Бактериопланктон озер Раифы (Татарстан, Россия)
")
Автор: Уманская М.В., Горбунов М.Ю., Унковская Е.Н.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 4 т.9, 2007 года.
Бесплатный доступ
В работе представлены первые сведения о развитии бактериопланктона в озерах Раифского участка Волжско-Камского государственного природного биосферного заповедника. Дана общая характерис- тика бактериопланктона, его численности и биомассы, морфологического и размерного разнообразия в озерах с различающимися условиями среды. Описано обнаружение некоторых морфологически своеобразных видов бактерий, в том числе аноксигенных фототрофных бактерий. Обсуждаются осо- бенности развития бактериопланктона в лесных гумифицированных озерах.
Короткий адрес: https://sciup.org/148198051
IDR: 148198051 | УДК: 574.57+579.26
Bacterioplankton of Raifa lakes (Tatarstan, Russia)
The first data of bacterioplankton development in several lakes in Raifa part of Volga-Kama natural reserve are presented. The general characteristics of bacterioplankton community, its abundance, biomass and diversity in lakes with different natural conditions are given. In some lakes, morphologically conspicuous species including anoxygenic phototrophic bacteria and consortia are detected. Peculiarities of bacterioplankton development in boreal humic lakes are discussed.
Текст научной статьи Бактериопланктон озер Раифы (Татарстан, Россия)
1Институт экологии Волжского бассейна РАН, г. Тольятти
2Волжско-Камский государственный природный биосферный заповедник, пос. Садовый
В работе представлены первые сведения о развитии бактериопланктона в озерах Раифского участка Волжско-Камского государственного природного биосферного заповедника. Дана общая характеристика бактериопланктона, его численности и биомассы, морфологического и размерного разнообразия в озерах с различающимися условиями среды. Описано обнаружение некоторых морфологически своеобразных видов бактерий, в том числе аноксигенных фототрофных бактерий. Обсуждаются особенности развития бактериопланктона в лесных гумифицированных озерах.
Озера Раифского участка Волжско-Камского государственного природного заповедника, находящиеся на южной границе таежной зоны, разнообразны по особенностям гидрологического режима и гидрохимического состава вод [1]. Исследования их экологического состояния проводятся в течение многих лет [1-3 и др.], однако до настоящего времени сведений о количественном развитии и особенностях функционирования бактериального планктонного сообщества в них не было. В июле 2006 г. впервые было проведено определение основных показателей развития бактериопланктона в озерах Раифы, результаты которого представлены в настоящей работе.
Методы исследования
Исследования проводили 12-18 июля 2006 г. в условиях устойчивой антициклонической погоды. Пробы воды отбирали батометром Руттнера в наиболее глубокой части озер, по глубинам. Микробиологические пробы переливали в стерильные склянки и фиксировали профильтрованным раствором формалина. Общую численность и биомассу бактерий определяли на мембранных фильтрах Влади-пор (диаметр пор - 0,2 мкм) [4, 5]. Морфологическую и размерную структуры бактериопланктона определяли, как описано ранее [6]. В соответствии с объемом клеток выделено 9 размерных классов: 1 – менее 0,025 мкм3; 2 – 0,025-0,049 мкм3; 3 – 0,05-0,09 мкм3; 4 – 0,100,19 мкм3; 5 – 0,20-0,39 мкм3; 6 – 0,40-0,79
мкм3; 7 – 0,80-1,59 мкм3; 8 – 1,60-3,19 мкм3; 9 – больше 3,20 мкм3. Для оценки размерной структуры рассчитывали вклад клеток каждого класса в общую численность или биомассу. Видовой состав фототрофных бактерий определяли по [7] с учетом современных таксономических ревизий [8].
В отобранных пробах определяли физические, электрохимические и химические показатели [9]. Первичную продукцию определяли скляночным методом в кислородной модификации [10].
Результаты и их обсуждение
Физико-химическая характеристика изучаемых озер
Расположение изучаемых озер, их основные морфологические и физико-химические показатели представлены на рис. 1 и в табл. 1 и 2.
Все исследованные озера имеют мало- и среднеминерализованную воду кальций-гид-рокарбонатного типа, с низким содержанием сульфатов и хлоридов. Хотя отмечены значительные межгодовые колебания общей минерализации озер, связанные, очевидно, с количеством осадков и интенсивностью весеннего паводка, соотношение основных ионов в гидрохимическом составе вод озер в целом изменяется мало [1]. Большинство озер имеет высокую цветность воды из-за повышенного содержания растворимых гуминовых кислот и солей железа (табл. 1 и 2).
