Бактерии - деструкторы стойких органических загрязнителей - эфиров фталевой кислоты как основа для создания новых экобиотехнологий

Автор: Корсакова Екатерина Сергеевна, Пьянкова Анна Александровна, Плотникова Елена Генриховна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биотехнология
Статья в выпуске: 3-5 т.15, 2013 года.
Бесплатный доступ
В данной статье рассмотрена проблема микробиологического разложения экологически опасных поллютантов на примере дибутилфталата - как одного из распространенных загрязнителей, в том числе в отходах калийного производства. Из образцов техногенно-минеральных образований, почвы и грунта территории соледобывающего предприятия ОАО «Уралкалий» (г. Березники, г. Соликамск, Пермский край) выделены активные галотолерантные штаммы-деструкторы дибутилфталата, перспективные для создания биотехнологии на основе микробной деструкции экотоксикантов. Проведен филогенетический анализ изолированных бактерий, на основании которого штаммы-деструкторы были отнесены к родам Rhodococcus, Arthrobacter, Dietzia, Bacillus, Exiguobacterium, Halomonas и Pseudomonas.
Орто-фталевая кислота, дибутилфталат, бактерии-деструкторы, ген 16s ррнк
Короткий адрес: https://sciup.org/148202060
IDR: 148202060 | УДК: 579.66:661.8…784
Bacteria-destructors of persistent organic pollutants - phthalic acid esters, as a basis for the creation of new ecobiotechnology
In this article we analyzed the problem of microbial degradation of environmentally dangerous pollutants by the example of dibutyl phthalate as one of the most common pollutants, including waste potash production. From the samples of technogenic mineral formations, soil and ground of a salt-mining plant JSC "Uralkali" (Berezniki, Solikamsk, Perm krai) there were isolated active halotolerant strains-destructors of dibutyl phthalate being promising for creation a biotechnology based on microbial degradation of toxicants. Following a phylogenetic analysis of the isolated bacteria the strains-destructors were assigned to the genera Rhodococcus, Arthrobacter, Dietzia, Bacillus, Exiguobacterium, Halomonas and Pseudomonas.
Текст научной статьи Бактерии - деструкторы стойких органических загрязнителей - эфиров фталевой кислоты как основа для создания новых экобиотехнологий
Перспективной методологией повышения уровня устойчивости растений к стрессам, вызванным водным дефицитом, является генетическая инженерия, одно из направлений которой связано с манипуляцией генами, принимающими участие в регуляции метаболизма L-пролина (Pro). Эта свободная аминокислота рассматривается, как осмолит, связанный с толерантностью к стрессам, и как фактор, имеющий отношение к процессам развития растений [1, 2].
Эндогенное содержание L-пролина в растительных клетках в норме и при стрессе координировано регулируется его синтезом, катаболизмом и транспортом [1]. Для повышения уровня накопления Pro главным образом исследуются возможности использования генов, контролирующих синтез белков скорость-лимитирующих энзимов его синтеза (пирролин-5-карбоксилатсинтетазу) и катаболизма (пролиндегидрогеназу, ProDH). Что касается генов ProDH , то частичную их супрессию можно осуществлять с применением векторных конструкций, созданных на основе чужеродных генов ProDH , расположенных в антисмысловой ориентации или в форме обращённого повтора. Считается, что использование siRNA –технологий является более эффективным [3, 4].
Гены ProDH клонированы для небольшого числа видов растений, их экспрессия преимущественно регулируется на транскрипционном уровне де- и регидратацией [1, 5].
осмотолерантность, двухцепочечный РНК-супрессор
Роль генов ProDH в процессах адапта-ции/устойчивости растений к разным абиотическим стрессам изучена у нескольких видов растений, при этом результаты исследований были не всегда однозначными. В связи с этим при разработке молекулярных биотехнологий очевидна целесообразность предварительного изучения их эффективности для повышения уровня стресс-устойчивости культурных растений. Что касается подсолнечника, то информация о генах катаболизма Pro и изменениях уровня их экспрессии методом генетической инженерии отсутствует. Цель данной работы состояла в анализе перспективности применения двухцепочечного РНК-супрессора гена пролиндегидрогеназы, созданного на основе гена ProDH1 арабидопсиса, для повышения уровня толерантности растений подсолнечника к водному дефициту и засолению.
Объектом исследования служил подсолнечник сорта Прометей (селекции Института масличных культур УААН) и инбредной линии 96А/3 (селекции Одесского селекционно-генетического Института УААН). Индукцию побегообразования in vitro из инокулированных эксплантов, а также интеграцию рекомбинантной молекулы ДНК в геном подсолнечника оценивали ПЦР-методом по наличию экзона, интрона гена ProDH 1 арабидопсиса и селективного npt II-гена так, как описано ранее [6, 7].
