Бактериоцины молочнокислых бактерий

Автор: Сультимова Т.Д., Захаров Е.В.
Журнал: Вестник Восточно-Сибирского государственного университета технологий и управления @vestnik-esstu
Рубрика: Биологические науки
Статья в выпуске: 2 (59), 2016 года.
Бесплатный доступ
Антимикробное действие молочнокислых бактерий люди использовали издавна для увеличения срока годности пищевых продуктов за счет образования молочной кислоты с сопровождающимся понижением уровня рН. В настоящее время доказано, что некоторые штаммы молочнокислых бактерий образуют так называемые бактериоцины - антимикробные вещества белковой природы. Синтез бактериоцинов - наследственная особенность микроорганизмов, проявляющаяся в том, что каждый штамм способен образовывать один или несколько определенных, строго специфичных для него антибиотических веществ. В настоящее время возрастает интерес к бактериоцинобразующим лактококкам рода Lactococcus lactis, которые вследствие своей безвредности, высокой ферментативной и антимикробной активности являются объектом фундаментальных исследований по созданию новых активных пробиотиков и биологических консервантов. В настоящее время предъявляются высокие требования к качеству продуктов питания и их безопасности для здоровья, поскольку широко используемые химические консерванты, увеличивающие срок хранения продуктов питания, вызывают опасения у потребителей.
Молочнокислые бактерии, бактериоцин, низин, низинпродуцирующий штамм
Короткий адрес: https://sciup.org/142148241
IDR: 142148241 | УДК: 577.18.05:577.182.44
Bacteriocins of lactic acid bacteria
The antimicrobial activity of lactic acid bacteria has been used by people long since to increase the storage of food products due to the formation of lactic acid with an accompanying decrease in pH level. The synthesis of bacteriocins - inherited trait of microorganisms, which is manifested in the fact that each strain is capable of forming one or several certain antibiotic substances, strictly specific to it. Now interest in forming bacteriocins by Lactococcus lactis increases. The bacteriocins are the subject of the basic research on the development of new active probiotics and biological preservatives owing to their safety, high enzymatic and antimicrobial activity. One of the main aspects of this interest is the increased consumer demand for quality food products and their safety for human health as widely used chemical preservatives cause concern among population.
Текст научной статьи Бактериоцины молочнокислых бактерий
Антимикробный эффект молочнокислых бактерий издавна использовался для продления срока годности пищевых продуктов [1]. Классические работы Л. Пастера открыли первые страницы в изучении этой группы микроорганизмов [2]. По мнению ряда исследователей, образование органических кислот (молочной и уксусной) из углеводов приводит к снижению рН среды и предотвращает развитие других микроорганизмов [3]. Кроме того, бактерицидным действием на специфические группы микроорганизмов, включая патогенные формы, развивающиеся в продуктах питания в процессе хранения и выделяющие энтеротоксины, обладают белковые вещества, выделяемые некоторыми видами бактерий - бактерио-цинов [4].
Бактериоцины представляют собой комплекс пептидов с молекулярной массой от 2 до 35 кДа, существенно отличающихся друг от друга по физико-химическим характеристикам и биологическим эффектам [6, 7]. Выявлено, что на проявление антагонистической активности бактериоцинов, продуцируемых молочнокислыми бактериями, влияют температура, электрическое поле, рН, состав, консистенция среды, присутствие Ca2+ и Mg2+ и другие факторы [7, 8, 9].
К числу общих свойств бактериоцинов относится их чувствительность к температуре, хотя это свойство также может варьировать в широких пределах. Некоторые разрушаются при температуре 48-50 ° С, другие кратковременно выдерживают температуру 60-70 ° С, а отдельные сохраняют активность даже при 100 ° С. Низин выдерживает кипячение до 120 ° С без потери активности.
Бактериоцины чувствительны к воздействию протеаз и имеют различный молекулярный вес (30000-100000 Да). Белок бактериоцина связан с липополисахаридом клеточной оболочки, но только белковая часть молекулы обладает антибактериальной активностью [11, 12].
На основании физико-химических свойств, аминокислотного состава, способов выведения, а также антимикробного спектра действия бактериоцины энтерококков различные исследователи разделяют на три класса: I, II, III [8, 13, 14].