Таблица 1. Морфометрические и гидрофизические показатели озер
|
Озеро |
Площадь, м2 |
Глубина макс., м |
Прозрачность, м |
Цветность, оPt |
Температура, оС * |
|
Белое |
64 100 |
3,8 |
1,4 |
80 |
26,5/16,4 |
|
Линево |
69 700 |
4,5 |
0,95 |
210 |
27,0/9,4 |
|
Илантово |
47 800 |
1,8 |
1,4 |
160 |
25,0/19,0 |
|
Гнилое |
8 000 |
4,5 |
0,6 |
н/д |
27,5/8,5 |
|
Долгое |
4 000 |
12 |
1,5 |
220 |
26,9/7,5 |
|
Карасиха |
4 000 |
10 |
0,5 |
380 |
25,0/8,0 |
|
Раифское |
319 900 |
17 |
1,65 |
н/д |
25,0/4,0 |
Примечание: * - перед чертой - поверхностный; за чертой - придонный слой.
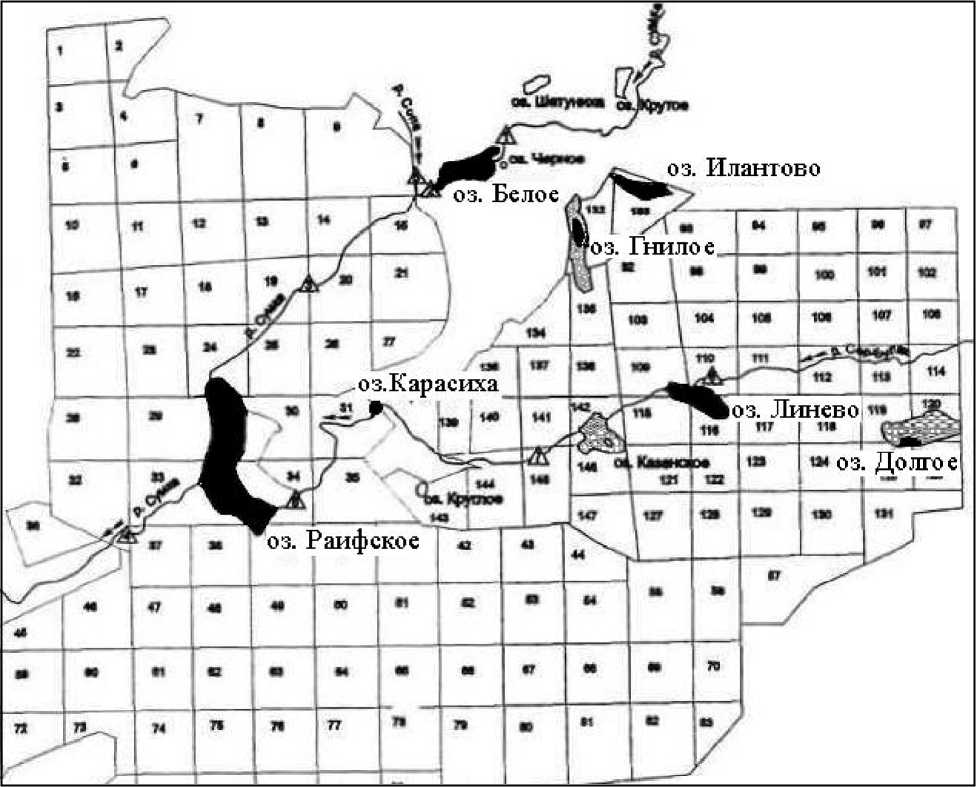
Рис. 1. Схема расположения исследованных озер Раифского участка Волжско-Камского государственного природного заповедника (по [1]). Черным цветом отмечены исследованные озера, серым - сплавина на озерах
Измерение физико-химических показателей показало, что все исследованные озера, включая мелководные, термически стратифицированы. Кроме главного, в большинстве озер был сформирован приповерхностный термоклин, обусловленный интенсивным дневным прогревом поверхностного слоя и слабым ветровым воздействием (рис. 2). Только в озерах Белое и Гнилое присутство- вал поверхностный гомотермический слой. В остальных водоемах поверхностный градиент температуры плавно переходил в главный термоклин, поэтому граница между эпи-и металимнионом выражена неявно. Верхняя граница гиполимниона в оз. Гнилое расположена на глубине 2 м, Карасиха – 3 м, Долгое – 4 м, Раифское – 6-8 м. В озерах Линево и Белое гиполимнион сформирован только в