Agrobacterium -опосредованную трансформацию осуществляли с использованием штамма LBA 4404, содержащиего бинарный вектор pBi2E с двухцепочечным РНК-супрессором гена пролиндегидрогеназы, полученным на основе гена арабидопсиса ProDH1 (рис. 1).
RB
LB
|
NOSpro |
npt II |
NOSterm |
35Spro |
ProDH-ex1 |
int1 |
ProDH-ex1 |
NOSterm |
Рис. 1. Блок-схема Т-ДНК области pBi2E. NOSpro и 35Spro – соответственно промоторы гена нопалинсинтазы и 35S вируса мозаики цветной капусты; ProDH-ex1 и int1 – фрагменты первого экзона и интрона гена ProDH1 арабидопсиса, соответственно; NOSterm – сигнал полиаденилирования гена нопалинсинтазы; npt II – ген неомицинфосфотрансферазы E. coli, RB, LB – повторы, ограничивающие Т-область
Уровень свободного L-пролина определяли методом Чинарда с модификациями [8]. Для сравнительного анализа использовали регенеранты одного и того же возраста. Пробы отбирали и фиксировали в одно и то же время суток. Стрессовые условия создавали добавлением в среду культивирования маннита (0.4М) или солей морской воды (2,0%). Динамику изменения содержания свободного Pro в условиях стресса и в период восстановления после водного дефицита и сульфатно-хлоридного засоления определяли в листьях 1-го и 2-го ярусов побегов индивидуальных трансформантов. Контролем служило содержание L-пролина в листьях 7-суточных нетрансформированных регенерантов, культивируемых в условиях стресса. Статистическую обработку проводили по Стьюденту.
Анализ перспективности частичной супрессии генов пролиндегидрогеназы подсолнечника для по- вышения уровня его осмотолерантности проводили в условиях in vitro, используя трансгенные регенеранты. Частота трансформации растений подсолнечника сорта Прометей и инбредной линии 96А/3, содержащих копию Т-ДНК, соответственно составляла 16,0% и 8,3%. Такие варианты рассматривались нами как предполагаемые трансформанты (Т0).
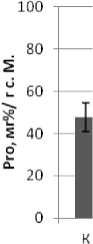
Предыдущие наши исследования показали, что индукция побегообразования подсолнечника на примере сорта Прометей осуществляется только из эксплантов, характеризующихся повышенным уровнем свободного L-пролина [8]. Это позволяет рассматривать данную аминокислоту как один из компонентов, участвующих в процессе органогенеза подсолнечника. В связи с этим, проводили сравнительные исследования содержания Pro в листьях Т0-побегов и трансформированных вариантов, в геном которых не был интегрирован трансген (рис. 2).
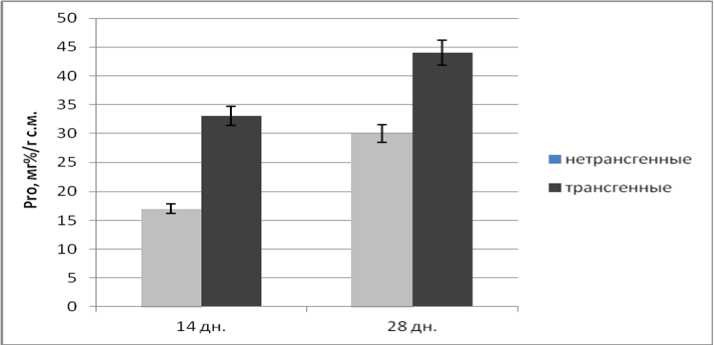
Рис. 2. Содержание свободного пролина в трансгенных и нетрансгенных растениях-регенерантах
Видно, что в нормальных условиях культивирования для трансгенных Km- устойчивых побегов в ходе их развития (14- и 28-суточные проростки) было характерно превышение (относительно контроля) уровня аккумуляции L-пролина, что возможно в случае уменьшения уровня транскрипции эндогенных генов пролиндегидрогеназы подсолнечника. Степень увеличения содержания пролина близка к показателю у трансформантов табака, несущих такую же конструкцию [2].
Известно, что ответная реакция растений на умеренные дозы стрессора сопровождается активацией транскрипции гена P5CS и снижением количества мРНК ProPDH, уровень которой возрастает в период регидратации, то есть, после накопления Pro осуществляется его снижение во время восстановления после стресса. На первый план выступает сопряжённость событий: осмоустойчивость - метаболизм свободного L-пролина, координировано регулируемая генами его синтеза и катаболизма.