Бактериоцины I класса
К первому классу бактериоцинов относятся лантибиотики, низкомолекулярные катионные, гидрофобные, устойчивые к нагреванию пептиды, в состав которых входят такие редкие тиоэфирные аминокислоты, как лантионин и Р —метиллантионин, дегидроаланин, дегидробутирин, и несколько тиоэфирных мостиков, которые замещают дисульфидные связи и придают этим белковым соединениям некоторую полициклическую структуру [6].
К этой группе относятся низины, образуемые Lactococcus (Streptococcus) lactis , эпидермин, галлидермин, продуцентом которого является St. gallinarum , а также субтилин (B. subtilis).
Примером первого класса бактериоцинов является низин - лантибиотик, секретируемый бактериями Lactococcus lactis . Содержит 34 остатка аминокислот (из них 30 остатков кислот (Lis, His, Asp, Ser, Pro, Gli, Ala, Val, Met, Ile, Leu), а также остатки редко встречающихся серосодержащих аминокислот: лантионина, р —метиллантионина и ненасыщенных аминокислот дегидроаланина и Р —метилдегидроаланина. Низин обладает бактерицидным действием на грамположительные микроорганизмы: стрептококки различных серологических групп, стафилококки, пневмококки, микобактерии, споровые - аэробные и анаэробные бактерии. Обладая способностью адсорбироваться на поверхности спор чувствительных к нему спорообразующих микроорганизмов, низин нарушает проницаемость спор, снижает устойчивость спор к воздействию высоких температур. Из аэробных споровых бактерий рода Bacillus к низину наиболее чувствительны Bac. stearothermophilus, Bac. coagulans, Bac. cereus , вызызвающие порчу молочных продуктов. Также чувствительны к действию низина споровые анаэробы рода Clostridium (Budde, Rasch, 2001) .
Бактериоцины II класса
Большинство бактериоцинов относится ко II классу. Представляют собой низкомолекулярные катионные термостабильные пептиды, сохраняющие свою активность при широком диапазоне рН (3–9), слабо иммуногенные и нетоксичные для человека и животных [6, 20, 21]. Они вызывают повреждение цитоплазматической мембраны грамположительных бактерий, а по современным данным - и грамотрицательных бактерий [10, 21, 22, 23]. Бактериоцины II класса чаще всего синтезируются как препептиды, а затем без модификации их аминокислотных остатков, в процессе секреции из клетки продуцентов утрачивают свои лидирующие компоненты. Гены, необходимые для продукции бактериоцинов II класса (регуляторные, гены, кодирующие препептид (или пептид), обеспечивающие транспорт через мембрану и устойчивость бактерий к собственному антимикробному пептиду), располагаются в плазмидах или в хромосоме. Кроме того, гены бактериоцинов могут быть обнаружены в мобильных транспозоноподобных генетических модулях ДНК-содержащих бактериофагов [20].
Бактериоцины II класса по химическому строению, особенностям выведения из клетки продуцента, спектру антимикробной активности разделяют на 3 подкласса: IIa, IIb, IIc [11, 20, 25].
Бактериоцины подкласса IIа
Бактериоцины подкласса IIa наиболее хорошо изучены. Чаще всего их называют пе-диоциноподобными пептидами. Во время экспорта через клеточную мембрану от молекулы их препептидов отрезается N-терминальная лидирующая последовательность с двумя глициновыми аминокислотными остатками на конце [26]. После секреции из клетки эти бактерио-цины представляют собой пептиды с молекулярной массой до 10 кДа, образующие, как правило, один дисульфидный мостик. После попадания в окружающую среду бактериоцины данного подкласса атакуют клетку-мишень за счет электростатического гидрофобного взаимодействия с последующим проникновением внутрь чувствительной клетки. Для специфического распознавания бактериоцином клетки-мишени необходимо присутствие на ее клеточной мембране маннозопротеиновых рецепторов, являющихся частью фосфотрансфераз-ной системы. В то же время нельзя исключить, что подобное взаимодействие является необязательным условием для проявления антагонистической активности к бактериям, в том числе и рода Listeria [22]. При помощи C-части пептиды проникают в мембрану и действуют на клетку-мишень, что определяет антибактериальную активность и специфичность бактерио-цина. Показано, что степень проникновения зависит от заряда липосом, а также от расположения аминокислотных остатков в С-части молекулы бактериоцина. Бактериоцины IIa типа формируют ионоселективные поры в клетках-мишенях, вызывая выход из последних ионов калия, аминокислот и других низкомолекулярных веществ. Это приводит к изменению ионного баланса, уровня рН в клетке и последующей быстрой потере внутриклеточной АТФ [26].