Таблица 2. Некоторые показатели химического состава воды озер
|
Озеро |
Сухой остаток, мг/л |
Электропроводнос ть, mkSm/см |
Fe общ , мг/л |
H 2 S+HS?, мг/л |
O 2 , мг/л |
|
|
Белое |
Пов. |
280 |
380 |
0,27 |
0 |
9,6 |
|
Дно |
353 |
465 |
1,53 |
0,059 |
6,71 |
|
|
Илантово |
Пов. |
53 |
75 |
0,95 |
0 |
6,45 |
|
Дно |
97 |
86 |
1,30 |
0 |
2,08 |
|
|
Гнилое |
Пов. |
124 |
38 |
0,21 |
0 |
н/д |
|
Дно |
168 |
138 |
3,94 |
0,413 |
0,14 |
|
|
Линево |
Пов. |
71 |
88 |
0,76 |
0 |
8,60 |
|
Дно |
229 |
195 |
15,10 |
0,173 |
0,00 |
|
|
Раифское |
Пов. |
142 |
273 |
0,18 |
0 |
9,88 |
|
Дно |
183 |
383 |
3,16 |
0,085 |
0,00 |
|
|
Карасиха |
Пов. |
238 |
98 |
0,58 |
0 |
7,30 |
|
Дно |
274 |
164 |
2,77 |
0,431 |
0,00 |
|
|
Долгое |
Пов. |
52 |
47 |
0,53 |
0 |
7,03 |
|
Дно |
74 |
66 |
1,56 |
0,059 |
0,30 |
|
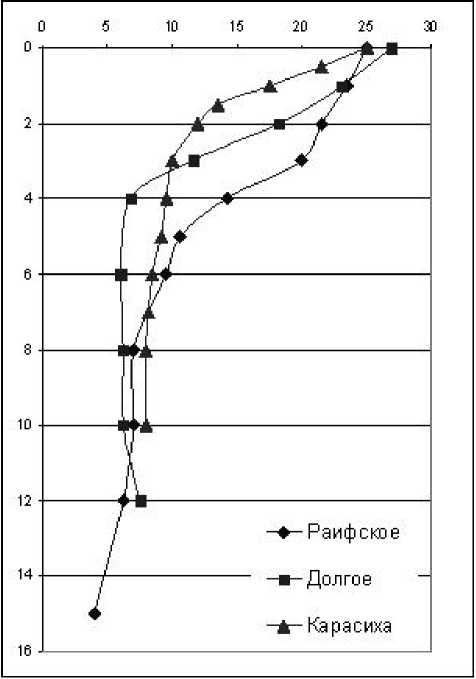
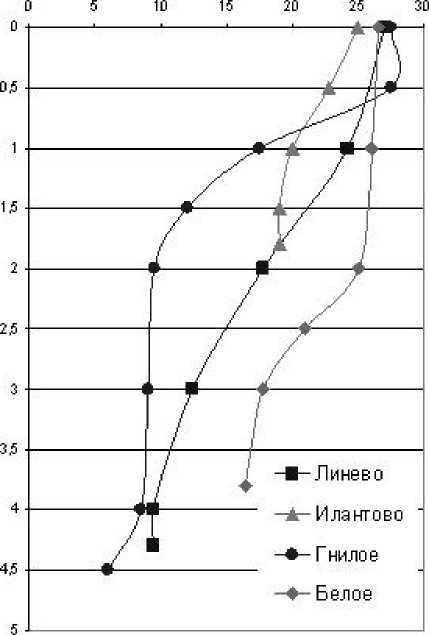
Рис. 2. Вертикальные профили температуры в исследованных озерах узком (менее 1 м) придонном слое; в оз. Илан-тово главный термоклин и, следовательно, мета- и гиполимнион отсутствует.
Во всех озерах, кроме Илантово и Белое, концентрация кислорода в гиполимнионе не достигает 1 мг/л; в ряде случаев в придонном слое кислород не обнаруживается (табл. 2; рис. 5).