Поскольку в работе Mani S. и соавт. [5] отмечено отсутствие корреляции между экспрессией гена ProPDH1 арабидопсиса, на основе которого создана используемая нами векторная конструкция, и осмоустойчивостью этого вида, анализ эффективности частичной супрессии гена пролиндегидрогеназы подсолнечника проводили при летальных дозах стрессоров, моделирующих in vitro водный де- фицит и сульфатно-хлоридное засоление. В условиях, когда происходит гибель всех контрольных растений, жизнеспособность предполагаемых трансформантов, сопряжённая с повышением уровня свободного L-пролина и его снижением в отсутствии стрессора, может осуществляться только при наличии функционального трансгена.
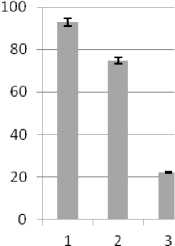
В связи с тем, что осмотический стресс имеет две разновидности - засоление и водный дефицит, имеющие свои особенности, рассматривали их действие и ответные реакции со стороны трансформантов раздельно. Изучение динамики накопле-ния/убыли Pro подтвердило корректность такого подхода. На рис. 3 представлены результаты определения содержания свободного Pro в листьях трансгенных регенерантов подсолнечника инбред-ной линии 96А/3 и сорта Прометей, выдержавших селективное давление. Изменения в содержании пролина трансгенных вариантов оценивали относительно 7-суточного контроля, культивируемого в стрессовых условиях. Побеги контрольной группы гибли к концу второй недели.
С увеличением продолжительности моделированного водного дефицита содержание свободного Pro в клетках жизнеспособных трансформантов обоих изучаемых генотипов подсолнечника многократно возрастало аналогичным образом в течение короткого периода, тогда как в условиях восстановления после стресса уровень аминокислоты резко снижался (рис. 3, А, Б). При культивировании трансформантов в условиях летальных доз засоления также имело место увеличение уровня Pro и его снижение при возвращении к нормальным условиям (рис. 4, В, Г). В то же время наблюдались различия в ответной реакции на стрессовое воздействие у индивидуальных трансформантов. Тем не менее, система метаболизма свободного Pro у обоих генотипов, безусловно, активно функционировала. Одна из причин наблюдаемых различий могла быть обусловлена дифференциальной экспрессией генов катаболизма - синтеза, что, в частности, установлено для люцерны [9].
А
Б

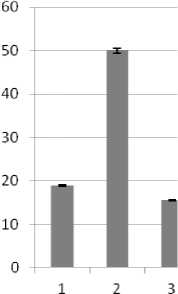

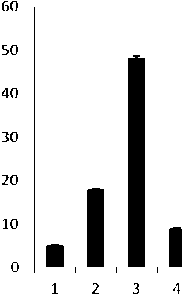
Рис. 3 . Уровень свободного L-пролина в листьях трансформантов подсолнечника сорта Прометей (А) и ин-бредной линии 96-А/3 (Б) в условиях водного дефицита. К - контроль, нетрансгенные регенеранты. 1, 2 (А) и 1, 2, 3 (Б) индивидуальные трансформанты, культивируемые в течение 7, 14, 21 сут при стрессе, соответственно. 3 (А) и 4(Б) - трансформанты, культивируемые в период восстановления после стресса (21-е и 28-е сут, соответственно)
В
Г
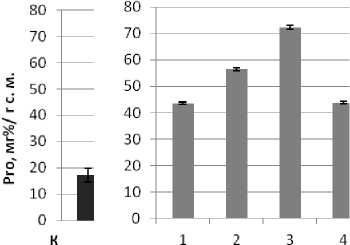
Рис. 4. Уровень свободного L-пролина в листьях трансформантов подсолнечника сорта Прометей (В) и инбред-ной линии 96-А/3 (Г) в условиях в сульфатно-хлоридного засоления. К - контроль, нетрансгенные регенеранты.
Таким образом, стресс-устойчивость трансформантов к водному дефициту и сульфатно-хлоридному засолению сопровождалась повышением уровня свободного L-пролина и его снижением в период восстановления, что свидетельствует, во-первых, о функциональной значимости этой системы для данного объекта, и, во-вторых, о возможности использования частичной супрессии гена пролиндегидрогеназы подсолнечника для получения стрессоустойчивых форм.
Работа поддержана грантом совместных научных проектов НАН Украины (16-05-2012) – СО РАН (№ 11).
-
1. Szabados L, Savoure A. Proline: a multifunctional amino acid // Trends in Plant Science. 2009. V. 15. N 2. P. 89-97.
-
2. Ибрагимова С.С.., Колодяжная Я.С., Герасимова С.В., Кочетов А.В. Частичная супрессия гена пролиндегидрогеназы увеличивает устойчивость растений к различным видам абиотических стрессов // Физиология растений. 2012. Т. 59. С. 99-107.