Бактериоцины подкласса IIb
Бактериоцины этого подкласса для наличия высокой антибактериальной активности должны состоять из двух различных субъединиц или пептидов, действующих только вместе или раздельно как синергисты [10, 11, 20]. До настоящего времени, несмотря на способность проявлять антибактериальную активность самостоятельно и расположение в различных генах, эти вещества не были выделены.
Бактериоцины подкласса IIс
К подклассу IIc относятся однопептидные бактериоцины, не имеющие каких-либо свойств, характерных для IIa бактериоцинов.
Бактериоцины подкласса IId
Этот подкласс бактериоцинов выделяют немногие авторы [20], предлагая отдельно рассматривать пептиды с особенным строением. Примером такого белкового соединения является бактериоцин AS-48. Бактериоцин AS-48 - первый из выделенных, один из наиболее изученных бактериоцинов [6, 12, 18, 22]. Он является циклическим пептидом с молекулярной массой 7,2 кДа, представляет собой полипептидную цепь из 70 аминокислотных остатков, замкнутых в кольцо. Известно, что подобная молекула обладает высокой конформационной изменчивостью [20]. Это свойство позволяет молекуле бактериоцина AS-48 связываться с поверхностными структурами широкого спектра бактерий. Девять генов, необходимых для синтеза, модификации, секреции этого первоначально пребактериоцина, устойчивости к нему продуцента, образуют кластер в плазмиде pAD1 (59 кБ) [21].
Бактериоцины III класса
Бактериоцины этого класса представляют собой антимикробные чувствительные к изменениям температуры белки с молекулярной массой более 30 кДа, способные расщеплять муреин клеточной стенки. К их числу относится энтеролизин А.
Классификация бактериоцинов позволяет систематизировать наши знания о природе и механизмах действия соответствующих групп антимикробных пептидов. Это создает возможность осуществлять подбор антимикробных пептидов или их комбинаций для терапии и профилактики соответствующей инфекционной патологии [17].
43 Вестник ВСГУТУ. № 2 (59). 2016
Механизм действия бактериоцинов
Учитывая большое разнообразие химического строения бактериоцинов, можно предположить, что они воздействуют на жизненно важные функции чувствительных клеток, но большинство действуют, образуя в мембране поры или каналы, способствующие нарушению мембранного потенциала чувствительных клеток [24, 25].
Анионные липиды цитоплазматической мембраны являются основными рецепторами бактериоцинов молочнокислых бактерий для стимуляции порообразования. Бактериоцины I класса могут вызывать формирование пор по типу клиноподобной модели, в то время как бактериоцины II класса способны функционировать, создавая поры, посредством которых пептиды располагаются параллельно мембранной поверхности и оказывают влияние на структуру мембран [36].
Наиболее хорошо изучен механизм действия бактериоцинов на примере низина (рис.).
При изучении действия низина стало ясно, что часть C-конца молекулы деполяризует цитоплазматическую мембрану, что ведет к мгновенной остановке синтеза компонентов клеточной стенки и всех биосинтетических процессов в клетке. Основной характер действия низина - это взаимодействие с отрицательно заряженными мембранами (билипидный слой), предпочтительно с мембранами, содержащими анионные липиды. Грамположительные бактерии имеют более высокие концентрации анионного липида в их цитоплазматической мембране по сравнению с грамотрицательными. Бактериоцин сначала связывает его C-конец с анионными липидами, после чего N-конец опускается в липид мембраны и пептид принимает параллельное положение на мембранной поверхности. И, в конечном счете, целый пептид имеет возможность переместиться по всей длине поры [24].