Особенностью исследованных озер является повышенное содержание NO2- - от 0,02 до 0,21 мг/л. Во всей водной толще оз. Илан-тово и в поверхностном слое воды озер Ра-ифское, Белое, Долгое и Гнилое содержание неорганического фосфора не превышает 0,1 мг/л. В придонном слое воды этих озер его концентрация составляет 0,2-1,2 мг/л. В озе-
Общая характеристика бактериопланктона
Численность бактерий в исследованных озерах изменялась в пределах 2,00-14,01 млн кл./мл, а биомасса – 0,281-4,229 мг/л. Во всех озерах общая численность и биомасса бактерий в придонном слое воды были выше, чем в поверхностном (табл. 3). Увеличение биомассы связано не только с возрастанием численности бактерий, но и с увеличением среднего размера бактериальных клеток в сообществе. Только в оз. Илантово средний размер клеток во всем столбе воды оставался в пределах 0,16-0,18 мкм3, тогда как в остальных озерах средний размер клеток поверхностного слоя воды изменялся в пределах от 0,10 до 0,21 мкм3, а придонного – от 0,20 до 0,34 мкм3.
Морфологически бактериопланктон озер был разнообразен. Обнаружены кокки, ово-иды, палочки, вибриоидные клетки, спириллы, нитевидные (прямые и извитые, одиноч- ные и многоклеточные), а также клетки сложной формы, например, звездообразной, серповидной и др. В составе планктона преобладали одиночные свободноплавающие клетки. В эпилимнионе озер Гнилое, Долгое и Раифское обнаружены бактерии с характерной коринеподобной морфологией. Кроме того, наблюдались микроколонии, консорциумы, скопления на частицах детрита и вокруг живых и отмирающих клеток водорослей. В поверхностном слое воды всех озер 37-57% общей численности и 18-50% общей биомассы составляли кокки и овоидные клетки. В придонном слое их доля уменьшалась и составляла 18-42% и 7-28% от общей численности и биомассы, соответственно. Количество нитевидных клеток (длиной более 10 мкм), в общем, не зависело от глубины, и изменялось в разных озерах от 0,1 до 1,4% от общей численности. Вклад нитей в общую биомассу был более существенным – 0,5-12%.
Таблица 3. Общая характеристика бактериопланктона озер
|
Озеро |
Горизонт |
Общая численность бактерий, млн кл./мл |
Общая биомасса бактерий, |
|
|
мг/л |
мкг С/л |
|||
|
Белое |
Пов. |
2,69 |
0,281 |
59,21 |
|
Дно |
2,86 |
0,809 |
128,59 |
|
|
Илантово |
Пов. |
4,07 |
0,747 |
138,26 |
|
Дно |
5,74 |
0,894 |
170,96 |
|
|
Гнилое |
Пов. |
3,91 |
0,714 |
129,77 |
|
Дно |
9,41 |
2,182 |
363,67 |
|
|
Линево |
Пов. |
3,99 |
0,852 |
147,93 |
|
Дно |
9,10 |
3,126 |
469,45 |
|
|
Раифское |
Пов. |
2,19 |
0,285 |
58,78 |
|
Дно |
3,00 |
1,058 |
143,38 |
|
|
Карасиха |
Пов. |
4,08 |
0,407 |
87,94 |
|
Дно |
11,61 |
3,753 |
581,84 |
|
|
Долгое |
Пов. |
4,11 |
0,519 |
104,80 |
|
Дно |
4,71 |
0,920 |
163,12 |
|
В эпилимнионе трех из исследованных озер были обнаружены планктомицеты Planctomyces bekefii и P. guttaeformis. Они встречены единично в озерах Белом и Раиф-ском (0,009 и 0,005 млн. кл./мл соответственно), но довольно многочисленны в оз. Лине-во. В нем оба вида присутствовали в равных количествах до глубины 0,5 м (0,04 млн кл./ мл в сумме), а в слое от 0,5 до 2 м регистрировался только P. bekefii (уменьшаясь с глу- биной от 0,04 до 0,003 млн. кл./мл). В анаэробной зоне (глубже 2 м) планктомицеты полностью исчезли. В эпилимнионе этих озер в момент наблюдений было выявлено массовое цветение цианобактерий [11], которое, как известно, часто сопровождается развитием планктомицетов, но в целом они и их функции в планктоне водоемов остаются мало изученными.
