ♦Rh7bel 4 NKDBFbel *NKDBFgelt
Л /;
Рис. 1. Рост штаммов-деструкторов на ФК ( A ) и ДБФ ( B ) (1.0 г/л) в МСР
[Growth of destructor strains on FA ( A ) and DBP ( B ) (1.0 g/L) in MMR]
С использованием метода ГЖХ-МС исследована способность штаммов к утилизации ДБФ (рис. 2). Наиболее высокие показатели утилизации субстрата (70.7% за 72 ч.) и максимальная удельная скорость потребления дибутилфталата (0.018±0.002 ч-1) зафиксированы у штамма Rhodococcus sp. Rh7bel. Два других штамма-деструктора рода Pseudarthrobacter показали снижение количества ДБФ лишь на 43–45% за 72 ч. эксперимента.
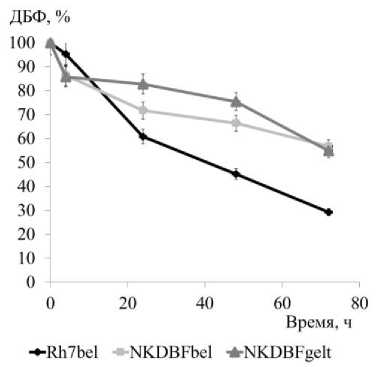
Рис. 2. Утилизация ДБФ (0.2 г/л) исследуемыми штаммами
[Utilization of DBP (0.2 g/L) by the studied strains]
На основании известных путей деструкции эфиров фталевых кислот бактериями рода Rhodococcus и Pseudarthrobacter можно предположить, что разложение дибутилфталата исследуемыми штаммами осуществляется до монобутилфталата, затем до образования ключевых метаболитов – фталевой кислоты и протокатеховой кислоты, которые далее метаболизируются в цикле трикарбоновых кислот [Choi et al., 2005; Jin et al., 2010; Chen et al., 2021].
Заключение
В результате проведенных исследований установлено, что штаммы-деструкторы Rhodococcus sp. Rh7bel, Pseudarthrobacter sp. NKDBFbel и NKDBFgelt, выделенные из ризосферы растений мятлика лугового ( Poa pratensis L.), произрастающих на почвах, загрязненных отходами калийного производства, способны к эффективному росту на среде с дибутилфталатом в качестве субстрата. Активность штаммов-деструкторов ДБФ подтверждена ростом на ключевом метаболите разложения ДБФ – орто фталевой кислоте, и эффективной утилизацией этого метаболита. Штамм Rhodococcus sp. Rh7bel имел наиболее высокое значение утилизации ДБФ (70.7% за 72 ч., при начальной концентрации субстрата 0.2 г/л) и максимальную удельную скорость потребления ДБФ (0.018±0.002 ч-1). Таким образом, ризосферные штаммы-деструкторы ДБФ Rhodococcus sp. Rh7bel, Pseudarthrobacter sp. NKDBFbel и NKDBFgelt являются перспективными для дальнейшего изучения и разработки технологии фиторемедиации почв, загрязненных фталатами.
Список литературы Бактерии-деструкторы дибутилфталата, выделенные из ризосферы мятлика лугового (Poa pratensis L.)
- Бачурин Б.А., Одинцова Т.А. Стойкие органические загрязнители в отходах горного производства // Современные экологические проблемы Севера. Апатиты: Изд-во Кольского НЦ РАН. 2006. Ч. 2. С. 7–9.
- Методы общей бактериологии / под ред. Ф. Герхардта и др. М.: Мир, 1983. Т. 1–3.
- Нетрусов А.И. Практикум по микробиологии. М.: Академия, 2005. 608 с.
- Chen F. et al. High-efficiency degradation of phthalic acid esters (PAEs) by Pseudarthrobacter defluvii E5: performance, degradative pathway, and key genes // Sci. Total Environ. 2021. Vol. 794. 148719.
- Choi K.Y. et al. Molecular and biochemical analysis of phthalate and terephthalate degradation by Rho-dococcus sp. strain DK17 // FEMS Microbiol. Letters. 2005. Vol. 252. P. 207–213.
- Eaton R.W. Plasmid-encoded phthalate catabolic pathway in Arthrobacter keyseri 12B // J. Bacteriol. 2001. Vol. 183. P. 3689–3703.
- Jin D.-C. et al. Biodegradation of di-n-butyl phthalate by Rhodococcus sp. JDC-11 and molecular detec-tion of 3,4-phthalate dioxygenase gene // J. Microbiol. Biotechnol. 2010. Vol. 20(10). P. 1440–1445.
- Kanaujiya D.K., Sivashanmugam S., Pakshirajan K. Biodegradation and toxicity removal of phthalate mixture by Gordonia sp. in a continuous stirred tank bioreactor system // Environmental Technology & Innova-tion. 2022. Vol. 26. 102324.
- Kasai D. et al. 2,3-dihydroxybenzoate meta-cleavage pathway is involved in o-phthalate utilization in Pseudomonas sp. strain PTH10 // Scientific Reports. 2019. Vol. 9. 1253.
- Li Y.W. et al. Plant uptake and enhanced dissipation of di(2-ethylhexyl) phthalate (DEHP) in spiked soils by different plant species // International Journal of Phytoremediation. 2014. Vol. 16. P. 609–620.
- Liang D.-W. et al. Phthalates biodegradation in the environment // Appl. Microbiol. Biotechnol. 2008. Vol. 80. P. 183–198.
- Liao C.S., Nishikawa Y., Shih Y.T. Characterization of di-n-butyl phthalate phytoremediation by garden lettuce (Lactuca sativa L. var. longifolia) through kinetics and proteome analysis // Sustainability. 2019. Vol. 11. P. 1–16.
- Randika J.L.P.C. et al. Bioremediation of pesticidecontaminated soil: a review on indispensable role of soil bacteria // The Journal of Agricultural Sciences – Sri Lanka. 2022. Vol. 17(1). P. 19–43.
- Raymond R.L. Microbial oxidation of n-paraffinic hydrocarbons // Develop. Ind. Microbiol. 1961. Vol. 2(1). P. 23–32.
- Stanislauskiene R. et al. Analysis of phthalate degradation operon from Arthrobacter sp. 68b // Biologi-ja. 2011. Vol. 57(3). P. 45–54.
- Vamsee-Krishna C., Phale P.S. Bacterial degradation of phthalate isomers and their esters // Indian J. Microbiol. 2008. Vol. 48. P. 19–34.
- Wenzel W.W. Rhizosphere processes and management in plant-assisted bioremediation (phytoremedia-tion) of soils // Plant & Soil. 2009. Vol. 321. P. 385–408.
- Wu K. et al. Responses of soil microbial community and enzymes during plant-assisted biodegradation of di-(2-ethylhexyl) phthalate and pyrene // International Journal of Phytoremediation. 2019. Vol. 21(7). P. 683–692.
- Zhang Y. et al. Effect of di-n-butyl phthalate on root physiology and rhizosphere microbial community of cucumber seedlings // Journal of Hazardous Materials. 2015. Vol. 289. P. 9–17.