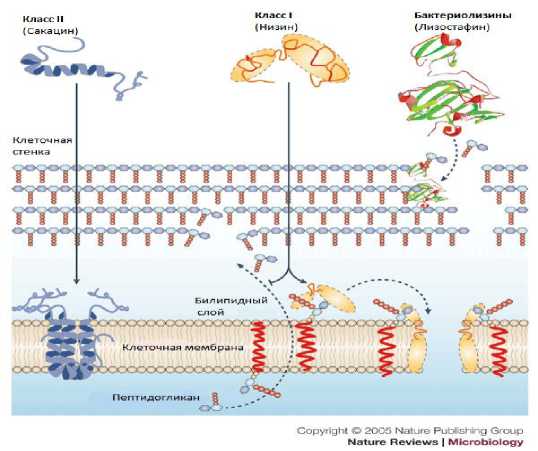
Рисунок - Механизм действия бактериоцинов
Низин ингибирует синтез муреина клеточной стенки. В результате действия антибиотика происходит накопление промежуточного липида, с которым низин образует прочный комплекс, что и приводит к остановке дальнейшего синтеза пептидогликана бактериальной стенки. Низин может вызывать лизис клеток вследствие необратимой деструкции клеточной мембраны, которая регулирует осмотическое давление клетки. Он изменяет поверхностное натяжение цитоплазматической мембраны, что приводит к нарушению клеточной проницаемости и, как следствие этого, к бактериальному лизису.
Перспективы применения бактериоцинов молочнокислых бактерий
Бактериоцин низин (имеет GRAS-cтатус – «признанный как безопасный», Е234), образуемый Lactococcus lactis, с успехом используют для увеличения сроков годности продуктов питания в пищевой промышленности многих стран уже более 50 лет. Низин впервые был ис- пользован в промышленном производстве как консервант в Великобритании для производства и сохранения сыров, и с этого момента его стали активно использовать в пище и напитках. В дальнейшем эффективность использования низина возможна вследствие его синер-гидного действия с хелатными и другими бактериоцинами как добавка в новые технологические процессы пищевой промышленности, при которых, возможно, будут пересмотрены режимы стерилизации при производстве консервов и пресервов для сохранения пищевой ценности продуктов [25].
Потребительский спрос на минимально обработанные пищевые продукты или «свежую еду» без химических консервантов стимулировал исследователей на поиск натуральных антимикробных средств [26]. Кроме того, бактериоцины, в отличие от антибиотиков, полностью расщепляются в организме, и вероятность возникновения осложнений от бактериоци-нов минимальна, а применение антибиотиков имеет для человека отрицательные последствия. Однако сферы применения известных форм бактериоцинов, как уже отмечалось, ограничены из-за низкой активности при нейтральных или щелочных значениях pH. Поэтому поиск бактериоцинов с улучшенными физико-химическими свойствами и широким антимикробным спектром представляет большой интерес не только для пищевой промышленности, но и для фармацевтической - при создании альтернативных антибиотиков, что является одной из самых актуальных проблем в области микробиологии и биотехнологии.
Список литературы Бактериоцины молочнокислых бактерий
- Нурушев М.Ж., Стоянова Л.Г., Бекеева С.А. и др. О проекте создания природного консерванта пищевых продуктов на основе бактериоцинов молочнокислых бактерий в Казахстане//Л.Н. Гумилев атындагы ЕҰУ Хабаршысы. -Вестник ЕНУ им. Л.Н. Гумилева. -2011. -№ 4.
- Тюрин М.В., Шендеров Б.А., Рахимова Н.Г. и др. К механизму антагонистической активности лактобацилл//Журн. микробиол., эпидемиол. и иммунобиол. -1989. -№ 2. -С. 3-8.
- Шлегель Г. Общая микробиология. -М.: Мир, 1987. -567 с.
- Блинкова Л.П., Альтшуллер М.Л., Дорофеева Е С. и др. Молекулярные основы продукции и действия бактериоцинов//Журн. микробиол. -2007. -№ 2. -С. 97-104.
- Ускова М.А. Изучение свойств пробиотических молочнокислых бактерий как биологически активных компонентов пищи: автореф.. канд. биол. наук. -М., 2010. -28 с.
- Minah C.J., Morero R.D. Inhibition of enterocin CRL35 antibiotic activity by mono-and divalent ions//Lett. Appl. Microbiol. -2003. -Vol. 37, N 5. -P. 374-379.
- Van den Berghe E., de Winter T., de Vuyst L. Enterocin A production by Enterococcus faecium FAIR-E 406 is characterised by a temperature and pH-dependent switch-off mechanism when growth is limited due to nutrient depletion//Int. J. Food Microbiol. -2006. -Vol. 107, Is. 2. -P. 159-165.
- Moreno M. R. F., Sarantinopoulos P., Tsakalidou, De Vuyst L. The role of appplication of enterococci in food and health//Int. J. Food Microbiol. -2006. -Vol. 106. -P. 1-24.