В стратифицированных озерах с анаэроб- ным гиполимнионом развивались фототроф-ные бактерии. Зеленые серные бактерии (сем. Chlorobiaceae: Chlorobium limicola, Ch. luteum, Chlorobium spp., Pelodiction clatratiforme, Ancalochloris perfilievii и др. были обнаружены в зоне хемоклина и в гиполимнионе озер Гнилое, Карасиха, Долгое и Линево. Общая численность бактерий сем. Chlorobiaceae в этих озерах достигала 1,5 млн. кл./мл. Неподвижные консорции Chloroplana vacuolata, в состав которых входят зеленые серные бактерии, были обнаружены в оз. Линево (до 0,45 млн. кл./мл) и в оз. Карасиха (до 0,30 млн. кл./мл). Наряду с ними в этих озерах были зарегистрированы и подвижные фототроф-ные консорции Chlorochromatium и Pelochro-matium.
В оз. Линево в значительных количествах обнаружены также нитчатые зеленые бактерии Chloronema giganteum (сем. Chloro-flexaceae, тип Chloroflexi), представленные двумя формами, прямой и спиральной, в примерно равных количествах. Численность клеток на разных глубинах изменялась в пределах 0,06-0,35 млн. кл./мл. Нитчатые бактерии, морфологически идентичные спиральной форме Chloronema giganteum , были обнаружены и в оз. Раифское. В отличие от оз. Ли-нево, в оз. Раифское их слизистый чехол сильно инкрустирован железом, причем интенсивность инкрустации трихомов увеличивается с глубиной. Поскольку в оз. Раифское, судя по данным пигментного анализа, практически отсутствует бактериохлорофилл d, основной светособирающий пигмент Chloronema , для определения таксономической принадлежности этой формы необходимы дополнительные исследования.
Пурпурные серные бактерии (сем. Chromatiaceae, тип Proteobacteria) были встречены в меньших количествах, преимущественно в области хемоклина озер Линево, Гнилое, Карасиха и Долгое (максимальная численность составляла 0,14; 0,01; 0,18 и 0,06 млн. кл./мл соответственно). В указанных озерах эти бактерии довольно разнообразны: в оз. Линево обнаружены Thiocapsa spp. (доминировали типичные формы, единично встречались крупные колонии с газовыми вакуолями типа Amoebobacter), Allochro-matium spp., Thiopedia rosea. В озерах Долгое и Карасиха обнаружены Thiocapsa spp., Allochromatium spp., Thiodiction bacillosum и Chromatium minus; в оз. Гнилое единично встречены только бактерии Allochromatium sp.
В озерах Линево, Раифское и Илантово кислородным методом были определены первичная продукция фитопланктона (А) и деструкция (D) в фотическом слое. Расчет продукции показал, что она максимальна в оз. Раифское (0,9 г С/м2) и минимальна в макро-фитном, сильно заросшем оз. Илантово (0,11 г С/м2). Только в оз. Раифское продукция на всех горизонтах фотического слоя (0-3 м) превышала деструкцию (отношение A/D на разных глубинах составило 1,75-2,82). В макро-фитном оз. Илантово деструкция во всей водной толще превышала продукцию фитопланктона, что, видимо, связано с использованием бактериями продукции макрофитов, которые массово развиваются в этом озере.
Особенности вертикального распределения бактериопланктона
Количественные и качественные показатели бактериопланктона исследованных озер существенно различаются не только от озера к озеру, но от горизонта к горизонту в одном и том же озере. В озерах Илантово и Белое микробиологические показатели определяли только в поверхностном и придонном горизонтах. Вертикальное распределение общей численности, биомассы и среднего объема бактериальных клеток в пяти остальных озерах показано на рис. 3 и 4. В озерах Гнилое, Раифское, Карасиха и Долгое главный максимум численности бактериопланктона располагался примерно в области термо- и/или оксиклина. В оз. Линево при отборе проб через 0,5-1 м (13.07.2006), наибольшая численность была зарегистрирована в придонном слое. При более дробном отборе проб из области хемоклина (1,5-3,5 м, через каждые 20 см, 14.07.2006) был обнаружен очень узкий максимум численности на глубине 2,4 м (13,42 млн. кл./мл).