-
3. Tateishi Y., Nakagama T., Esaka M. Osmotolerance and growth stimulation of transgenic tobacco cells accumulating free proline by dehydrogenase expression with double-
stranded RNA interference technique // Physiologia Plantarum. 2005. V. 125. P. 1399-3054.
-
4. Титов С.Е. Изучение генетически модифицированных растений табака ( Nicotiana tabacum L.), экспрессирующих антисмысловой супрессор гена пролиндегидрогеназы: Автореф. дис. …канд. биол. наук. Новосибирск, 2008. 18 с.
-
5. Mani S., Van de Cotte B, Van Montagu M., Verbruggen N. Altered levels of proline dehydrogenase cause hypersensitivity to proline and its analogs in Arabidopsis // Plant Physiology. 2002. V. 128. P. 73-83.
-
6. КомисаренкоА.Г., Михальская С.И., Кочетов А.В., Тищенко Е.Н. Индукция регенерации in vitro при Agrobacterium –опосредованной трансформации инбредных линий подсолнечника // Біотехнологія. 2010. Т. 3. № 4. С. 67-74.
-
7. Михальская С.И., Адаменко Н.И., Моргун Б.В . и др . Компетентность к Agrobacterium –опосредованной трансформации сегментов побега элитных инбредных линий кукурузы // Біотехнологія. 2012. Т. 5. № 3. С. 98-103.
-
8. Сергеева Л.Е., Комисаренко А.Г., Бронникова Л.И. и др. Содержание свободного пролина в тканях подсолнечника при реализации морфогенетического потенциала in vitro // Біотехнологія. 2013. Т. 6. № 1. С. 113-118.
-
9. Miller G., Stein H., Honig A. et al. Responsive modes of Medicago sativa proline dehydrogenase genes during salt stress and recovery dictate proline accumulation // Planta. 2005. V. 222. N 1. P. 70-79.
THE ANALYSIS OF EFFECTIVENESS OF DOUBLE-STRANDED RNA-SUPPRESSOR OF PROLINE DEHYDROGENASE GENE FOR ELEVATION OF SUNFLOWER ( HELIANTHUS ANNUUS L.) TOLERANCE TO WATER DEFICIT AND SALINITY ©2013 А.G. Komisarenko1, S.I. Mykhalska1, L.E. Sergeeva1, A.V. Kochetov2, E.N. Tishchenko1
1Institute of Plant Physiology and Genetics of National Academy of Sciences of Ukraine, Kiev 2Institute of Cytology and Genetics, Russian Academy of Sciences, Novosibirsk
Perspective of using of strain LВА4404 harboring plasmid pBi2E with double-stranded RNA-suppressor, which was prepared on basis of Arabidopsis ProDH1 gene, for the elevation of sunflower ( Helianthus annuus L.) tolerance to water deficit and salinity was estimated. The transgenic regenerants that maintained stress pressure of lethal doses of mannitol (0.4M) or sea water salts (2,0%) demonstrated the elevated level of free proline under stress conditions and its decline under recovery period. These data declare that ProDH gene confer the osmotolerance of sunflower.
Список литературы Бактерии - деструкторы стойких органических загрязнителей - эфиров фталевой кислоты как основа для создания новых экобиотехнологий
- URL: http://chm.pops.int/
- Барштейн Р.С., Кирилович В.И., Носовский Ю.Е. Пластификаторы для полимеров. М.: Химия, 1982. 197 с.
- Liang D.-W., Zhang T., Fang H. Phthalates biodegradation in the environment//Appl. Microbiol. Biotechnol. 2008. V. 80. P. 183-198.
- Бачурин Б.А., Одинцова Т.А. Стойкие органические загрязнители в отходах горного производства//Современные экологические проблемы Севера. Апатиты: Издво Кольского НЦ РАН, 2006. Ч. 2. С. 7-9.
- Jin D.-C., Liang R.-X. Biodegradation of Di-n-Butyl Phthalate by Rhodococcus sp. JDC-11 and Molecular Detection of 3,4-Phthalate Dioxygenase Gene//Microbiol. Biotechnol. 2010. V. 20. N 10. P. 1440-1445.
- Егорова Д.О., Корсакова Е.С., Демаков В.А., Плотникова Е.Г. Деструкция ароматических углеводородов штаммом Rhodococcus wratislaviensis КТ112-7, выделенным из отходов соледобывающего предприятия//Прикладная биохимия и микробиология. 2013. Т. 49. № 3. С. 267-278.
- URL: http://www.ncbi.nlm.nih.gov/
- URL: http://www.eztaxon.org/