- De Vuyst L., Leroy F. Bacteriocins from Lactic Acid Bacteria: Production, Purification, and Food Applications//Mol. Microbiol. Biotechnol. -2007. -Vol. 13. -P. 194-199.
- Kraus A., Peschel D. Molecular Mechanisms of Bacterial Resistance to Antimicrobial Peptides. -Berlin; Heidelberg, 2006. -P. 231-250.
- Oscariz J.C., Pissabarro A.G. Classification and mode of action of membrane-active bacteriocins produced by gram-positive bacteria//Int. Microbiol. -2011. -N 18. -P. 13-19.
- Segarra R.A., Gilmore M. S., Booth M C. Genetic structure of the Enterococcus faecalis plasmid pAD1-encoded cytolytic toxin system and its relationship to lantibiotic determinants//J. Bacteriol. -2004. -Vol. 176. -P. 734-735.
- Bittencourt E., Suzart S. Occurrence of virulence-associated genes in clinical Enterococcus faecalis strains isolated in Londrina, Brazil//J. Med. Microbiol. -2004. -Vol. 53. -P. 1069-1073.
- Drider D., Fimland G., Héchard Y. The Continuing Story of Class IIa Bacteriocins//Microbiol. Mol. Biol. Rev. -2006. -Vol. 70, N 2. -Р. 564-582.
- Klaenhammer T.R. Genetics of bacteriocins produced by lactic acid bacteria//FEMS Microbiol. Rev. -2008. -N 22. -P. 39-86.
- Diep D.B., Havarstein L.S., Nes I.F. A family of bacteriocin ABC transporters carry out proteolytic processing of their substrates concomitant with export//Mol. Microbiol. -2008. -N 26. -P. 230-261.
- Ennahar S., Descamps N. Anti-Listerial effect of enterocin A, produced by cheese-isolated Enterococcus faecium EFMO1, relative to other bacteriocins from lactic acid bacteria//J. Appl. Microbiol. -2010. -Vol. 76, Is. 3. -P. 449-457.
- Brasseui R. Bennik M. H. J., Berlinda V. A novel bacteriocin from vegetable associated Enterococcus mundtii: full characterization and interaction with target organisms//Biochim. Biophys. Acta: Biomembr. -2008. -Vol. 1679, Is. 1. -P. 47-58.
- Zendo T., Eungruttanagorn N., Fujoka S. Identification and production of a bacteriocin from Enterococcus mundtii QU2 isolated from soybean//J. Appl. Microbiol. -2005. -Vol. 99, Is. 5. -P. 1181-1516.
- Farias R. N., Farias M. E., Holgado R. Letters in Purification and N-terminal acid sequence of enterocin CRL35, a pediocin-like bacteriocin produced by Enterococcus faecium CRL35//Appl. Microbiol. -2005. -Vol. 56. -P. 417-419.
- Dicks L. M. T., Balla E., Toit M. Characterization and Cloning of the Genes Encoding Enterocin 1071A and Enterocin 1071B, Two Antimicrobial Peptides Produced by Enterococcus faecalis BFE 1071//Appl. Environ. Microbiol. -2013. -Vol. 93, N 4. -P. 1298-1304.
- Criado R., Dzung B., Diep A. et al. Complete Sequence of the Enterocin Q-Encoding Plasmid pCIZ2 from the Multiple Bacteriocin Producer Enterococcus faecium L50 and Genetic Characterization of Enterocin Q Production and Immunity//Ibid. -2006. -Vol. 72, N 10. -P. 6653-6666.
- Глушанова Н.А., Шендеров Б.А. Взаимоотношения пробиотических и индигенных лактобацилл хозяина в условиях совместного культивирования in vitro II Журнал микробиол. -2005. -№ 2. -С. 56-61.
- Surugau L.N. Peptide separation by capillary electrophoresis with ultraviolet detection: some simple approaches to enhance detection sensitivity and resolution//The Malaysian Journal of Analytical Sciences. -2011. -Vol. 15 (2). -P. 273-287.
- Сультимова Т.Д. Выделение активных бактериоцинобразующих лактококков и их практическое использование: автореф.. канд. биол. наук. -М., 2006. -30 с.
- Ермоленко Е.И. Бактериоцины энтерококков: проблемы и перспективы использования II Вестник Санкт-Петербургского университета. Сер. 11. -2009. -Вып. 3.