Из-за изменения среднего объема клеток вертикальные изменения биомассы бактери-
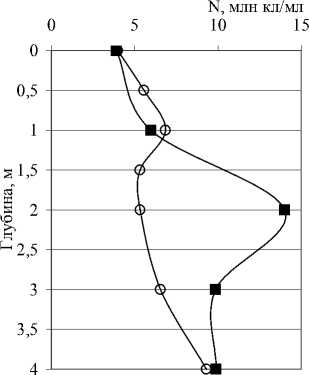
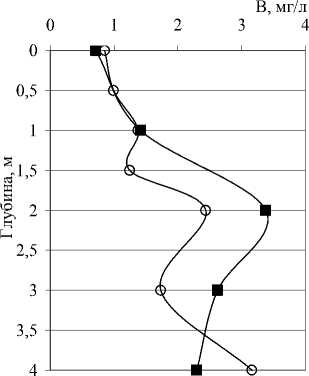
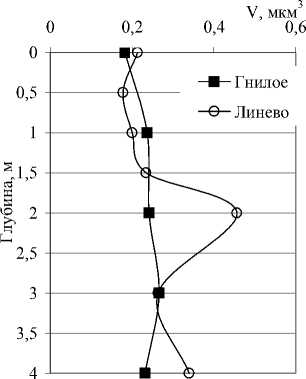
Рис. 3. Вертикальное распределение общей численности (N), биомассы (B) и среднего объема (V) бактериальных клеток в исследованных озерах
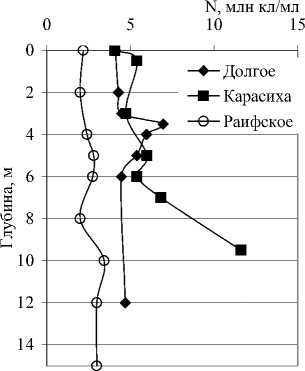
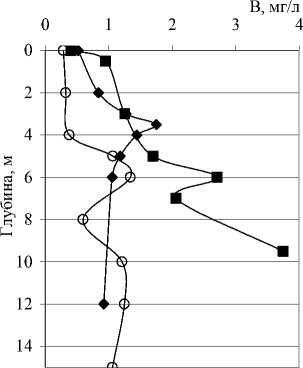
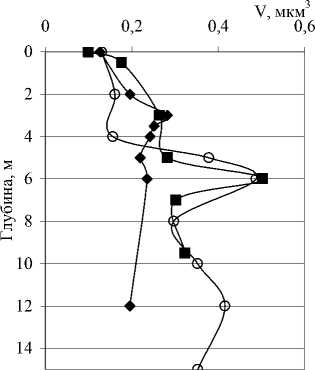
Рис. 4. Вертикальное распределение общей численности (N), биомассы (B) и среднего объема (V) бактериальных клеток в исследованных озерах опланктона не вполне совпадали с изменениями численности. Средний объем, в свою очередь, определялся изменениями морфологической и размерной структуры сообщества по вертикали.
Изменения морфологической структуры бактериопланктона с глубиной во всех озерах выражаются в уменьшении доли овоид-ных клеток в общей численности и биомассе. Причем эти изменения коррелируют с изменением концентрацией растворенного кислорода в воде. В слоях воды, где она превышает 3 мг/л, доля кокков составляет в среднем 47%, при более низкой концентрации и при полном отсутствии кислорода их доля снижается до 36-35% общей численности.
Такая же тенденция наблюдается и для биомассы бактериопланктона. Соответственно, вклад палочковидных клеток в численность и биомассу увеличивается с уменьшением содержания кислорода. Зависимости численности и биомассы нитевидных клеток от глубины и концентрации кислорода не выявлено.
Размерная структура бактериального сообщества тоже существенно меняется с глубиной. Как видно из рис. 5, особенно резкие изменения наблюдались на верхней границе оксиклина, т.е. когда концентрация кислорода становилась меньше 1 мг/л. При этом доля мелких клеток (1-4 размерные классы) уменьшается, а крупных (6-9 классы) возрастает.
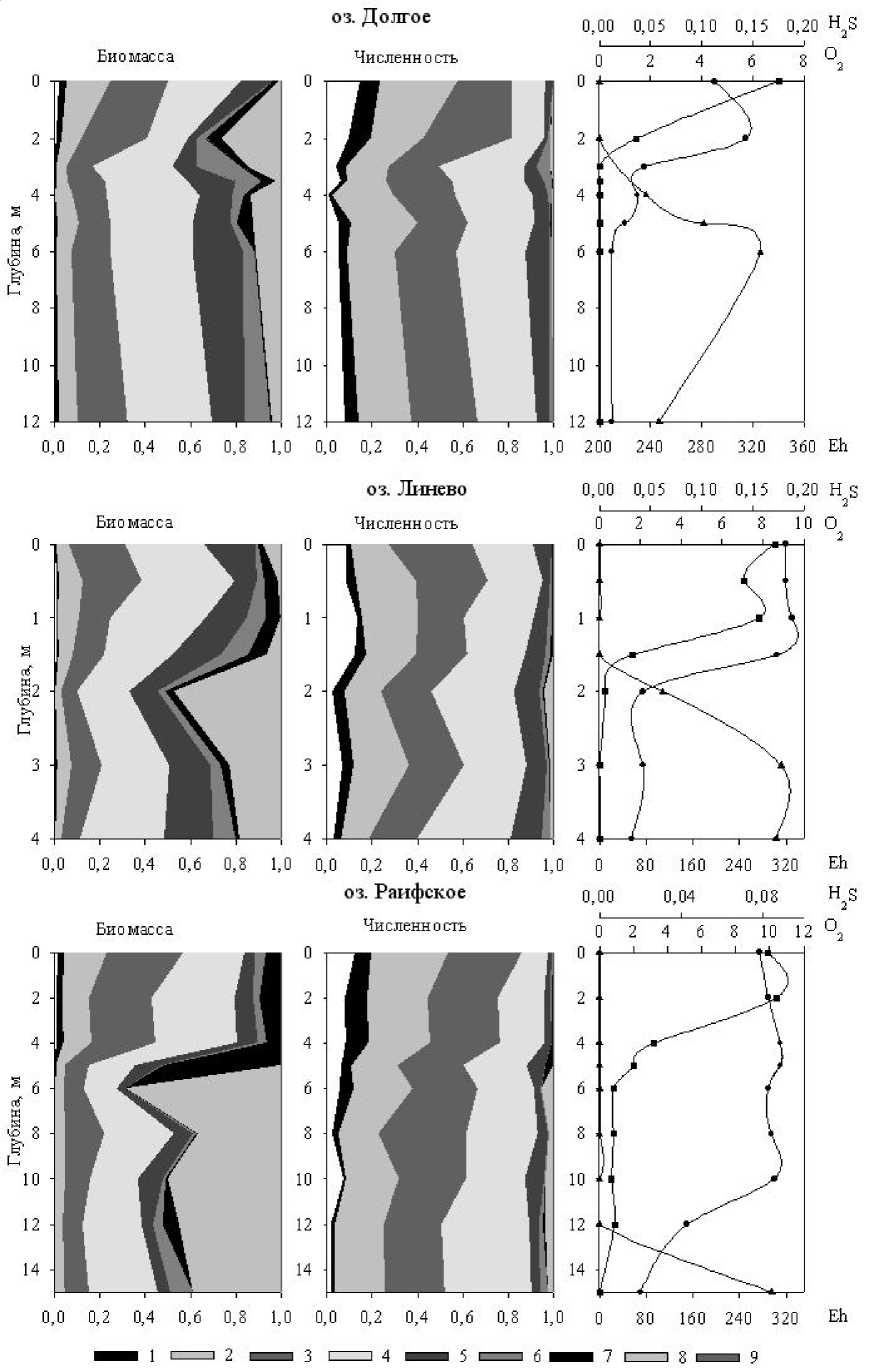
* —^- 02 —*- Eh
Рис. 5. Влияние вертикального распределения некоторых физико-химических факторов на изменение размерной структуры бактериопланктона: 1-9 – номера размерных классов бактериальных клеток
Особенно четко это видно на графиках изменения структуры по биомассе (рис. 5). Дальнейшее снижение и полное исчерпание кислорода не вызывало аналогичной перестройки сообщества.
При сохранении общей тенденции, в каждом конкретном озере доминирование того или иного размерного класса и их соотношение зависит от освещенности переходного слоя, наличия и концентрации сероводорода и сульфидов, концентрации неорганического фосфора и некоторых других факторов. Так, в оз. Линево, в котором верхняя граница содержащего сероводород слоя находится в фоти-ческой зоне, в этом слое обнаружены крупные (7-9 классы) овоидные клетки различных серных пурпурных бактерий.
Дробный отбор в области хемоклина оз. Линево позволил выявить некоторые закономерности вертикального распределения отдельных видов фототрофных бактерий (рис. 6). Максимум численности Chloronema giganteum был зарегистрирован на глубине 2 м, далее ее количество уменьшалось. Число клеток в прямых и спиральных трихомах было близким и изменялось от 13 до 20 кл./трихом. Интересно отметить, что количество двух-четырех клеточных фрагментов трихомов увеличивалось с глубиной. Численность фото-трофных клеток, входящих в состав консорций Chloroplana vacuolata возрастала с глубиной, образуя промежуточный максимум на глубине 2,4 м.
Основная масса пурпурных бактерий (сем. Chromatiaceae ) была локализована на глубине 2-2,8 м, причем они образовывали 2 пика численности, на глубине - 2 м и 2,6 м (рис. 6). На глубине 2 м преобладали крупные колонии с газовыми вакуолями типа Amoebo-bacter и также колониальные Thiopedia rosea . В области второго пика по численности доминировали одиночные и образующие небольшие колонии виды Thiocapsa spp. В нижележащих слоях пурпурные бактерии были представлены почти исключительно
Список литературы Бактериопланктон озер Раифы (Татарстан, Россия)
- Унковская Е.Н., Мингазова Н.М., Павлова Л.Р. Гидрологическая и гидрохимическая характеристика водоемов Раифы//Тр. Волжско-Камск. гос. природн. заповедника. 2002. Вып. 5.
- Деревенская О.Ю., Унковская Е.Н., Мингазова Н.М., Павлова Л.Р. Структура сообществ зоопланктона озер Раифского участка Волжско-Камского заповедника и его охранной зоны//Тр. Волжско-Камского государственного природного заповедника. 2002. Вып. 5.
- Палагушкина О.В., Бариева Ф.Ф., Унковская Е.Н. Видовой состав, биомасса и продуктивность фитопланктона озер Раифского участка Волжско-Камского заповедника и его охранной зоны//Тр. Волжско-Камск. гос. природн. заповедника. 2002. Вып. 5.
- Norland S. The relation between biomass and volume of bacteria//Handbook of methods in aquatic microbiology./Eds. P. Kemp, B. Sherr, E. Sherr, J. Cole. Lewis Publishers, Boca Raton, 1993.
- Straskrabova V., Callieri C., Carrillo P. et al. Pelagic food web in mountain lakes//J. Limnol. 1999. V. 58. № 2.
- Уманская М.В. Микробиологическая характеристика малых эвтрофных озер национального парка «Самарская Лука»//Изв. Самар. НЦ РАН. 2004. Т. 7. № 1.
- Pfennig N., Truper H.G. Anoxigenic phototrophic bacteria//Bergey's Manual of Systematic Bacteriology. V. 3./Ed. James T. Staley. USA, Williams & Wilkins, 1989.
- Garrity G.M., Bell J.A., Lilburn T.G. Taxonomic outline of the Procaryotes//Bergey's manual of systematic bacteriology. 2d ed. Release 5.0 May 2004. Springer, 2004. -DOI 10.1007/bergeysoutline200405
- Унифицированные методы анализа вод/Под ред. Ю.Ю. Лурье. 2-е изд. М.: Химия, 1973.
- Методика изучения биогеоценозов внутренних водоемов. М.: Наука, 1975.
- Тарасова Н.Г. Водоросли биоценозов высших водных растений озер Волжско-Камского заповедника//Материалы конф. «Природное наследие России-2». Уфа, 2007.
- Горленко В.М., Локк С.И. Вертикальное распределение и особенности видового состава микроорганизмов некоторых стратифицированных озер Эстонии//Микробиология. 1979. Т. 48.
- Лаптева Н.А. Микробиологическая характеристика некоторых карстовых озер Марийской АССР//Пресноводные гидробионты и их биология/Тр. ИБВВ АН СССР. 1983. Вып. 48(51).
- Lindstrom E.S. Bacterioplankton community composition in a boreal forest lake//FEMS Microbiol Ecol. 1988. V. 27.
- Abella C.A., Garcia-Gil L.J. Microbial ecology of planktonic filamentous phototrophic bacteria in holomictic freshwater lakes//Hydrobiol. 1992. V. 243/244.
- Горбунов М.Ю., Уманская М.В. Аноксигенные фототрофные бактирии в водоемах особо охраняемых территорий Самарской области//Экологические проблемы заповедных территорий России/Под ред. С.В.Саксонова. Тольятти: ИЭВБ РАН, 2003.
- Горбунов М.Ю., Уманская М.В. К вертикальному распределению прокариотического фототрофного планктона в Нижнем пруду Самарского Ботанического сада//Самарская Лука: Бюл. 2007. Т. 16. № 1-2 (19-20).
